
Severity, Logging and Microsite Influence Post-Fire Regeneration of Maritime Pine

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
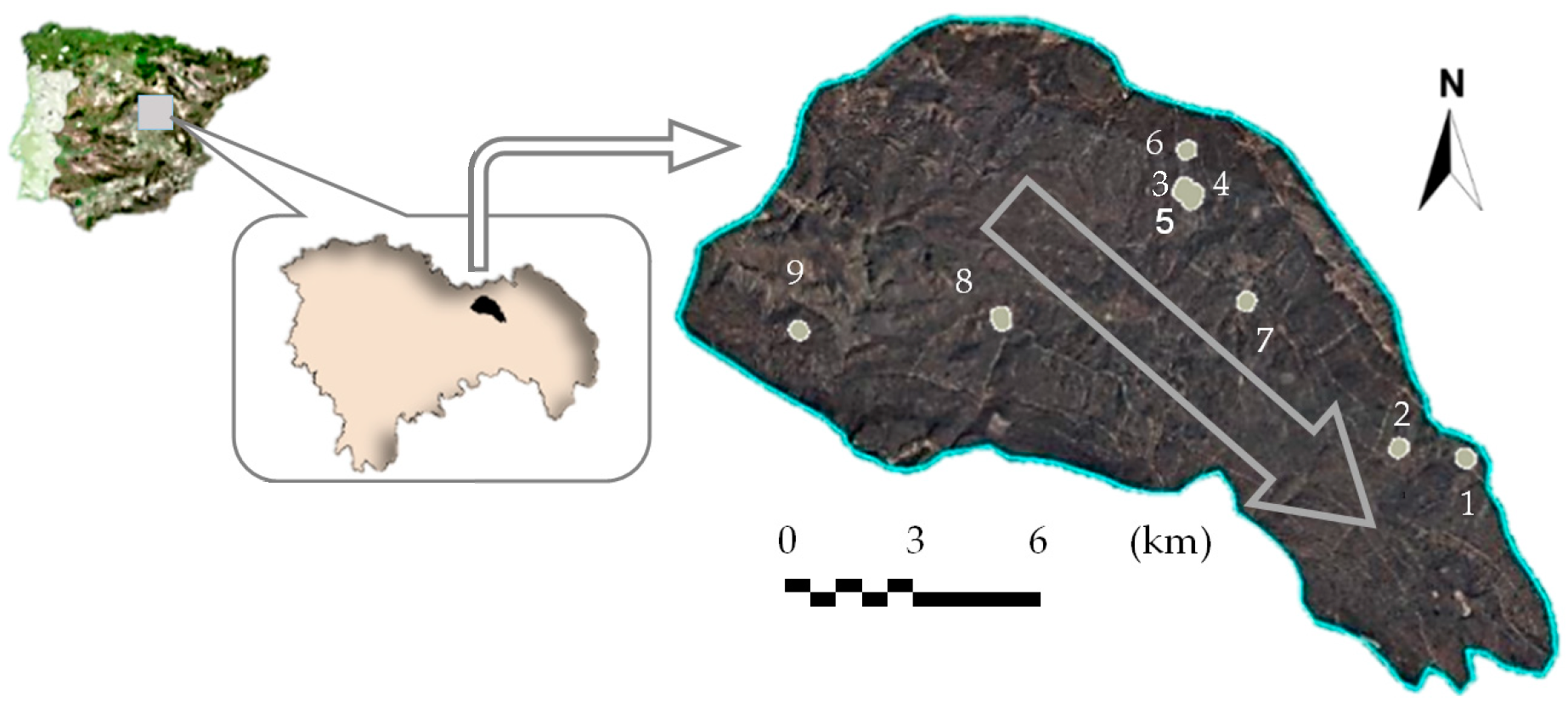
2.1. Study Area
2.2. Experimental Design and Field Sampling
2.3. Data Analysis
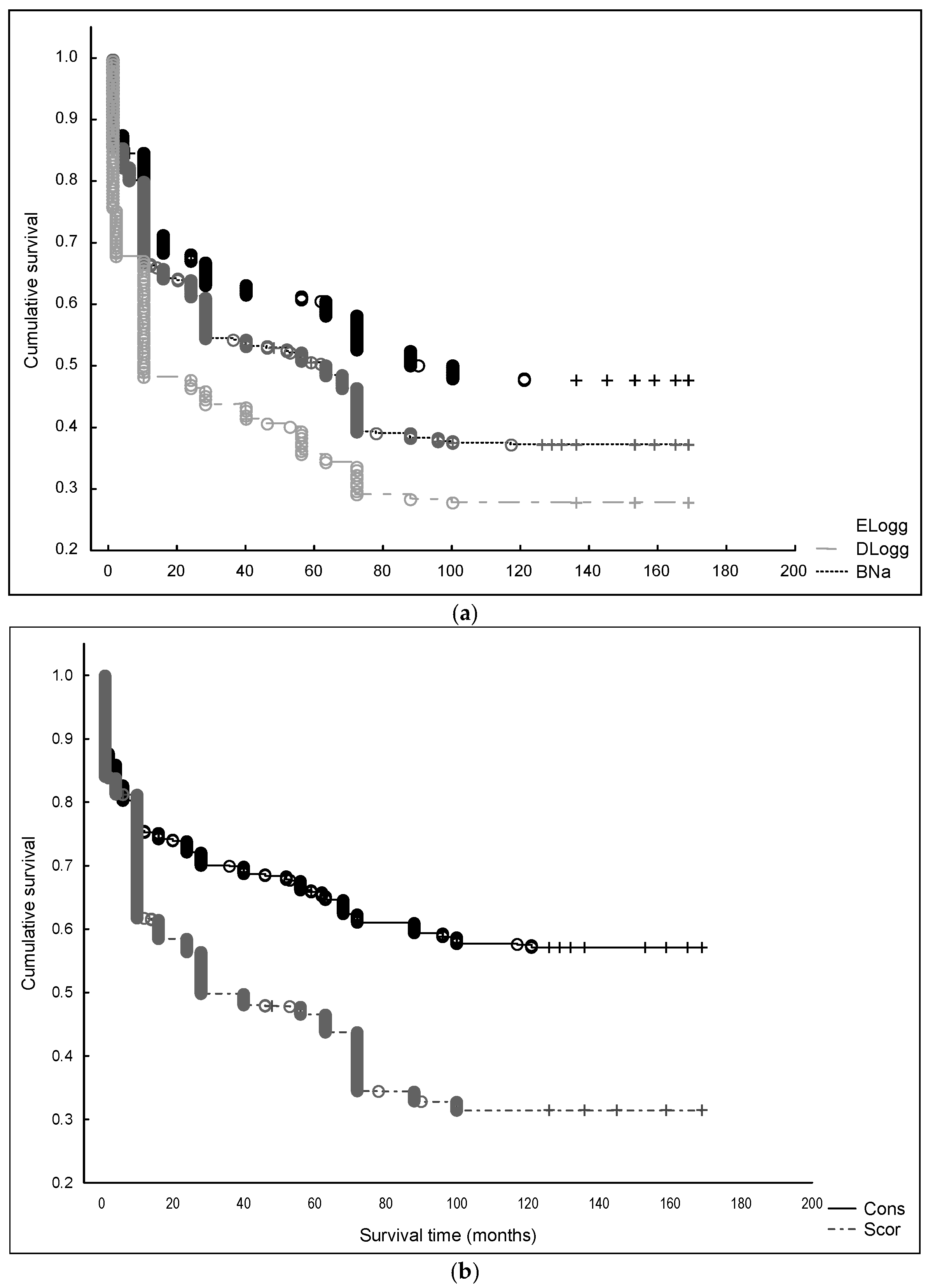
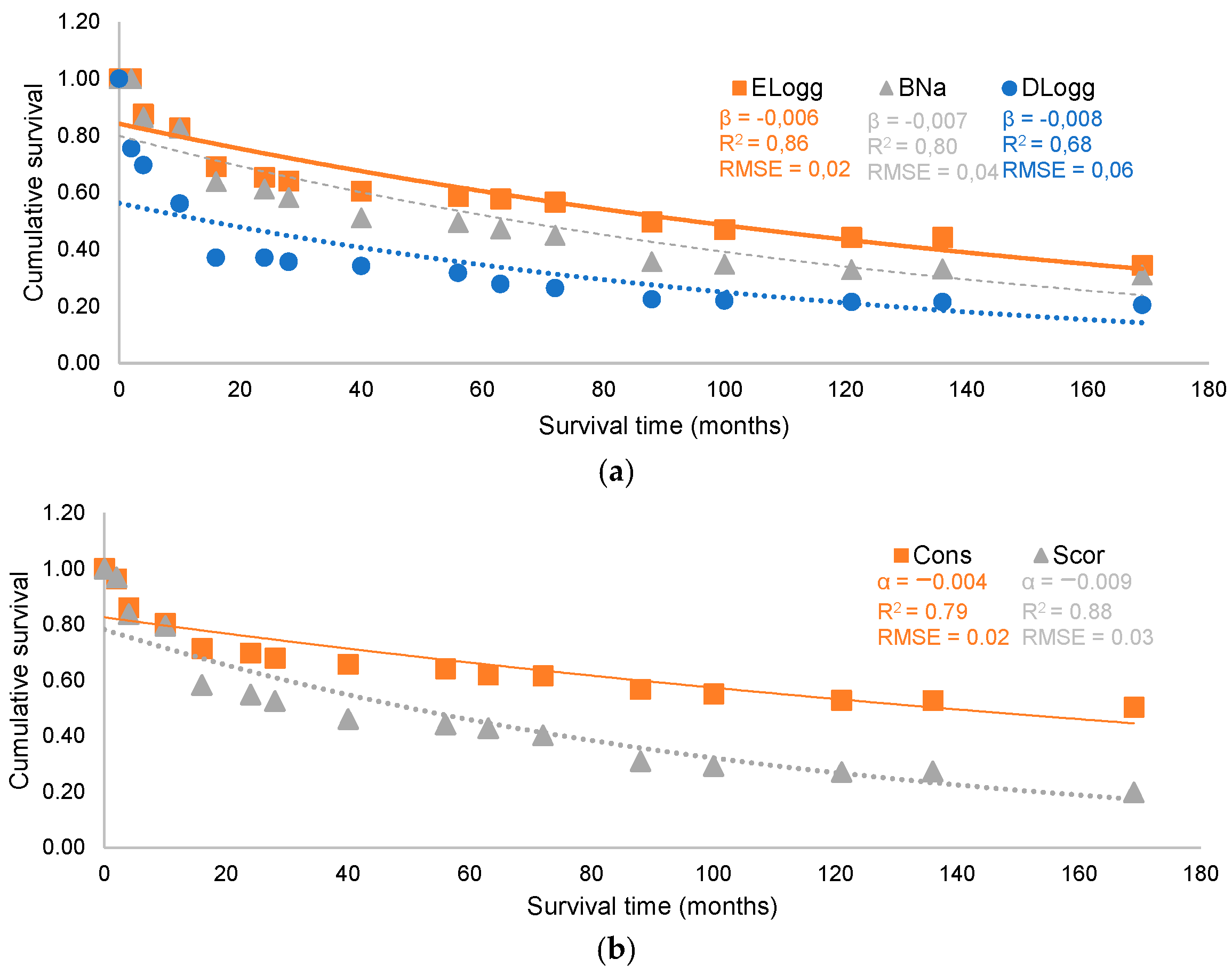
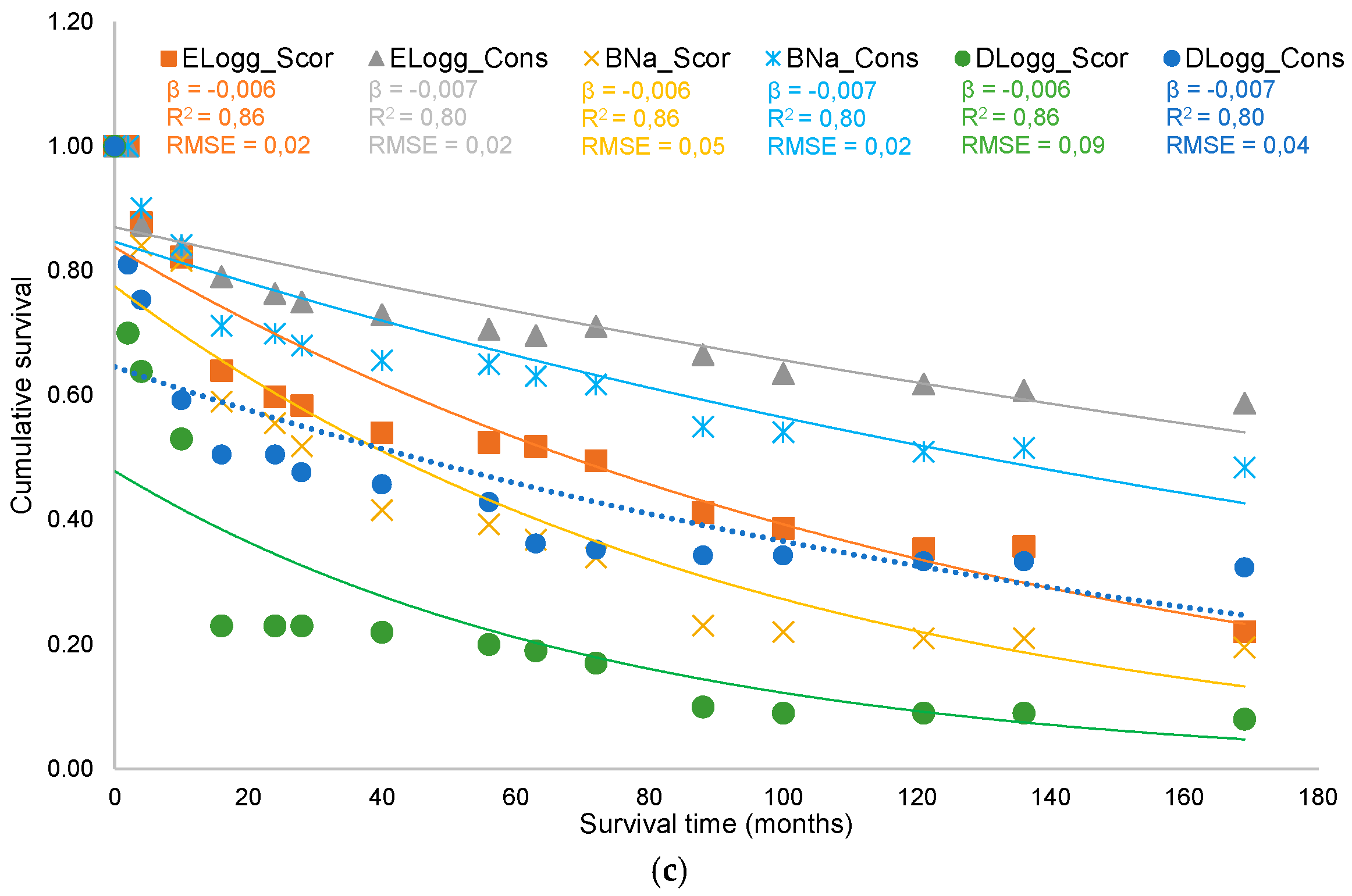
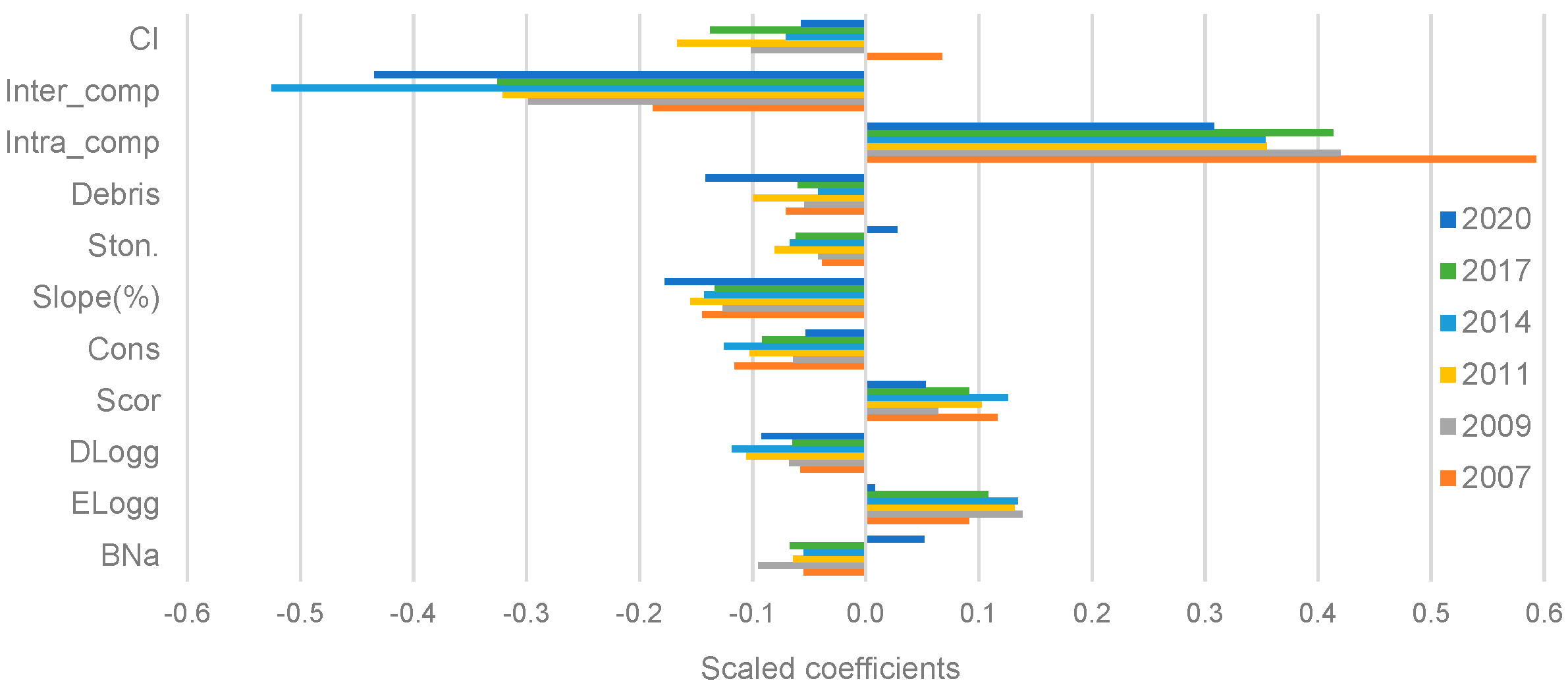
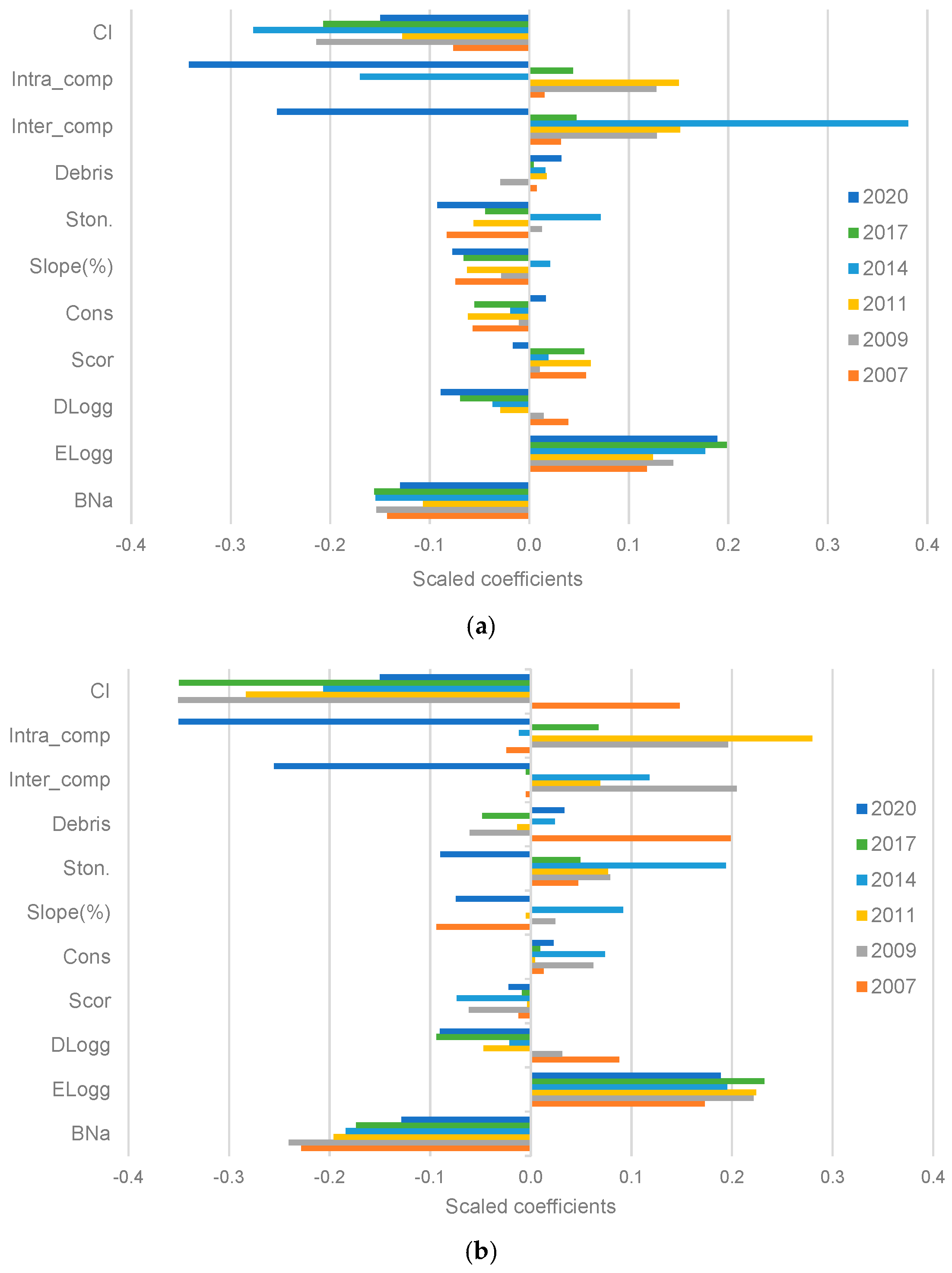
3. Results
3.1. Factors Influencing the Survival of Maritime Pine Seedlings: Stand and Microsite Scale
3.2. Factors Influencing the Regeneration Process: Stand Scale
4. Discussion
4.1. Factors Influencing the Survival of Maritime Pine Seedlings
4.2. Factors Influencing the Regeneration Process: Stand Scale
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pereira, P.; Francos, M.; Brevik, E.C.; Ubeda, X.; Bogunovic, I. Post-fire soil management. Curr. Opin. Environ. Sci. Health 2018, 5, 26–32. [Google Scholar] [CrossRef]
- Pellegrini, A.F.A.; Refsland, T.; Averill, C.; Terrer, C.; Staver, A.C.; Brockway, D.G.; Caprio, A.; Clatterbuck, W.; Coetsee, C.; Haywood, J.D.; et al. Decadal changes in fire frequencies shift tree communities and functional traits. Nat. Ecol. Evol. 2021, 5, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Yan, S.; He, L.; Shan, Y. Spatiotemporal changes in forest loss and its linkage to burned areas in China. J. For. Res. 2020, 31, 2525–2536. [Google Scholar] [CrossRef]
- Keeley, J.E.; Bond, W.J.; Bradstock, R.A.; Pausas, J.G.; Rundel, P.W. Fire in Mediterranean Ecosystems: Ecology, Evolution and Management; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar] [CrossRef]
- Dupuy, J.; Fargeon, H.; Martin-StPaul, N.; Pimont, F.; Ruffault, J.; Guijarro, M.; Hernando, C.; Madrigal, J.; Fernandes, P. Climate change impact on future wildfire danger and activity in southern Europe: A review. Ann. For. Sci. 2020, 77, 35. [Google Scholar] [CrossRef]
- Aquilué, N.; Fortin, M.J.; Messier, C.; Brotons, L. The potential of agricultural conversion to shape forest fire regimes in Mediterranean landscapes. Ecosystems 2020, 23, 34–51. [Google Scholar] [CrossRef]
- Madrigal, J.; Fernández-Migueláñez, I.; Hernando, C.; Guijarro, M.; Vega-Nieva, D.J.; Tolosana, E. Does forest biomass harvesting for energy reduce fire hazard in the Mediterranean basin? A case study in the Caroig Massif (Eastern Spain). Eur. J. For. Res. 2017, 136, 13–26. [Google Scholar] [CrossRef]
- García-Llamas, P.; Suárez-Seoane, S.; Taboada, A.; Fernández-Manso, A.; Quintano, C.; Fernández-García, V.; Fernández-Guisuraga, J.M.; Marcos, E.; Calvo, L. Environmental Drivers of Fire Severity in Extreme Fire Events That Affect Mediterranean Pine Forest Ecosystems. For. Ecol. Manag. 2019, 433, 24–32. [Google Scholar] [CrossRef]
- Rodrigues, M.; Cunill Camprubí, À.; Balaguer-Romano, R.; Coco Megía, C.J.; Castañares, F.; Ruffault, J.; Fernandes, P.M.; de Dios, V.R. Drivers and implications of the extreme 2022 wildfire season in Southwest Europe. Sci. Total Environ. 2023, 859, 160320. [Google Scholar] [CrossRef] [PubMed]
- Tedim, F.; Leone, V.; Amraoui, M.; Bouillon, C.; Coughlan, M.R.; Delogu, G.M.; Fernandes, P.M.; Ferreira, C.; McCaffrey, S.; McGee, T.K.; et al. Defining Extreme Wildfire Events: Difficulties, Challenges, and Impacts. Fire 2018, 1, 9. [Google Scholar] [CrossRef]
- Moya, D.; De Las Heras, J.; Ferrandis, F.; López-Serrano, F.R. Estimación de Cambios En La Comunidad Vegetal Tras Un Incendio Forestal. In Actualización de Métodos y Técnicas Para El Estudio de Los Suleos Afectados Por Incendios Forestales; Cerdá, A., Jordán, A., Eds.; Fuegored: Universitat de Valencia: Valencia, Spain, 2010; pp. 439–461. [Google Scholar]
- Pausas, J.; Llovet, J.; Rodrigo, A.; Vallejo, R. Are wildfires a disaster in the Mediterranean basin? A review. Int. J. Wildland Fire 2008, 17, 713. [Google Scholar] [CrossRef]
- Vega, J.A.; Fernández, C.; Pérez-Gorostiaga, P.; Fonturbel, T. Response of maritime pine (Pinus pinaster Ait.) recruitment to fire severity and post-fire management in a coastal burned area in Galicia (NW Spain). Plant Ecol. 2010, 206, 297–308. [Google Scholar] [CrossRef]
- Niccoli, F.; Esposito, A.; Altieri, S.; Battipaglia, G. Fire Severity Influences Ecophysiological Responses of Pinus pinaster Ait. Front. Plant Sci. 2019, 10, 448943. [Google Scholar] [CrossRef] [PubMed]
- Vega, J.A.; Fernández, C.; Pérez-Gorostiaga, P.; Fonturbel, T. The influence of fire severity, serotiny, and post-fire management on Pinus pinaster Ait. recruitment in three burnt areas in Galicia (NW Spain). For. Ecol. Manag. 2008, 256, 1596–1603. [Google Scholar] [CrossRef]
- Vega, J.A.; Fontúrbel, T.; Merino, A.; Fernández, C.; Ferreiro, A.; Jiménez, E. Testing the ability of visual indicators of soil burn severity to reflect changes in soil chemical and microbial properties in pine forests and shrubland. Plant Soil. 2013, 369, 73–91. [Google Scholar] [CrossRef]
- Fernández-García, V.; Miesel, J.; Baeza, M.J.; Marcos, E.; Calvo, L. Wildfire effects on soil properties in fire-prone pine ecosystems: Indicators of burn severity legacy over the medium term after fire. Appl. Soil Ecol. 2019, 135, 147–156. [Google Scholar] [CrossRef]
- Robichaud, P.R. After the smoke clears: Post-fire erosion and rehabilitation strategies in the United States. In FUEGORED 2010: International Workshop; Research and post-fire management soil protection and rehabilitation techniques for burnt forest ecosystems; Díaz Raviña, M., Benito, E., Carballas, T., Fontúrbel, M.T., Vega, J.A., Eds.; Consejo Superior de Investigaciones Científicas, CSIC, Santiago de Compostela: Santiago de Compostela, Spain, 2010; pp. 23–25. [Google Scholar]
- Vieira, D.C.S.; Fernández, C.; Vega, J.A.; Keizer, J.J. Does soil burn severity affect the post-fire runoff and interrill erosion response? A review based on meta-analysis of field rainfall simulation data. J. Hydrol. 2015, 523, 452–464. [Google Scholar] [CrossRef]
- Parente, J.; Girona-García, A.; Lopes, A.R.; Keizer, J.J.; Vieira, D.C.S. Prediction, validation, and uncertainties of a nation-wide post-fire soil erosion risk assessment in Portugal. Sci. Rep. 2022, 12, 2945. [Google Scholar] [CrossRef] [PubMed]
- Wolf, K.D.; Higuera, P.E.; Davis, K.T.; Dobrowski, S.Z. Wildfire impacts on forest microclimate vary with biophysical context. Ecosphere 2021, 12, e03467. [Google Scholar] [CrossRef]
- Fernández-Guisuraga, J.M.; Fernandes, P.M.; Tárrega, R.; Beltrán-Marcos, D.; Calvo, L. Vegetation recovery drivers at short-term after fire are plant community-dependent in mediterranean burned landscapes. For. Ecol. Manag. 2023, 539, 121034. [Google Scholar] [CrossRef]
- León-Sánchez, L.; Nicolás, E.; Prieto, I.; Nortes, P.; Maestre, F.T.; Querejeta, J.I. Altered leaf elemental composition with climate change is linked to reductions in photosynthesis, growth and survival in a semi-arid shrubland. J. Ecol. 2020, 108, 47–60. [Google Scholar] [CrossRef]
- Rodríguez-García, E.; Santana, V.M.; Alloza, J.A.; Ramón Vallejo, V. Predicting natural hyperdense regeneration after wildfires in Pinus halepensis (Mill.) forests using prefire site factors, forest structure and fire severity. For. Ecol. Manag. 2022, 512, 120164. [Google Scholar] [CrossRef]
- Tapias, R.; Climent, J.; Pardos, J.A.; Gil, L. Life histories of Mediterranean pines. Plant Ecol. 2004, 171, 53–68. [Google Scholar] [CrossRef]
- Vizcaíno-Palomar, N.; Revuelta-Eugercios, B.; Zavala, M.A.; Alía, R.; González-Martínez, S.C. The Role of Population Origin and Microenvironment in Seedling Emergence and Early Survival in Mediterranean Maritime Pine (Pinus pinaster Aiton). PLoS ONE 2014, 9, e109132. [Google Scholar] [CrossRef] [PubMed]
- Bhola, N.; Ogutu, J.O.; Dublin, H.T.; Van Der Plas, F.; Piepho, H.P.; Olff, H. Tree community structure and recruitment dynamics in savanna woodlands. Int. J. Biodivers. Conserv. 2020, 12, 71–103. [Google Scholar] [CrossRef]
- Vallejo, V.R.; Alloza, J.A. Chapter 12—Postfire Ecosystem Restoration. In Wildfire Hazards, Risks and Disasters; Shroder, J.F., Paton, D., Eds.; Elsevier: Oxford, UK, 2015; pp. 229–246. [Google Scholar] [CrossRef]
- Pereira, P.; Martínez-Murillo, J.F.; Francos, M. Chapter Four—Environments Affected by Fire. In Advances in Chemical Pollution, Environmental Management and Protection; Pereira, P., Ed.; Soil Degradation, Restoration and Management in a Global Change Context; Elsevier: Amsterdam, Netherlands, 2019; Volume 4, pp. 119–155. [Google Scholar] [CrossRef]
- Underwood, E.C.; Hollander, A.D.; Molinari, N.A.; Larios, L.; Safford, H.D. Identifying priorities for post-fire restoration in California chaparral shrublands. Rest. Ecol. 2022, 30, e13513. [Google Scholar] [CrossRef]
- Espinosa, J.; Dejene, T.; Guijarro, M.; Cerdá, X.; Madrigal, J.; Martín-Pinto, P. Fungal diversity and community composition responses to the reintroduction of fire in a non-managed Mediterranean shrubland ecosystem. For. Ecosyst. 2023, 10, 100110. [Google Scholar] [CrossRef]
- Vallejo, V.R.; Arianoutsou, M.; Moreira, F. Fire Ecology and Post-Fire Restoration Approaches in Southern European Forest Types. In Post-Fire Management and Restoration of Southern European Forests; Moreira, F., Arianoutsou, M., Corona, P., De las Heras, J., Eds.; Managing Forest Ecosystems; Springer: Dordrecht, The Netherlands, 2012; pp. 93–119. [Google Scholar] [CrossRef]
- Bravo, F.; Fabrika, M.; Ammer, C.; Barreiro, S.; Bielak, K.; Coll, L.; Fonseca, T.; Kangur, A.; Löf, M.; Merganičová, K.; et al. Modelling Approaches for Mixed Forests Dynamics Prognosis. Research Gaps and Opportunities. For. Syst. 2019, 28, eR002. [Google Scholar] [CrossRef]
- Moreira, F.; Ascoli, D.; Safford, H.; Adams, M.A.; Moreno, J.M.; Pereira, J.M.C.; Catry, F.X.; Armesto, J.; Bond, W.; González, M.E.; et al. Wildfire management in Mediterranean-type regions: Paradigm change needed. Environ. Res. Lett. 2020, 15, 011001. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Delgado-Baquerizo, M.; Muñoz-Rojas, M.; Plaza-Álvarez, P.A.; Gómez-Sanchez, M.E.; González-Romero, J.; Peña-Molina, E.; Moya, D.; de las Heras, J. Changes in ecosystem properties after post-fire management strategies in wildfire-affected Mediterranean forests. J. Appl. Ecol. 2021, 58, 836–846. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Van Stan, J.T., II.; Heydari, M.; Omidipour, R.; Rocha, F.; Plaza-Alvarez, P.A.; Zema, D.A.; Muñoz-Rojas, M. Post-fire restoration with contour-felled log debris increases early recruitment of spanish black pine (Pinus nigra arn. ssp. salzmannii) in Mediterranean forests. Rest. Ecol. 2021, 29, e13338. [Google Scholar] [CrossRef]
- Alfaro-Sánchez, R.; López-Serrano, F.R.; Rubio, E.; Moya, D.; Sánchez-Salguero, R.; De Las Heras, J. Biomass storage in low timber productivity Mediterranean forests managed after natural post-fire regeneration in south-eastern Spain. Eur. J. For. Res. 2014, 133, 793–807. [Google Scholar] [CrossRef]
- Thorn, S.; Bässler, C.; Brandl, R.; Burton, P.J.; Cahall, R.; Campbell, J.L.; Castro, J.; Choi, C.Y.; Cobb, T.; Donato, D.C.; et al. Impacts of salvage logging on biodiversity: A meta-analysis. J. Appl. Ecol. 2018, 55, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Thorn, S.; Chao, A.; Georgiev, K.B.; Müller, J.; Bässler, C.; Campbell, J.L.; Castro, J.; Chen, H.Y.H.; Choi, C.Y.; Cobb, T.P.; et al. Estimating retention benchmarks for salvage logging to protect biodiversity. Nat. Commun. 2020, 11, 4762. [Google Scholar] [CrossRef] [PubMed]
- Leverkus, A.B.; Gustafsson, L.; Lindenmayer, D.B.; Castro, J.; Rey Benayas, J.M.; Ranius, T.; Thorn, S. Salvage logging effects on regulating ecosystem services and fuel loads. Front. Ecol. Environ. 2020, 18, 391–400. [Google Scholar] [CrossRef]
- Juan-Ovejero, R.; Molinas-González, C.R.; Leverkus, A.B.; Martín Peinado, F.J.; Castro, J. Decadal effect of post-fire management treatments on soil carbon and nutrient concentrations in a burnt Mediterranean forest. For. Ecol. Manag. 2021, 498, 119570. [Google Scholar] [CrossRef]
- Castro, J.; Allen, C.D.; Molina-Morales, M.; Marañón-Jiménez, S.; Sánchez-Miranda, Á.; Zamora, R. Salvage logging versus the use of burnt wood as a nurse object to promote post-fire tree seedling establishment. Restor. Ecol. 2011, 19, 537–544. [Google Scholar] [CrossRef]
- Peterson, D.W.; Dodson, E.K.; Harrod, R.J. Post-fire logging reduces surface woody fuels up to four decades following wildfire. For. Ecol. Manag. 2015, 338, 84–91. [Google Scholar] [CrossRef]
- Stevens-Rumann, C.S.; Hudak, A.T.; Morgan, P.; Arnold, A.; Strand, E.K. Fuel Dynamics Following Wildfire in US Northern Rockies Forests. Front. For. Glob. Chang. 2020, 3, 51. [Google Scholar] [CrossRef]
- Calama Sainz, R.A.; Manso, R.; Lucas-Borja, M.E.; Espelta, J.M.; Piqué, M.; Bravo, F.; del Peso, C.; Pardos Mínguez, M. Natural regeneration in Iberian pines: A review of dynamic processes and proposals for management. For. Syst. 2017, 26, eR02S. [Google Scholar] [CrossRef]
- Carrillo-García, C.; Girola-Iglesias, L.; Guijarro, M.; Hernando, C.; Madrigal, J.; Mateo, R.G. Ecological niche models applied to post-megafire vegetation restoration in the context of climate change. Sci. Total Environ. 2023, 855, 158858. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Murillo, P.G.; Doña, V.J.; Pausas, J.G. Wildfires: Opportunity for restoration? Science 2019, 363, 134–135. [Google Scholar] [CrossRef] [PubMed]
- Berkey, J.K.; Belote, R.T.; Maher, C.T.; Larson, A.J. Structural diversity and development in active fire regime mixed-conifer forests. For. Ecol. Manag. 2021, 479, 118548. [Google Scholar] [CrossRef]
- Fernandes, P.M.; Rigolot, E. The fire ecology and management of maritime pine (Pinus pinaster Ait.). For. Ecol. Manag. 2007, 241, 1–13. [Google Scholar] [CrossRef]
- Pausas, J.G.; Verdú, M. Plant persistence traits in fire-prone ecosystems of the Mediterranean basin: A phylogenetic approach. Oikos 2005, 109, 196–202. [Google Scholar] [CrossRef]
- Cruz, O.; García-Duro, J.; Casal, M.; Reyes, O. Role of serotiny on Pinus pinaster Aiton germination and its relation to mother plant age and fire severity. iForest. 2019, 12, 491. [Google Scholar] [CrossRef]
- Alía, R.; Martín, S.; De Miguel, J.; Galera, R.; Agúndez, D.; Gordo, L.; Salvador, L.; Catalán, G.; Gil, L. Regiones de Procedencia de Pinus Pinaster Aiton; Dirección General de Conservación de la Naturaleza, Ministerio de Medio Ambiente: Madrid, Spain, 1996; 75p. [Google Scholar]
- Tapias, R.; Gil, L.; Fuentes-Utrilla, P.; Pardos, J.A. Canopy seed banks in Mediterranean pines of south-eastern Spain: A comparison between Pinus halepensis Mill., P. pinaster Ait., P. nigra Arn. and P. pinea L. J. Ecol. 2001, 89, 629–638. [Google Scholar] [CrossRef]
- Madrigal, J.; Hernando, C.; Guijarro, M.; Vega, J.A.; Fontúrbel, T.; Pérez-Gorostiaga, P. Smouldering fire-induced changes in a Mediterranean soil (SE Spain): Effects on germination, survival and morphological traits of 3-year-old Pinus pinaster Ait. Plant Ecol. 2010, 208, 279–292. [Google Scholar] [CrossRef]
- Batllori, E.; De Cáceres, M.; Brotons, L.; Ackerly, D.D.; Moritz, M.A.; Lloret, F. Compound fire-drought regimes promote ecosystem transitions in Mediterranean ecosystems. J. Ecol. 2019, 107, 1187–1198. [Google Scholar] [CrossRef]
- Fernández, C.; Vega, J.A.; Fonturbel, T.; Jiménez, E.; Pérez-Gorostiaga, P. Effects of wildfire, salvage logging and slash manipulation on Pinus pinaster Ait. recruitment in Orense (NW Spain). For. Ecol. Manag. 2008, 255, 1294–1304. [Google Scholar] [CrossRef]
- Taboada, A.; Tárrega, R.; Marcos, E.; Valbuena, L.; Suárez-Seoane, S.; Calvo, L. Fire recurrence and emergency post-fire management influence seedling recruitment and growth by altering plant interactions in fire-prone ecosystems. For. Ecol. Manag. 2017, 402, 63–75. [Google Scholar] [CrossRef]
- Ribeiro, S.; Cerveira, A.; Soares, P.; Fonseca, T. Natural Regeneration of Maritime Pine: A Review of the Influencing Factors and Proposals for Management. Forests 2022, 13, 386. [Google Scholar] [CrossRef]
- Del Campo, A.D.; Segura-Orenga, G.; Ceacero, C.J.; González-Sanchis, M.; Molina, A.J.; Reyna, S.; Hermoso, J. Reforesting drylands under novel climates with extreme drought filters: The importance of trait-based species selection. For. Ecol. Manag. 2020, 467, 118156. [Google Scholar] [CrossRef]
- Serrada Hierro, R.; Aguilar Parra, V.; Aroca Fernández, M.J.; Carrillo Patiño, A.; Ocaña Bueno, L. Estudio sobre la regeneración de las masas forestales afectadas por el incendio ocurrido el día 16 de julio en la comarca del Rodenal (Guadalajara). In V Congreso Forestal Español; Sociedad Española de Ciencias Forestales y Junta de Castilla y León: Ávila, Spain, 2009. [Google Scholar]
- Alía, R.; Moro, J. Comportamiento de procedencias de Pinus pinaster Ait. en el centro de España. Investig. Agrar. Sist. Recur. For. 1996, 5, 57–75. [Google Scholar]
- Varner, J.M.; Hood, S.M.; Aubrey, D.P.; Yedinak, K.; Hiers, J.K.; Jolly, W.M.; Shearman, T.M.; McDaniel, J.K.; O’Brien, J.J.; Rowell, E.M. Tree crown injury from wildland fires: Causes, measurement and ecological and physiological consequences. N. Phytol. 2021, 231, 1676–1685. [Google Scholar] [CrossRef] [PubMed]
- Brand, D.G. A competition index for predicting the vigour of planted Douglas-fir in southwestern British Columbia. Can. J. For. Res. 1986, 16, 23–29. [Google Scholar] [CrossRef]
- Weigelt, A.; Jolliffe, P. Indices of Plant Competition. J. Ecol. 2003, 91, 707–720. Available online: http://www.jstor.org/stable/3599697 (accessed on 19 April 2023). [CrossRef]
- Vilà, M.M.; Sardans, J. Plant competition in mediterranean-type vegetation. J. Veg. Sci. 2009, 10, 281–294. [Google Scholar] [CrossRef]
- Caryle, C.N.; Lauchlan, H.F.; Turkington, R. Using three pairs of competitive indices to test for changes in plant competition under different resource and disturbance levels. J. Veg. Sci. 2010, 21, 1025–1034. [Google Scholar] [CrossRef][Green Version]
- Kaplan, E.L.; Meier, P. Nonparametric estimation from incomplete observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Cox, D.R.; Oakes, D. Analysis of Survival Data; CRC Press: Boca Raton, FL, USA, 1984; Volume 21. [Google Scholar]
- De Jong, S. SIMPLS: An alternative approach to partial least squares regression. Chemom. Intell. Lab. Syst. 1993, 18, 251–263. [Google Scholar] [CrossRef]
- Castro, J.; Leverkus, A. Effect of Herbaceous Layer Interference on the Post-Fire Regeneration of a Serotinous Pine (Pinus pinaster Aiton) across Two Seedling Ages. Forests 2019, 10, 74. [Google Scholar] [CrossRef]
- Martínez-Sánchez, J.J.; Ferrandis, P.; de las Heras, J.; María Herranz, J. Effect of burnt wood removal on the natural regeneration of Pinus halepensis after fire in a pine forest in Tus valley (SE Spain). For. Ecol. Manag. 1999, 123, 1–10. [Google Scholar] [CrossRef]
- De Las Heras, J.; Martínez-Sánchez, J.; González-Ochoa, A.J.; Ferrandis, P.; Herranz, J. Desarrollo y mortalidad de plántulas de Pinus halepensis Mill. en competencia con Cistus monspeliensis L. durante los 4 primeros años post-incendio. Cuadernos SECF 2000, 10, 37–42. [Google Scholar]
- Matney, T.G.; Hodges, J.D. Evaluating Regeneration Success. In Forest Regeneration Manual; Duryea, M.L., Dougherty, P.M., Eds.; Forestry Sciences; Springer: Dordrecht, The Netherlands, 1991; pp. 321–331. [Google Scholar] [CrossRef]
- Cansler, C.A.; Kane, V.R.; Hessburg, P.F.; Kane, J.T.; Jeronimo, S.M.A.; Lutz, J.A.; Povak, N.A.; Churchill, D.J.; Larson, A.J. Previous wildfires and management treatments moderate subsequent fire severity. For. Ecol. Manag. 2022, 504, 119764. [Google Scholar] [CrossRef]
- Madrigal, J.; Hernando, C.; Martínez, E.; Guijarro, M.; Díez, C. Regeneración post-incendio de Pinus pinaster Ait. en la Sierra de Guadarrama (Sistema Central, España): Modelos descriptivos de los factores influyentes en la densidad inicial y la supervivencia. For. Syst. 2005, 14, 36–51. [Google Scholar]
- Santos, L.; Capelo, J.; Tavares, M. Germination Patterns of Soil Seed Banks in Relation to Fire in Portuguese Littoral Pine Forest Vegetation. Fire Ecol. 2010, 6, 1–15. [Google Scholar] [CrossRef]
- Martínez, J.L.; Lucas-Borja, M.E.; Plaza-Alvarez, P.A.; Denisi, P.; Moreno, M.A.; Hernández, D.; González-Romero, J.; Zema, D.A. Comparison of Satellite and Drone-Based Images at Two Spatial Scales to Evaluate Vegetation Regeneration after Post-Fire Treatments in a Mediterranean Forest. Appl. Sci. 2021, 11, 5423. [Google Scholar] [CrossRef]
- Fernández-Guisuraga, J.M.; Calvo, L.; Suárez-Seoane, S. Monitoring post-fire neighborhood competition effects on pine saplings under different environmental conditions by means of UAV multispectral data and structure-from-motion photogrammetry. J. Environ. Manag. 2022, 305, 114373. [Google Scholar] [CrossRef]
- Callaway, R.M.; Walker, L.R. Competition and facilitation: A synthetic approach to interactions in plant communities. Ecology 1997, 78, 1958–1965. [Google Scholar] [CrossRef]
- Coppoletta, M.; Merriam, K.E.; Collins, B.M. Post-fire vegetation and fuel development influences fire severity patterns in reburns. Ecol. Appl. 2016, 26, 686–699. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Z (m.a.s.l.) | Ho (m) | Denso (Trees ha−1) | Exp. (°) | Slope (%) | Ston. (%) | Depth (cm) | Sev. | Initial Emergence (Seedlings ha−1) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ELogg | BNa | DLogg | |||||||||
| 1 | 1368 | 9.0 | 587 | 270 | 33 | 89 | 26 | Cons | 4978 | 3911 | NA |
| 2 | 1205 | 8.9 | 612 | 290 | 6 | 25 | 38 | Scor | 68,622 | 43,022 | NA |
| 3 | 1238 | 8.7 | 933 | 88 | 33 | 33 | 44 | Cons | 1422 | 1422 | NA |
| 4 | 1238 | 12.7 | 1146 | 225 | 15 | 19 | 32 | Scor | 3022 | 1067 | NA |
| 5 | 1243 | 16.0 | 363 | 163 | 6 | 4 | 71 | Scor | 32,533 | NA | 20,978 |
| 6 | 1267 | 16.4 | 219 | 248 | 11 | 5 | 51 | Scor | 12,089 | 3378 | NA |
| 7 | 1248 | 16.0 | 62 | 130 | 2 | 5 | 51 | Scor | 8711 | 8355 | NA |
| 8 | 1244 | 16.2 | 512 | 76 | 32 | 68 | 48 | Cons | 533 | 3733 | 3200 |
| 9 | 1227 | 12.7 | 575 | 198 | 17 | 42 | 50 | Scor | 3022 | 20,800 | NA |
| Paired Interaction (A versus B) | Z Test | p-Value | |||
|---|---|---|---|---|---|
| (Treatment × Severity) A | (Treatment × Severity) B | ||||
| Early logging | Consumed | Burn No action | Consumed | 2.46 | 0.01 |
| Early logging | Consumed | Delayed logging | Consumed | 4.21 | 0.00 |
| Early logging | Scorched | Early logging | Consumed | 6.21 | 0.00 |
| Burn No action | Scorched | Early logging | Consumed | 8.92 | 0.00 |
| Delayed logging | Scorched | Early logging | Consumed | 8.34 | 0.00 |
| Delayed logging | Consumed | Burn No action | Consumed | 2.98 | 0.00 |
| Early logging | Scorched | Burn No action | Consumed | 3.25 | 0.00 |
| Burn No action | Consumed | Burn No action | Scorched | 6.18 | 0.00 |
| Burn No action | Consumed | Delayed logging | Scorched | 6.98 | 0.00 |
| Early logging | Scorched | Delayed logging | Consumed | −1.2 | 0.23 |
| Burn No action | Scorched | Delayed logging | Consumed | 0.65 | 0.52 |
| Delayed logging | Scorched | Delayed logging | Consumed | 2.94 | 0.00 |
| Early logging | Scorched | Burn No action | Scorched | 3.79 | 0.00 |
| Delayed logging | Scorched | Early logging | Scorched | 6.07 | 0.00 |
| Burn No action | Scorched | Delayed logging | Scorched | 4.3 | 0.00 |
| Variable | 4 B | 5 SE | 6 Wald | 7 Sig. | 8 Exp (B) | 95.0% Confidence Interval for Exp (B) | |
|---|---|---|---|---|---|---|---|
| Lower | Upper | ||||||
| Woody debris | −0.244 | 0.358 | 0.464 | 0.496 | 0.784 | 0.389 | 1.580 |
| 1 Inter_comp_09 | 0.472 | 0.239 | 3.883 | 0.049 | 1.603 | 1.003 | 2.563 |
| 2 Intra_comp_11 | −0.444 | 0.202 | 4.836 | 0.028 | 0.642 | 0.432 | 0.953 |
| 3 CI_11 | 0.040 | 0.043 | 0.871 | 0.351 | 1.041 | 0.957 | 1.132 |
| Slope (%) | −0.010 | 0.010 | 0.903 | 0.342 | 0.990 | 0.971 | 1.010 |
| Stoniness | −2.987 | 0.509 | 34.446 | 0.000 | 0.050 | 0.019 | 0.137 |
| Year | 1 h (m) | 2 bd (cm) | 3 Gral_comp | 4 h_comp (m) | 5 Inter_comp | 6 Intra_comp | 7 CI |
|---|---|---|---|---|---|---|---|
| 2007 | 0.14 (±0.08) | 0.4 (±0.6) | 0.58 (±0.49) | 0.21 (±0.24) | 0.56 (±0.49) | 0.03 (±0.18) | 1.19 (±2.10) |
| 2009 | 0.27 (±0.16) | 0.7 (±0.9) | 0.73 (±0.44) | 0.72 (±0.47) | 0.68 (±0.46) | 0.08 (±0.26) | 3.25 (±2.89) |
| 2011 | 0.39 (±0.22) | 0.9 (±0.6) | 0.77 (±0.42) | 0.77 (±0.47) | 0.71 (±0.45) | 0.20 (±0.40) | 2.30 (±1.70) |
| 2014 | 0.54 (±0.33) | 1.5 (±1.1) | 0.81 (±0.39) | 0.99 (±0.52) | 0.73 (±0.44) | 0.23 (±0.42) | 2.23 (±1.71) |
| 2017 | 0.82 (±0.52) | 2.2 (±1.8) | 0.88 (±0.32) | 1.13 (±0.59) | 0.74 (±0.43) | 0.29 (±0.45) | 1.65 (±1.13) |
| 2020 | 1.11 (±0.64) | 3.3 (±2.5) | 0.93 (±0.25) | 1.26 (±0.58) | 0.83 (±0.37) | 0.33 (±0.46) | 1.40 (±1.04) |
| Year | Dependent Variable | R2Y cum. | R2X cum. | RMSE | Highest Absolute Value Scaled Coefficient | |
|---|---|---|---|---|---|---|
| Value | Variable | |||||
| 2007 | 1 Dens | 0.74 | 0.52 | 1.83 | 0.59 | 4 Intra_comp |
| 2 h | 0.2 | 0.46 | 0.06 | −0.14 | Burn No action | |
| 3 Ø | 0.12 | −0.23 | Burn No action | |||
| 2009 | Dens | 0.6 | 0.49 | 1.28 | 0.42 | Intra_comp |
| h | 0.36 | 0.5 | 0.09 | −0.21 | 5 CI | |
| Ø | 0.06 | −0.35 | CI | |||
| 2011 | Dens | 0.57 | 0.52 | 1.05 | 0.35 | Intra_comp |
| h | 0.38 | 0.54 | 0.09 | 0.15 | 6 Inter_comp | |
| Ø | 0.07 | −0.28 | CI | |||
| 2014 | Dens | 0.69 | 0.47 | 0.95 | −0.53 | Inter_comp |
| h | 0.43 | 0.48 | 0.04 | −0.28 | CI | |
| Ø | 0.04 | 0.21 | CI | |||
| 2017 | Dens | 0.68 | 0.49 | 1.54 | 0.41 | Intra_comp |
| h | 0.36 | 0.49 | 0.07 | −0.21 | CI | |
| Ø | 0.07 | −0.35 | CI | |||
| 2020 | Dens | 0.58 | 0.45 | 0.97 | −0.43 | Inter_comp |
| h | 0.43 | 0.48 | 0.14 | −0.34 | Intra_comp | |
| Ø | 0.22 | −0.35 | Intra_comp | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrillo-García, C.; Hernando, C.; Díez, C.; Guijarro, M.; Madrigal, J. Severity, Logging and Microsite Influence Post-Fire Regeneration of Maritime Pine. Fire 2024, 7, 125. https://doi.org/10.3390/fire7040125
Carrillo-García C, Hernando C, Díez C, Guijarro M, Madrigal J. Severity, Logging and Microsite Influence Post-Fire Regeneration of Maritime Pine. Fire. 2024; 7(4):125. https://doi.org/10.3390/fire7040125
Chicago/Turabian StyleCarrillo-García, Cristina, Carmen Hernando, Carmen Díez, Mercedes Guijarro, and Javier Madrigal. 2024. "Severity, Logging and Microsite Influence Post-Fire Regeneration of Maritime Pine" Fire 7, no. 4: 125. https://doi.org/10.3390/fire7040125
APA StyleCarrillo-García, C., Hernando, C., Díez, C., Guijarro, M., & Madrigal, J. (2024). Severity, Logging and Microsite Influence Post-Fire Regeneration of Maritime Pine. Fire, 7(4), 125. https://doi.org/10.3390/fire7040125