
Fabp7 Is Required for Normal Sleep Suppression and Anxiety-Associated Phenotype following Single-Prolonged Stress in Mice

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
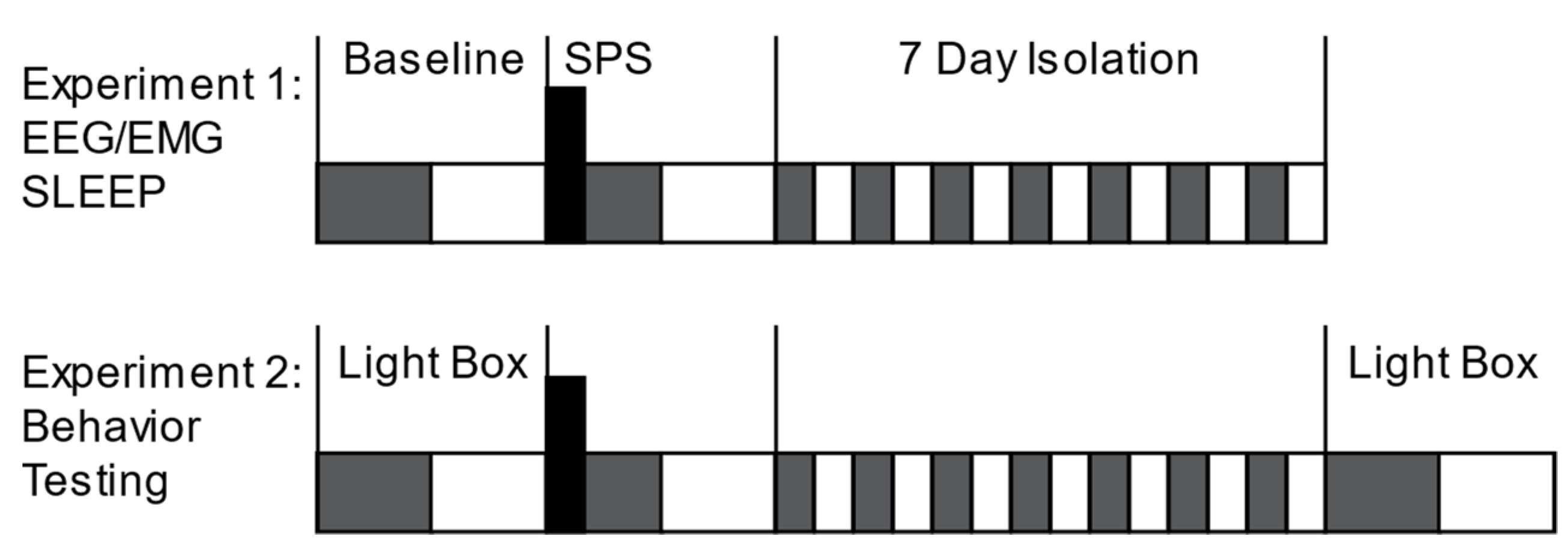
2.1. Animals and Surgery
2.2. Sleep Recording and Analysis
2.3. Single Prolonged Stress
2.4. Light–Dark Box Anxiety Testing
3. Results
3.1. Sleep–Wake Behavior
3.1.1. SPS Changes in Wakefulness in Fabp7 KO versus WT
3.1.2. SPS Changes in Sleep Stages between Fabp7 KO versus WT
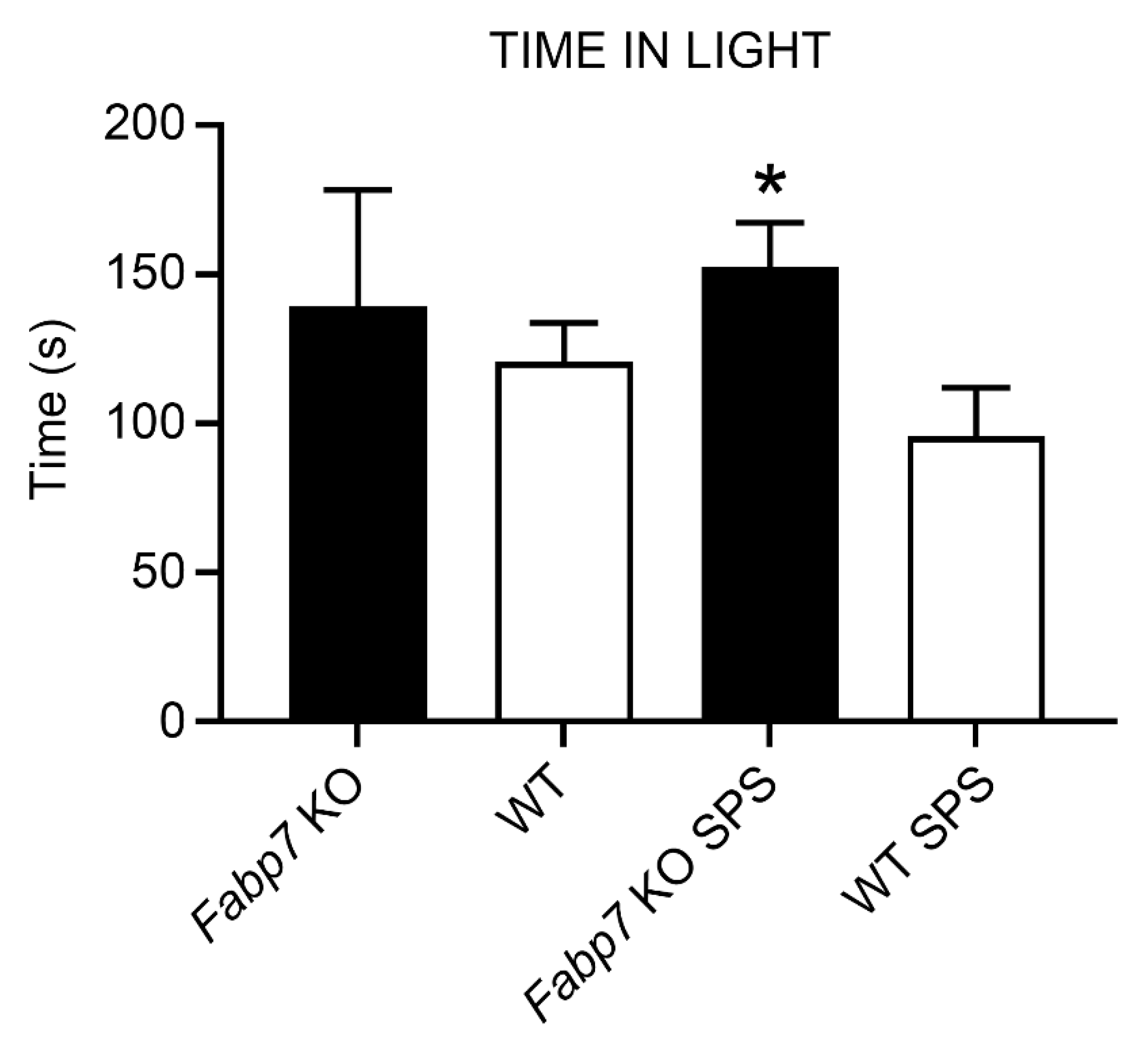
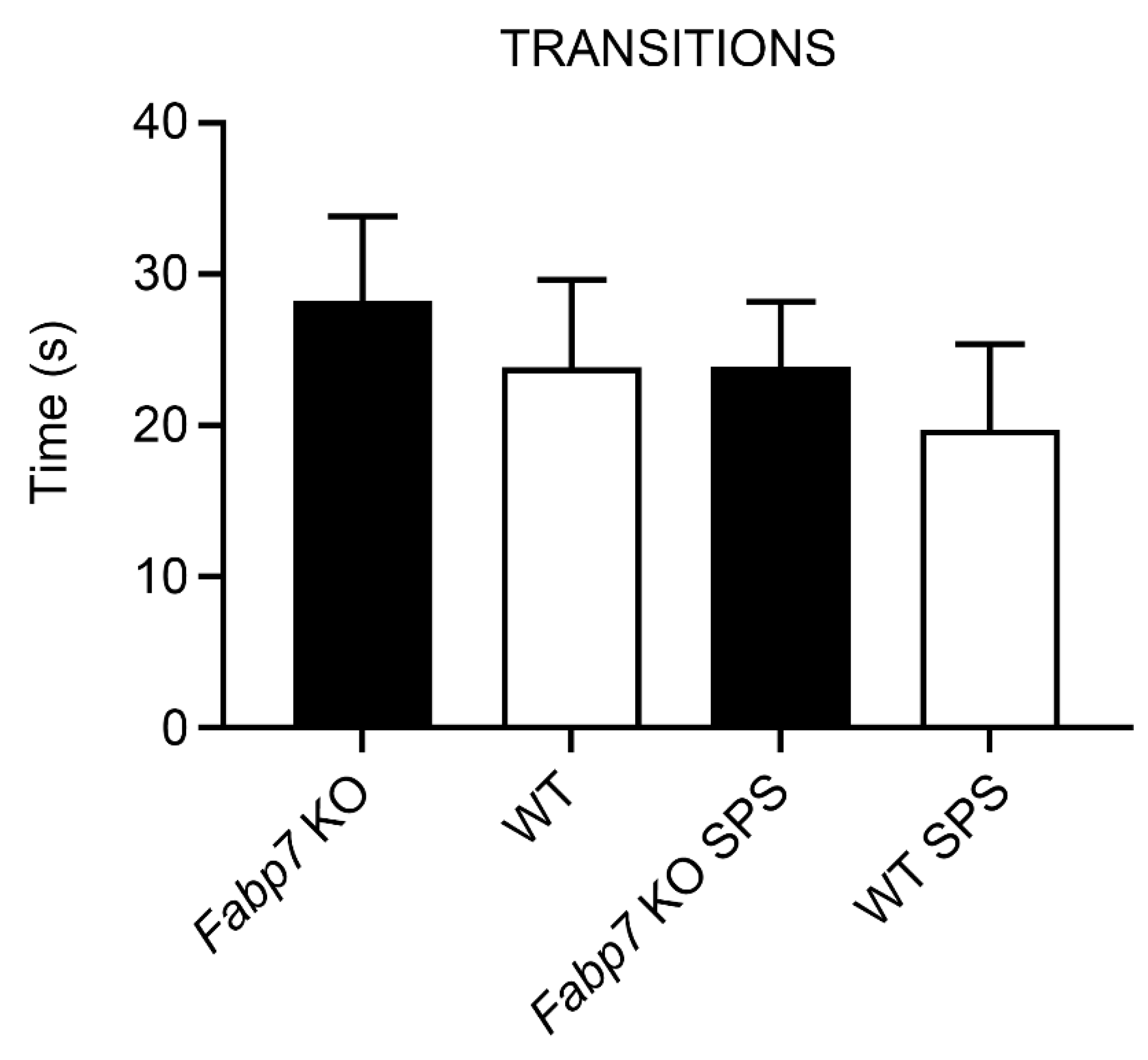
3.2. Light–Dark Box Anxiety Test
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

References
- Brown, T.H.; Mellman, T.A.; Alfano, C.A.; Weems, C.F. Sleep fears, sleep disturbance, and PTSD symptoms in minority youth exposed to Hurricane Katrina. J. Trauma. Stress 2011, 24, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Mellman, T.A.; David, D.; Kulick-Bell, R.; Hebding, J.; Nolan, B. Sleep disturbance and its relationship to psychiatric morbidity after Hurricane Andrew. Am. J. Psychiatry 1995, 152, 1659–1663. [Google Scholar] [PubMed]
- Mellman, T.A.; Bustamante, V.; Fins, A.I.; Pigeon, W.R.; Nolan, B. REM sleep and the early development of posttraumatic stress disorder. Am. J. Psychiatry 2002, 159, 1696–1701. [Google Scholar] [CrossRef] [PubMed]
- Vanderheyden, W.M.; Poe, G.R.; Liberzon, I. Trauma exposure and sleep: Using a rodent model to understand sleep function in PTSD. Exp. Brain Res. 2014, 232, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Knox, D.; Nault, T.; Henderson, C.; Liberzon, I. Glucocorticoid receptors and extinction retention deficits in the single prolonged stress model. Neuroscience 2012, 223, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Knox, D.; George, S.A.; Fitzpatrick, C.J.; Rabinak, C.A.; Maren, S.; Liberzon, I. Single prolonged stress disrupts retention of extinguished fear in rats. Learn. Mem. 2012, 19, 43–49. [Google Scholar] [CrossRef]
- Yamamoto, S.; Morinobu, S.; Takei, S.; Fuchikami, M.; Matsuki, A.; Yamawaki, S.; Liberzon, I. Single prolonged stress: Toward an animal model of posttraumatic stress disorder. Depress. Anxiety 2009, 26, 1110–1117. [Google Scholar] [CrossRef]
- Ferland-Beckham, C.; Chaby, L.E.; Daskalakis, N.P.; Knox, D.; Liberzon, I.; Lim, M.M.; McIntyre, C.; Perrine, S.A.; Risbrough, V.B.; Sabban, E.L.; et al. Systematic Review and Methodological Considerations for the Use of Single Prolonged Stress and Fear Extinction Retention in Rodents. Front. Behav. Neurosci. 2021, 15, 652636. [Google Scholar] [CrossRef]
- Perrine, S.A.; Eagle, A.L.; George, S.A.; Mulo, K.; Kohler, R.J.; Gerard, J.; Harutyunyan, A.; Hool, S.M.; Susick, L.L.; Schneider, B.L.; et al. Severe, multimodal stress exposure induces PTSD-like characteristics in a mouse model of single prolonged stress. Behav. Brain Res. 2016, 303, 228–237. [Google Scholar] [CrossRef]
- Wang, H.; Zuo, D.; He, B.; Qiao, F.; Zhao, M.; Wu, Y. Conditioned fear stress combined with single-prolonged stress: A new PTSD mouse model. Neurosci. Res. 2012, 73, 142–152. [Google Scholar] [CrossRef]
- Nedelcovych, M.T.; Gould, R.W.; Zhan, X.; Bubser, M.; Gong, X.; Grannan, M.; Thompson, A.T.; Ivarsson, M.; Lindsley, C.W.; Conn, P.J.; et al. A rodent model of traumatic stress induces lasting sleep and quantitative electroencephalographic disturbances. ACS Chem. Neurosci. 2015, 6, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Vanderheyden, W.M.; George, S.A.; Urpa, L.; Kehoe, M.; Liberzon, I.; Poe, G.R. Sleep alterations following exposure to stress predict fear-associated memory impairments in a rodent model of PTSD. Exp. Brain Res. 2015, 233, 2335–2346. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.J.; Vanderheyden, W.M. Optogenetic sleep enhancement improves fear-associated memory processing following trauma exposure in rats. Sci. Rep. 2020, 10, 18025. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, K.; Yabuki, Y.; Takahata, I.; Matsuo, K. Neurological mechanism and therapeutic strategy for posttraumatic stress disorders. Nihon Yakurigaku Zasshi 2018, 152, 194–201. [Google Scholar] [CrossRef][Green Version]
- Owada, Y.; Abdelwahab, S.A.; Kitanaka, N.; Sakagami, H.; Takano, H.; Sugitani, Y.; Sugawara, M.; Kawashima, H.; Kiso, Y.; Mobarakeh, J.I.; et al. Altered emotional behavioral responses in mice lacking brain-type fatty acid-binding protein gene. Eur. J. Neurosci. 2006, 24, 175–187. [Google Scholar] [CrossRef]
- Gerstner, J.R.; Vander Heyden, W.M.; Lavaute, T.M.; Landry, C.F. Profiles of novel diurnally regulated genes in mouse hypothalamus: Expression analysis of the cysteine and histidine-rich domain-containing, zinc-binding protein 1, the fatty acid-binding protein 7 and the GTPase, ras-like family member 11b. Neuroscience 2006, 139, 1435–1448. [Google Scholar] [CrossRef]
- Gerstner, J.R.; Bremer, Q.Z.; Vander Heyden, W.M.; Lavaute, T.M.; Yin, J.C.; Landry, C.F. Brain fatty acid binding protein (Fabp7) is diurnally regulated in astrocytes and hippocampal granule cell precursors in adult rodent brain. PLoS ONE 2008, 3, e1631. [Google Scholar] [CrossRef]
- Gerstner, J.R.; Smith, G.G.; Lenz, O.; Perron, I.J.; Buono, R.J.; Ferraro, T.N. BMAL1 controls the diurnal rhythm and set point for electrical seizure threshold in mice. Front. Syst. Neurosci. 2014, 8, 121. [Google Scholar] [CrossRef]
- Vanderheyden, W.M.; Fang, B.; Flores, C.C.; Jager, J.; Gerstner, J.R. The transcriptional repressor Rev-erbα regulates circadian expression of the astrocyte Fabp7 mRNA. Curr. Res. Neurobiol. 2021, 2, 100009. [Google Scholar] [CrossRef]
- Gerstner, J.R.; Vanderheyden, W.M.; Shaw, P.J.; Landry, C.F.; Yin, J.C. Cytoplasmic to nuclear localization of fatty-acid binding protein correlates with specific forms of long-term memory in Drosophila. Commun. Integr. Biol. 2011, 4, 623–626. [Google Scholar] [CrossRef]
- Gerstner, J.R.; Vanderheyden, W.M.; Shaw, P.J.; Landry, C.F.; Yin, J.C. Fatty-acid binding proteins modulate sleep and enhance long-term memory consolidation in Drosophila. PLoS ONE 2011, 6, e15890. [Google Scholar] [CrossRef] [PubMed]
- Gerstner, J.R.; Perron, I.J.; Riedy, S.M.; Yoshikawa, T.; Kadotani, H.; Owada, Y.; Van Dongen, H.P.A.; Galante, R.J.; Dickinson, K.; Yin, J.C.P.; et al. Normal sleep requires the astrocyte brain-type fatty acid binding protein FABP7. Sci. Adv. 2017, 3, e1602663. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.J.; Gerstner, J.R.; Vanderheyden, W.M. Single prolonged stress blocks sleep homeostasis and pre-trauma sleep deprivation does not exacerbate the severity of trauma-induced fear-associated memory impairments. PLoS ONE 2021, 16, e0243743. [Google Scholar] [CrossRef]
- Misslin, R.; Belzung, C.; Vogel, E. Behavioural validation of a light/dark choice procedure for testing anti-anxiety agents. Behav. Process. 1989, 18, 119–132. [Google Scholar] [CrossRef]
- Crawley, J.N. Exploratory behavior models of anxiety in mice. Neurosci. Biobehav. Rev. 1985, 9, 37–44. [Google Scholar] [CrossRef]
- Costall, B.; Jones, B.J.; Kelly, M.E.; Naylor, R.J.; Tomkins, D.M. Exploration of mice in a black and white test box: Validation as a model of anxiety. Pharmacol. Biochem. Behav. 1989, 32, 777–785. [Google Scholar] [CrossRef]
- Bourin, M.; Hascoët, M. The mouse light/dark box test. Eur. J. Pharmacol. 2003, 463, 55–65. [Google Scholar] [CrossRef]
- Ressler, K.J.; Mayberg, H.S. Targeting abnormal neural circuits in mood and anxiety disorders: From the laboratory to the clinic. Nat. Neurosci. 2007, 10, 1116–1124. [Google Scholar] [CrossRef]
- Yu, B.; Cui, S.Y.; Zhang, X.Q.; Cui, X.Y.; Li, S.J.; Sheng, Z.F.; Cao, Q.; Huang, Y.L.; Xu, Y.P.; Lin, Z.G.; et al. Different neural circuitry is involved in physiological and psychological stress-induced PTSD-like “nightmares” in rats. Sci. Rep. 2015, 5, 15976. [Google Scholar] [CrossRef]
- Miller, K.E.; Brownlow, J.A.; Gehrman, P.R. Sleep in PTSD: Treatment approaches and outcomes. Curr. Opin. Psychol. 2020, 34, 12–17. [Google Scholar] [CrossRef]
- Brownlow, J.A.; Harb, G.C.; Ross, R.J. Treatment of Sleep Disturbances in Post-Traumatic Stress Disorder: A Review of the Literature. Curr. Psychiatry Rep. 2015, 17, 41. [Google Scholar] [CrossRef] [PubMed]
- Lamarche, L.J.; De Koninck, J. Sleep disturbance in adults with posttraumatic stress disorder: A review. J. Clin. Psychiatry 2007, 68, 1257–1270. [Google Scholar] [CrossRef] [PubMed]
- Fenster, R.J.; Lebois, L.A.M.; Ressler, K.J.; Suh, J. Brain circuit dysfunction in post-traumatic stress disorder: From mouse to man. Nat. Rev. Neurosci. 2018, 19, 535–551. [Google Scholar] [CrossRef] [PubMed]
- Sheynin, J.; Liberzon, I. Circuit dysregulation and circuit-based treatments in posttraumatic stress disorder. Neurosci. Lett. 2017, 649, 133–138. [Google Scholar] [CrossRef]
- Amzica, F.; Massimini, M.; Manfridi, A. Spatial buffering during slow and paroxysmal sleep oscillations in cortical networks of glial cells in vivo. J. Neurosci. 2002, 22, 1042–1053. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.G. The Role of Glia in Sleep Regulation and Function. In Sleep-Wake Neurobiology and Pharmacology; Springer: Cham, Switzerland, 2018; pp. 83–96. [Google Scholar] [CrossRef]
- Halassa, M.M.; Haydon, P.G. Integrated brain circuits: Astrocytic networks modulate neuronal activity and behavior. Annu. Rev. Physiol. 2010, 72, 335–355. [Google Scholar] [CrossRef]
- Watanabe, A.; Toyota, T.; Owada, Y.; Hayashi, T.; Iwayama, Y.; Matsumata, M.; Ishitsuka, Y.; Nakaya, A.; Maekawa, M.; Ohnishi, T.; et al. Fabp7 maps to a quantitative trait locus for a schizophrenia endophenotype. PLoS Biol. 2007, 5, e297. [Google Scholar] [CrossRef]
- Li, Y.F.; Huang, Y.; Amsdell, S.L.; Xiao, L.; O’Donnell, J.M.; Zhang, H.T. Antidepressant- and anxiolytic-like effects of the phosphodiesterase-4 inhibitor rolipram on behavior depend on cyclic AMP response element binding protein-mediated neurogenesis in the hippocampus. Neuropsychopharmacology 2009, 34, 2404–2419. [Google Scholar] [CrossRef]
- Maher, A.; El Sayed, N.; Nafea, H.; Gad, M.Z. Rolipram rescues memory consolidation deficits caused by sleep deprivation: Implication of the cAMP/PKA and cAMP/Epac pathways. CNS Neurol. Disord. Drug Targets 2021, 21, 631–639. [Google Scholar] [CrossRef]
- Vecsey, C.G.; Baillie, G.S.; Jaganath, D.; Havekes, R.; Daniels, A.; Wimmer, M.; Huang, T.; Brown, K.M.; Li, X.Y.; Descalzi, G.; et al. Sleep deprivation impairs cAMP signalling in the hippocampus. Nature 2009, 461, 1122–1125. [Google Scholar] [CrossRef]
- Havekes, R.; Vecsey, C.G.; Abel, T. The impact of sleep deprivation on neuronal and glial signaling pathways important for memory and synaptic plasticity. Cell. Signal. 2012, 24, 1251–1260. [Google Scholar] [CrossRef]
- Han, F.; Xiao, B.; Wen, L. Loss of Glial Cells of the Hippocampus in a Rat Model of Post-traumatic Stress Disorder. Neurochem. Res. 2015, 40, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.E.; Paniccia, J.E.; Lebonville, C.L.; Reissner, K.J.; Lysle, D.T. Chemogenetic Manipulation of Dorsal Hippocampal Astrocytes Protects Against the Development of Stress-enhanced Fear Learning. Neuroscience 2018, 388, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.E.; Lebonville, C.L.; Paniccia, J.E.; Balentine, M.E.; Reissner, K.J.; Lysle, D.T. Hippocampal interleukin-1 mediates stress-enhanced fear learning: A potential role for astrocyte-derived interleukin-1β. Brain Behav. Immun. 2018, 67, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Gerstner, J.R.; Vanderheyden, W.M.; LaVaute, T.; Westmark, C.J.; Rouhana, L.; Pack, A.I.; Wickens, M.; Landry, C.F. Time of day regulates subcellular trafficking, tripartite synaptic localization, and polyadenylation of the astrocytic Fabp7 mRNA. J. Neurosci. 2012, 32, 1383–1394. [Google Scholar] [CrossRef]
- Kagawa, Y.; Yasumoto, Y.; Sharifi, K.; Ebrahimi, M.; Islam, A.; Miyazaki, H.; Yamamoto, Y.; Sawada, T.; Kishi, H.; Kobayashi, S.; et al. Fatty acid-binding protein 7 regulates function of caveolae in astrocytes through expression of caveolin-1. Glia 2015, 63, 780–794. [Google Scholar] [CrossRef]
- Cheng, A.; Jia, W.; Kawahata, I.; Fukunaga, K. A novel fatty acid-binding protein 5 and 7 inhibitor ameliorates oligodendrocyte injury in multiple sclerosis mouse models. EBioMedicine 2021, 72, 103582. [Google Scholar] [CrossRef]
- Stevens, H.E.; Jiang, G.Y.; Schwartz, M.L.; Vaccarino, F.M. Learning and memory depend on fibroblast growth factor receptor 2 functioning in hippocampus. Biol. Psychiatry 2012, 71, 1090–1098. [Google Scholar] [CrossRef]
- Graham, B.M.; Richardson, R. Memory of fearful events: The role of fibroblast growth factor-2 in fear acquisition and extinction. Neuroscience 2011, 189, 156–169. [Google Scholar] [CrossRef]
- Xia, L.; Zhai, M.; Wang, L.; Miao, D.; Zhu, X.; Wang, W. FGF2 blocks PTSD symptoms via an astrocyte-based mechanism. Behav. Brain Res. 2013, 256, 472–480. [Google Scholar] [CrossRef]
- Kipp, M.; Gingele, S.; Pott, F.; Clarner, T.; van der Valk, P.; Denecke, B.; Gan, L.; Siffrin, V.; Zipp, F.; Dreher, W.; et al. BLBP-expression in astrocytes during experimental demyelination and in human multiple sclerosis lesions. Brain Behav. Immun. 2011, 25, 1554–1568. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dark Phase | |||||
|---|---|---|---|---|---|
| WT | Fabp7 KO | ||||
| NREM | Mean | SEM | Mean | SEM | p-Value |
| BL | 1389.688 | 67.48 | 1349.896 | 69 | 0.680569 |
| SPS | 897.6042 | 82.37 | 830.1042 | 83.97 | 0.56674 |
| Day 7 | 1021.042 | 56.28 | 813.2143 | 65.74 | 0.016713 |
| REM | |||||
| BL | 144.5833 | 14.5 | 156.25 | 13.59 | 0.557893 |
| SPS | 75.72917 | 12.12 | 85.3125 | 12.56 | 0.583581 |
| Day 7 | 74.27083 | 8.361 | 73.80952 | 9.651 | 0.971073 |
| Light Phase | |||||
|---|---|---|---|---|---|
| WT | Fabp7 KO | ||||
| NREM | Mean | SEM | Mean | SEM | p-Value |
| BL | 1991.25 | 45.7 | 1880.521 | 53.82 | 0.118468 |
| SPS | 1883.854 | 46.98 | 1747.708 | 49.05 | 0.046434 |
| Day 7 | 1934.583 | 50.25 | 1873.214 | 49.07 | 0.386286 |
| REM | |||||
| BL | 306.5625 | 13.49 | 249.8958 | 12.77 | 0.002612 |
| SPS | 323.5417 | 12.89 | 275.8333 | 12.6 | 0.00879 |
| Day 7 | 299.8958 | 14.65 | 291.5476 | 15.68 | 0.697698 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vanderheyden, W.M.; Lefton, M.; Flores, C.C.; Owada, Y.; Gerstner, J.R. Fabp7 Is Required for Normal Sleep Suppression and Anxiety-Associated Phenotype following Single-Prolonged Stress in Mice. Neuroglia 2022, 3, 73-83. https://doi.org/10.3390/neuroglia3020005
Vanderheyden WM, Lefton M, Flores CC, Owada Y, Gerstner JR. Fabp7 Is Required for Normal Sleep Suppression and Anxiety-Associated Phenotype following Single-Prolonged Stress in Mice. Neuroglia. 2022; 3(2):73-83. https://doi.org/10.3390/neuroglia3020005
Chicago/Turabian StyleVanderheyden, William M., Micah Lefton, Carlos C. Flores, Yuji Owada, and Jason R. Gerstner. 2022. "Fabp7 Is Required for Normal Sleep Suppression and Anxiety-Associated Phenotype following Single-Prolonged Stress in Mice" Neuroglia 3, no. 2: 73-83. https://doi.org/10.3390/neuroglia3020005
APA StyleVanderheyden, W. M., Lefton, M., Flores, C. C., Owada, Y., & Gerstner, J. R. (2022). Fabp7 Is Required for Normal Sleep Suppression and Anxiety-Associated Phenotype following Single-Prolonged Stress in Mice. Neuroglia, 3(2), 73-83. https://doi.org/10.3390/neuroglia3020005