Using a Multi-Century Post-Fire Chronosequence to Develop Criteria to Distinguish Prior and Bowman’s (2020) Post-Fire Obligate Coloniser and Fire-Intolerant Flora



Abstract
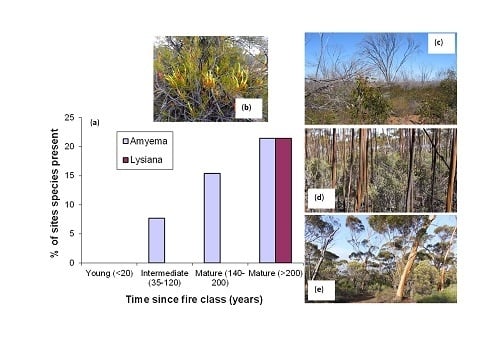
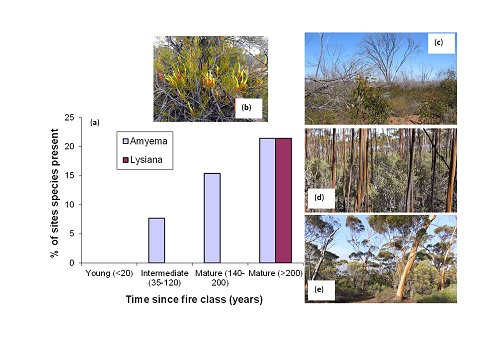
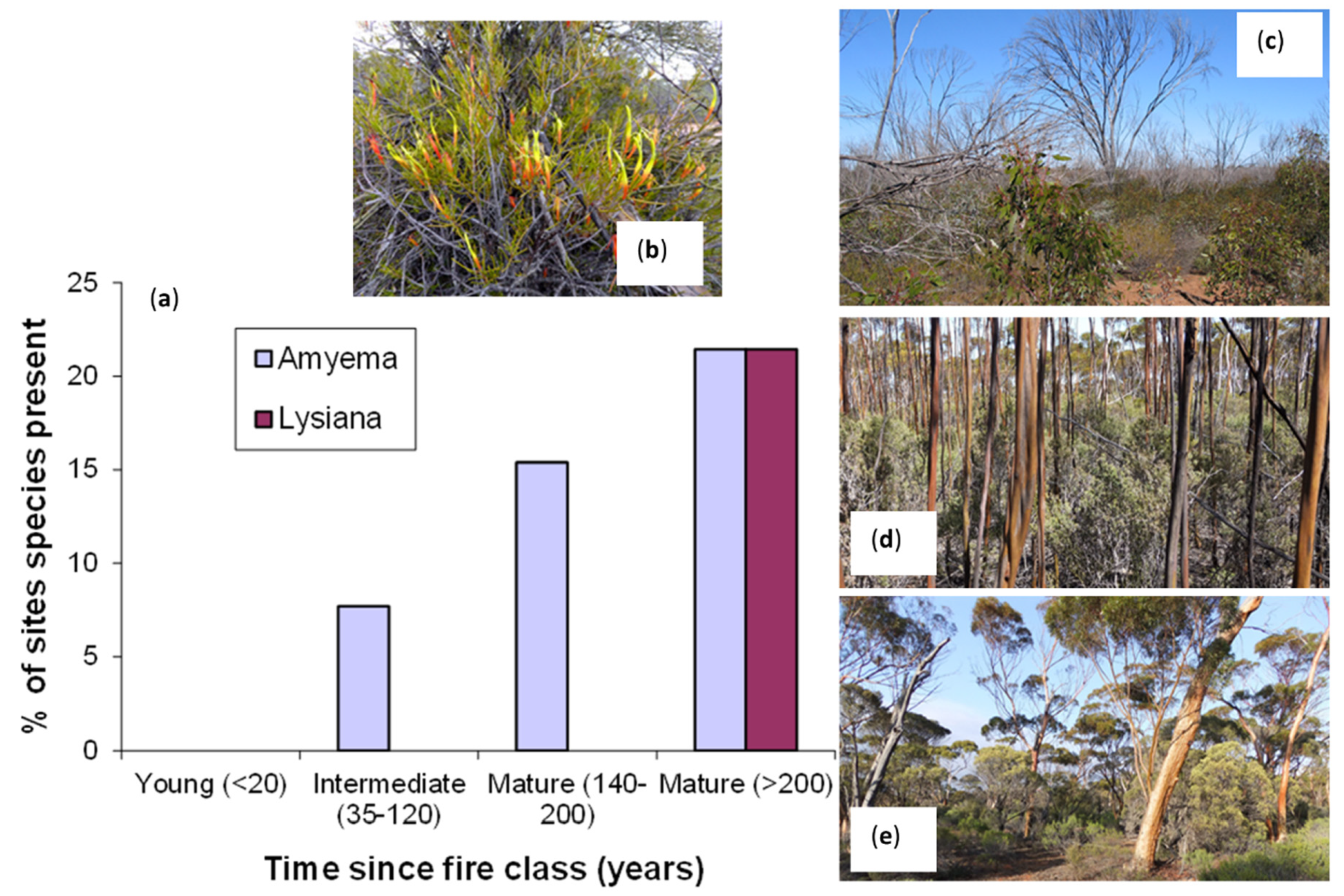
:
{kind=link}
{kind=link}
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Prior, L.D.; Bowman, D.M.J.S. Classification of post-fire responses of woody plants to include pyrophobic communities. Fire 2020, 3, 15. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J. Evolutionary ecology of resprouting and seeding in fire-prone ecosystems. New Phytol. 2014, 204, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.J.; Lawes, M.; Murphy, B.P.; Russell-Smith, J.; Nano, C.E.M.; Bradstock, R.; Enright, N.J.; Fontaine, J.B.; Gosper, C.R.; Radford, I.; et al. A synthesis of postfire recovery traits of woody plants in Australian ecosystems. Sci. Total Environ. 2015, 534, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Cochrane, M.A. Fire science for rainforests. Nature 2003, 421, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Holz, A.; Wood, S.W.; Veblen, T.T.; Bowman, D.M. Effects of high-severity fire drove the population collapse of the subalpine Tasmanian endemic conifer Athrotaxis cupressoides. Glob. Change Biol. 2015, 21, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Kitzberger, T.; Perry, G.L.W.; Paritsis, J.; Gowda, J.H.; Tepley, A.J.; Holz, A.; Veblen, T.T. Fire–vegetation feedbacks and alternative states: Common mechanisms of temperate forest vulnerability to fire in southern South America and New Zealand. N. Z. J. Bot. 2016, 54, 247–272. [Google Scholar] [CrossRef] [Green Version]
- Gosper, C.R.; Yates, C.J.; Prober, S.M. Floristic diversity in fire-sensitive eucalypt woodlands shows a ‘U’-shaped relationship with time since fire. J. Appl. Ecol. 2013, 50, 1187–1196. [Google Scholar] [CrossRef]
- O’Donnell, A.J.; Boer, M.M.; McCaw, W.L.; Grierson, P.F. Vegetation and landscape connectivity control wildfire intervals in unmanaged semi-arid shrublands and woodlands in Australia. J. Biogeogr. 2011, 38, 112–124. [Google Scholar] [CrossRef]
- Gosper, C.R.; Prober, S.M.; Yates, C.J. Multi-century changes in vegetation structure and fuel availability in fire-sensitive eucalypt woodlands. For. Ecol. Manag. 2013, 310, 102–109. [Google Scholar] [CrossRef]
- Gosper, C.R.; Prober, S.M.; Yates, C.J. Continental-scale syntheses of Australian pyromes—Misclassification of south-western eucalypt woodlands misinforms management. J. Biogeogr. 2016, 43, 858–861. [Google Scholar] [CrossRef]
- Gosper, C.; Prober, S.; Yates, C. Changes in Plant Diversity Indices, Composition and Cover in Eucalyptus salubris Woodlands Across Time Since Fire Chronosequence, Great Western Woodlands SuperSite. TERN Australian SuperSite Network. 2012. Available online: http://www.tern-supersites.net.au/knb/metacat/lloyd.276.20/html (accessed on 3 September 2020).
- Start, A.N. The mistletoe flora of southern Western Australia, with a particular reference to host relationships and fire. Aust. J. Bot. 2015, 63, 636–646. [Google Scholar] [CrossRef]
- Miller, B.P.; Dixon, K.W. Plants and fire in kwongan vegetation. In Plant Life on the Sandplains in Southwest Australia: A Global Biodiversity Hotspot; Lambers, H., Ed.; University of Western Australia Publishing: Perth, Australia, 2014; pp. 147–170. [Google Scholar]
- Ward, M.J.; Paton, D.C. Predicting mistletoe seed shadow and patterns of seed rain from movements of the mistletoebird, Dicaeum hirundinaceum. Austral Ecol. 2007, 32, 113–121. [Google Scholar] [CrossRef]
- Rawsthorne, J.; Watson, D.M.; Roshier, D.A. The restricted seed rain of a mistletoe specialist. J. Avian Biol. 2012, 43, 9–14. [Google Scholar] [CrossRef]
- McCaw, L.; Reynen, V.; Zdunic, K.; Peace, M. Reconstructing the spread of landscape-scale fires in semiarid southwestern Australia. In Advances in Forest Fire Research; Viegas, D.X., Ed.; Imprensa da Universidade de Coimbra: Coimbra, Portugal, 2014; pp. 912–920. [Google Scholar]
- Gosper, C.R.; Yates, C.J.; Cook, G.D.; Harvey, J.M.; Liedloff, A.C.; McCaw, W.L.; Thiele, K.R.; Prober, S.M. A conceptual model of vegetation dynamics for the unique obligate-seeder eucalypt woodlands of south-western Australia. Austral Ecol. 2018, 43, 681–695. [Google Scholar] [CrossRef]
- Vivian, L.M.; Doherty, M.D.; Cary, G.J. Classifying the fire-response traits of plants: How reliable are species-level classifications? Austral Ecol. 2010, 35, 264–273. [Google Scholar] [CrossRef]
- Gosper, C.R.; Fox, E.; Burbidge, A.H.; Craig, M.D.; Douglas, T.K.; Fitzsimons, J.A.; McNee, S.; Nicholls, A.O.; O’Connor, J.; Prober, S.M.; et al. Multi-century periods since fire in an intact woodland landscape favour bird species declining in an adjacent agricultural region. Biol. Cons. 2019, 230, 82–90. [Google Scholar] [CrossRef]
- Gallagher, R. Interim National Prioritisation of Australian Plants Affected by the 2019–2020 Bushfire Season. Research for the Wildlife and Threatened Species Bushfire Recovery Expert Panel Version 1.3. 2020. Available online: http://www.environment.gov.au/biodiversity/bushfire-recovery/priority-plants (accessed on 24 June 2020).

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gosper, C.R.; Prober, S.M. Using a Multi-Century Post-Fire Chronosequence to Develop Criteria to Distinguish Prior and Bowman’s (2020) Post-Fire Obligate Coloniser and Fire-Intolerant Flora. Fire 2020, 3, 48. https://doi.org/10.3390/fire3030048
Gosper CR, Prober SM. Using a Multi-Century Post-Fire Chronosequence to Develop Criteria to Distinguish Prior and Bowman’s (2020) Post-Fire Obligate Coloniser and Fire-Intolerant Flora. Fire. 2020; 3(3):48. https://doi.org/10.3390/fire3030048
Chicago/Turabian StyleGosper, Carl R., and Suzanne M. Prober. 2020. "Using a Multi-Century Post-Fire Chronosequence to Develop Criteria to Distinguish Prior and Bowman’s (2020) Post-Fire Obligate Coloniser and Fire-Intolerant Flora" Fire 3, no. 3: 48. https://doi.org/10.3390/fire3030048