How Much Does Fire Behavior of Leaf Litter Beds Change within Two Months?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
- (i)
- Resin rich, relatively long and thick (compared to broadleaved species) needles of P. halepensis would create leaf litter beds with low initial BD which would burn with high, fast spreading flames, low flaming duration and high fuel consumption, regardless of the treatment.
- (ii)
- Due to a large proportion of dense and thick leaf stems in the leaf litter and leaflet lamina thicker than those of Q. pubescens leaves, C. siliqua would create leaf litter beds with high initial bulk density. These fuel beds would burn with small, slow spreading flames, long flaming duration and low fuel consumption regardless of the treatment.
- (iii)
- Larger, thinner, lobed leaves of Q. pubescens would initially (“0” treatment) create leaf litter beds with low BD, which would burn similarly to P. halepensis litter beds.
- (iv)
- Due to low particle thickness and density Q. pubescens leaf litter would be susceptible to compaction, resulting in a pronounced increase of the BD in the treatments “10” and “60” compared to the treatment “0”.
- (v)
- The expected increase in BD of Q. pubescens would translate into alterations in the fire behavior, resulting in small, slow spreading flames and long flaming duration (i.e., in a fire behavior comparable to that of C. siliqua) in the treatment “60”.
2. Materials and Methods
2.1. Leaf Litter Handling
2.2. Experimental Set-Up
2.3. Fire Behavior Testing
2.4. Fuel Characteristics Measurements
2.5. Data Analysis
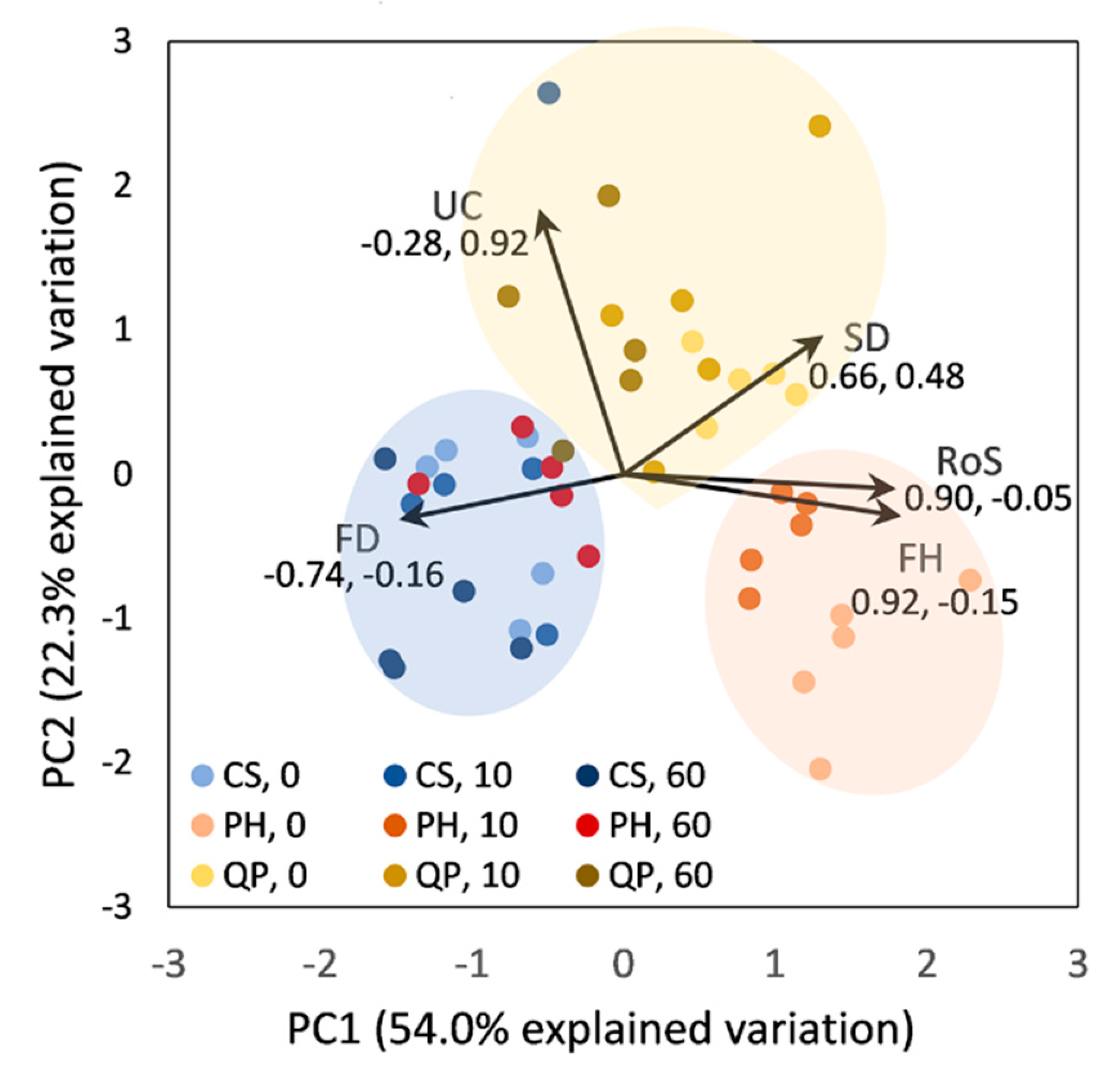
3. Results
3.1. Fire Behavior Characteristics and Grouping of Samples
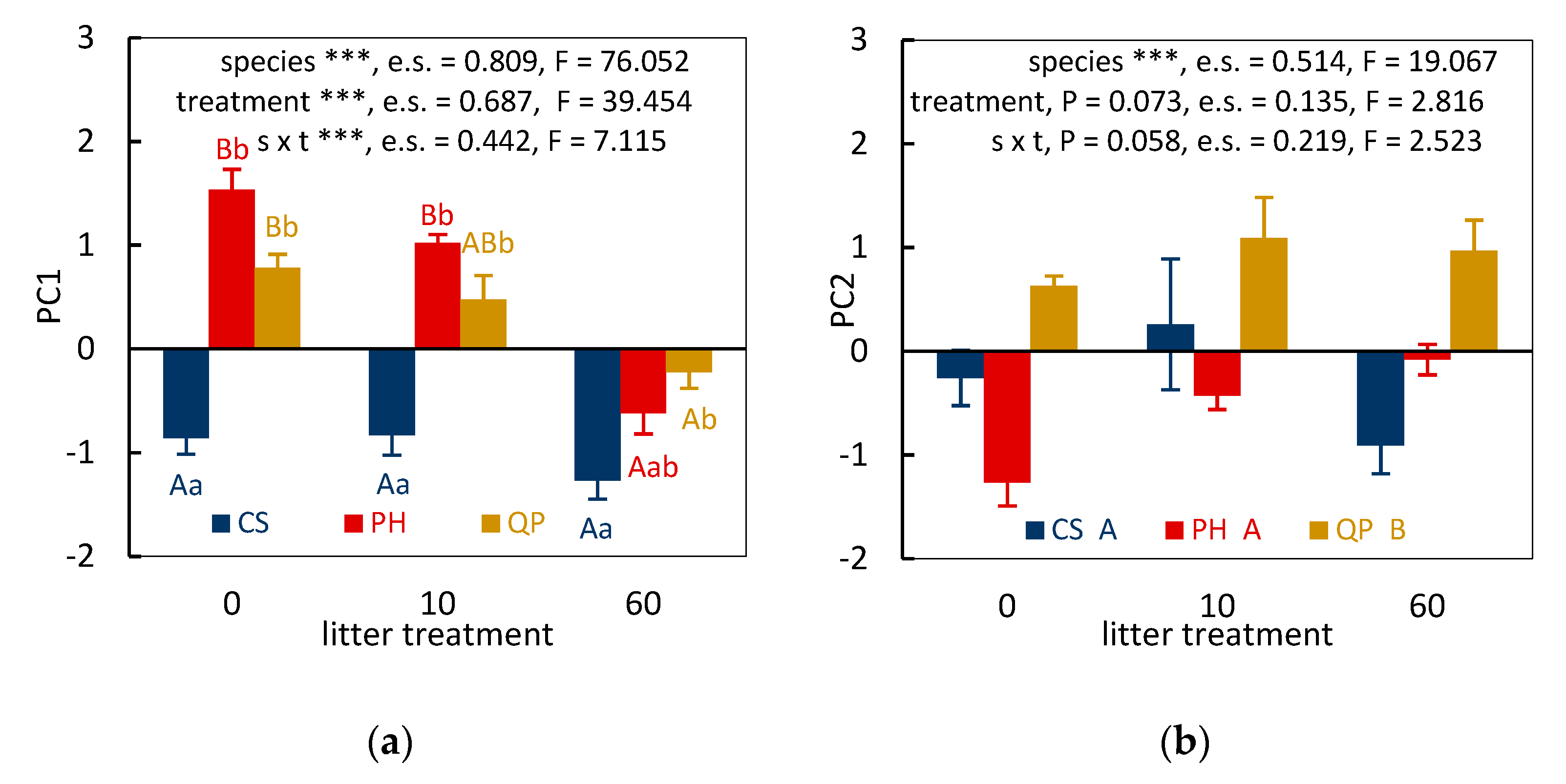
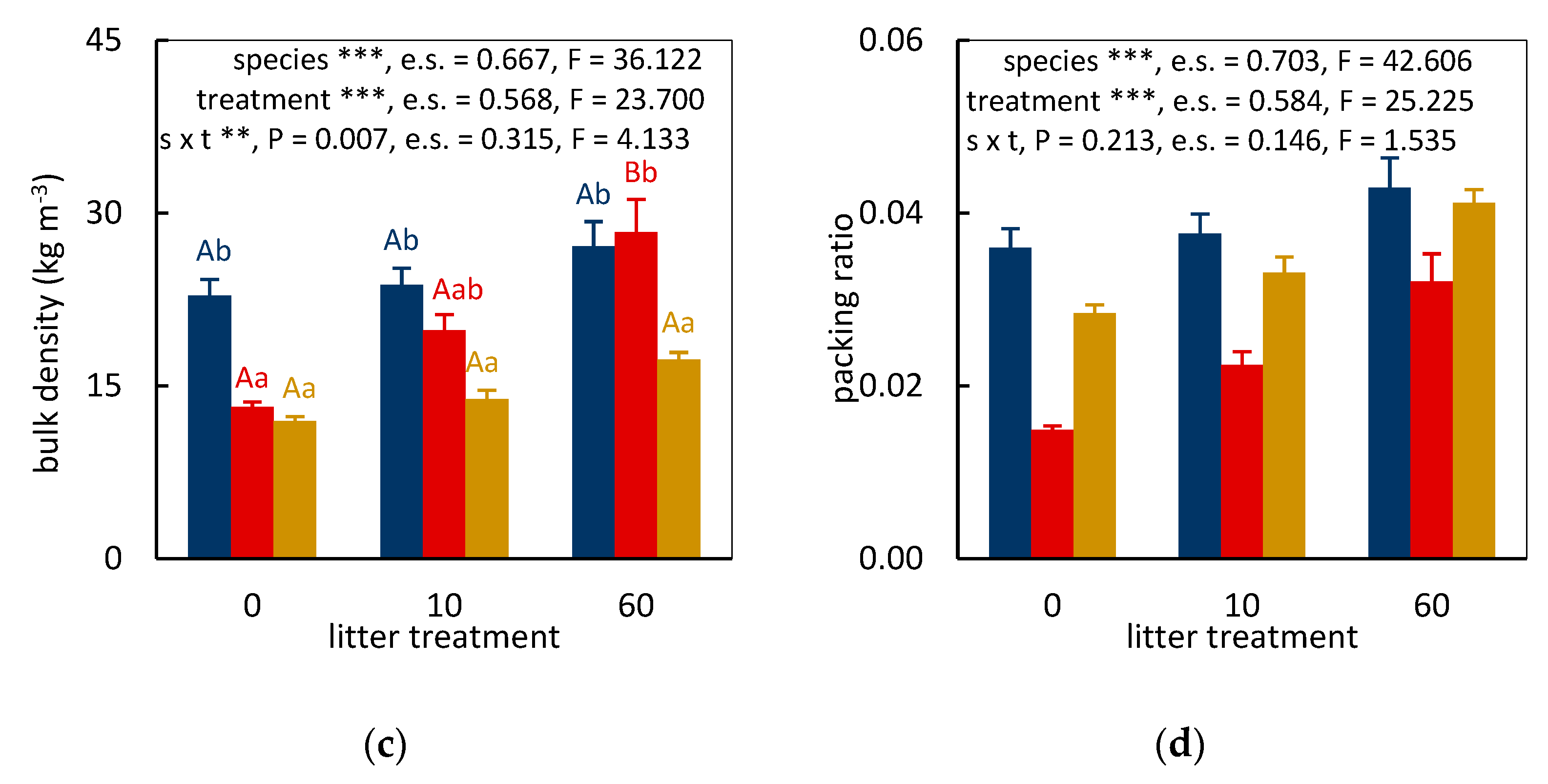
3.2. Significance of Treatment Effects
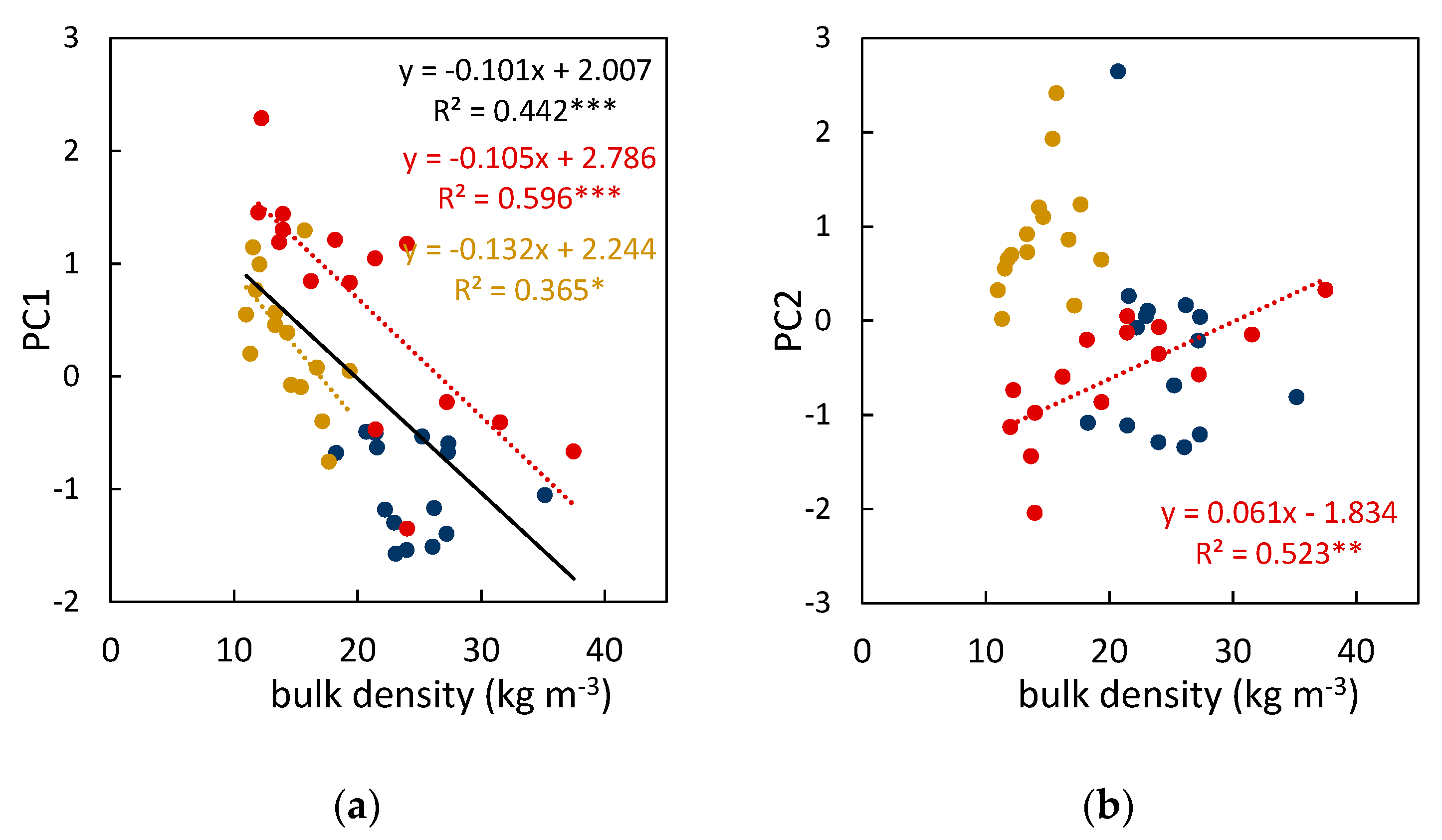
3.3. Fuel Bed Aeration and Fire Behavior of Leaf Litter Beds
4. Discussion
4.1. Current Understanding of the Fire Behavior of the Leaf Litter
4.2. Which Characteristics Control Fuel Bed Compaction?
4.3. Is the Observed Fuel Bed Compaction a Result of Inadequate Fuel Bed Construction?
4.4. Treatment Induced Alterations in the Fire Behavior
4.5. Importance of the Observed Changes
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scott, A.C.; Glasspool, I.J. The diversification of Paleozoic fire systems and fluctuations in atmospheric oxygen concentration. Proc. Natl. Acad. Sci. USA 2006, 103, 10861–10865. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Keeley, J.E.; Schwilk, D.W. Flammability as an ecological and evolutionary driver. J. Ecol. 2017, 105, 289–297. [Google Scholar] [CrossRef]
- Archibald, S.; Lehmann, C.E.R.; Belcher, C.M.; Bond, W.J.; Bradstock, R.A.; Daniau, A.-L.; Dexter, K.G.; Forrestel, E.J.; Greve, M.; He, T.; et al. Biological and geophysical feedbacks with fire in the Earth system. Environ. Res. Lett. 2018, 13, 033003. [Google Scholar] [CrossRef]
- Anderson, H.E. Forest fuel ignitibility. Fire Technol. 1970, 6, 312–319. [Google Scholar] [CrossRef]
- Martin, R.E.; Gordon, D.A.; Gutierrez, M.E.; Lee, D.S.; Molina, D.M.; Schroeder, R.A.; Sapsis, D.B.; Stephens, S.L.; Chambers, M. Assessing the flammability of domestic and wildland vegetation. In Proceedings of the 12th Conference on Fire and Forest Meteorology, Jekyll Island, GA, USA, 26–28 October 1993; Society of American Foresters: Bethesda, MD, USA, 1994; pp. 130–137. [Google Scholar]
- Prior, L.; Murphy, B.; Bowman, D. Conceptualizing Ecological Flammability: An Experimental Test of Three Frameworks Using Various Types and Loads of Surface Fuels. Fire 2018, 1, 14. [Google Scholar] [CrossRef]
- Varner, J.M.; Kane, J.M.; Kreye, J.K.; Engber, E. The Flammability of Forest and Woodland Litter: A Synthesis. Curr. For. Rep. 2015, 1, 91–99. [Google Scholar] [CrossRef]
- Manea, A.; Grootemaat, S.; Leishman, M.R. Leaf flammability and fuel load increase under elevated CO2 levels in a model grassland. Int. J. Wildl. Fire 2015, 24, 819–827. [Google Scholar] [CrossRef]
- Ellair, D.P.; Platt, W.J. Fuel composition influences fire characteristics and understorey hardwoods in pine savanna. J. Ecol. 2013, 101, 192–201. [Google Scholar] [CrossRef]
- Schwilk, D.W.; Caprio, A.C. Scaling from leaf traits to fire behaviour: Community composition predicts fire severity in a temperate forest. J. Ecol. 2011, 99, 970–980. [Google Scholar] [CrossRef]
- Zylstra, P.; Bradstock, R.A.; Bedward, M.; Penman, T.D.; Doherty, M.D.; Weber, R.O.; Gill, A.M.; Cary, G.J. Biophysical Mechanistic Modelling Quantifies the Effects of Plant Traits on Fire Severity: Species, Not Surface Fuel Loads, Determine Flame Dimensions in Eucalypt Forests. PLoS ONE 2016, 11, e0160715. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Elvira, A.; van Kempen, L.; van Logtestijn, R.S.P.P.; Aptroot, A.; Cornelissen, J.H.C. Flammability across the gymnosperm phylogeny: The importance of litter particle size. New Phytol. 2015, 206, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Engber, E.A.; Varner, J.M. Patterns of flammability of the California oaks: The role of leaf traits. Can. J. For. Res. 2012, 42, 1965–1975. [Google Scholar] [CrossRef]
- Simpson, K.J.; Ripley, B.S.; Christin, P.-A.; Belcher, C.M.; Lehmann, C.E.R.R.; Thomas, G.H.; Osborne, C.P. Determinants of flammability in savanna grass species. J. Ecol. 2016, 104, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Ganteaume, A.; Jappiot, M.; Curt, T.; Lampin, C.; Borgniet, L. Flammability of litter sampled according to two different methods: Comparison of results in laboratory experiments. Int. J. Wildl. Fire 2014, 23, 1061. [Google Scholar] [CrossRef]
- Grootemaat, S.; Wright, I.J.; van Bodegom, P.M.; Cornelissen, J.H.C.; Cornwell, W.K. Burn or rot: Leaf traits explain why flammability and decomposability are decoupled across species. Funct. Ecol. 2015, 29, 1486–1497. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Grootemaat, S.; Verheijen, L.M.; Cornwell, W.K.; van Bodegom, P.M.; van der Wal, R.; Aerts, R. Are litter decomposition and fire linked through plant species traits? New Phytol. 2017, 216, 653–669. [Google Scholar] [CrossRef]
- Weir, J.R.; Limb, R.F. Seasonal Variation in Flammability Characteristics of Quercus marilandica and Quercus stellata Leaf Litter Burned in the Laboratory. Fire Ecol. 2013, 9, 80–88. [Google Scholar] [CrossRef]
- Kauf, Z.; Fangmeier, A.; Rosavec, R.; Španjol, Ž. Seasonal and Local Differences in Leaf Litter Flammability of Six Mediterranean Tree Species. Environ. Manag. 2015, 55, 687–701. [Google Scholar] [CrossRef]
- Kauf, Z.; Damsohn, W.; Fangmeier, A. Do relationships between leaf traits and fire behaviour of leaf litter beds persist in time? PLoS ONE 2018, 13, e0209780. [Google Scholar] [CrossRef]
- Ormeño, E.; Céspedes, B.; Sánchez, I.A.; Velasco-García, A.; Moreno, J.M.; Fernandez, C.; Baldy, V. The relationship between terpenes and flammability of leaf litter. For. Ecol. Manag. 2009, 257, 471–482. [Google Scholar] [CrossRef]
- Scarff, F.R.; Westoby, M. Leaf litter flammability in some semi-arid Australian woodlands. Funct. Ecol. 2006, 20, 745–752. [Google Scholar] [CrossRef]
- Grootemaat, S.; Wright, I.J.; van Bodegom, P.M.; Cornelissen, J.H.C. Scaling up flammability from individual leaves to fuel beds. Oikos 2017, 126, 1428–1438. [Google Scholar] [CrossRef]
- De Magalhães, R.M.Q.Q.; Schwilk, D.W. Leaf traits and litter flammability: Evidence for non-additive mixture effects in a temperate forest. J. Ecol. 2012, 100, 1153–1163. [Google Scholar] [CrossRef]
- Prior, L.D.; Murphy, B.P.; Williamson, G.J.; Cochrane, M.A.; Jolly, W.M.; Bowman, D.M.J.S. Does inherent flammability of grass and litter fuels contribute to continental patterns of landscape fire activity? J. Biogeogr. 2017, 44, 1225–1238. [Google Scholar] [CrossRef]
- Van Altena, C.; van Logtestijn, R.S.P.; Cornwell, W.K.; Cornelissen, J.H.C. Species composition and fire: Non-additive mixture effects on ground fuel flammability. Front. Plant Sci. 2012, 3, 63. [Google Scholar] [PubMed]
- Nyman, P.; Metzen, D.; Noske, P.J.; Lane, P.N.J.; Sheridan, G.J. Quantifying the effects of topographic aspect on water content and temperature in fine surface fuel. Int. J. Wildl. Fire 2015, 24, 1129. [Google Scholar] [CrossRef]
- Baum, C.; Fienemann, M.; Glatzel, S.; Gleixner, G. Overstory-specific effects of litter fall on the microbial carbon turnover in a mature deciduous forest. For. Ecol. Manag. 2009, 258, 109–114. [Google Scholar] [CrossRef]
- Berger, T.W.; Berger, P. Greater accumulation of litter in spruce (Picea abies) compared to beech (Fagus sylvatica) stands is not a consequence of the inherent recalcitrance of needles. Plant Soil 2012, 358, 349–369. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, J.H.C. An experimental comparison of leaf decomposition rates in a wide range of temperate plant species and types. J. Ecol. 1996, 84, 573–582. [Google Scholar] [CrossRef]
- Berg, B. Decomposition patterns for foliar litter—A theory for influencing factors. Soil Biol. Biochem. 2014, 78, 222–232. [Google Scholar] [CrossRef]
- Kreye, J.K.; Varner, J.M.; Hiers, J.K.; Mola, J. Toward a mechanism for eastern North American forest mesophication: Differential litter drying across 17 species. Ecol. Appl. 2013, 23, 1976–1986. [Google Scholar] [CrossRef] [PubMed]
- Anderson, H.E. Predicting Equilibrium Moisture Content of Some Foliar Forest Litter in the Northern Rocky Mountains; Research Paper INT-429; U.S. Department of Agriculture, Forest Service, Intermountain Research Station (USA): Ogden, UT, USA, 1990. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Rothermel, R.C. A Mathematical Model for Predicting Fire Spread in Wildland Fuels; U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1972. [Google Scholar]
- Cohen, J. A power primer. Psychol. Bull. 1992, 112, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, P.M.; Cruz, M.G. Plant flammability experiments offer limited insight into vegetation-fire dynamics interactions. New Phytol. 2012, 194, 606–609. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Moreira, B. Flammability as a biological concept. New Phytol. 2012, 194, 610–613. [Google Scholar] [CrossRef] [PubMed]
- Parsons, A.L.; Balch, J.K.; de Andrade, R.B.; Brando, P.M. The role of leaf traits in determining litter flammability of south-eastern Amazon tree species. Int. J. Wildl. Fire 2015, 24, 1143. [Google Scholar] [CrossRef]
- Dickinson, M.B.; Hutchinson, T.F.; Dietenberger, M.; Matt, F.; Peters, M.P. Litter Species Composition and Topographic Effects on Fuels and Modeled Fire Behavior in an Oak-Hickory Forest in the Eastern USA. PLoS ONE 2016, 11, e0159997. [Google Scholar] [CrossRef] [PubMed]
- Ottmar, R.; Andreu, A. Litter and Duff Bulk Densities in the Southern United States Joint Fire Science Program Project #04-2-1-49 Final Report; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Seattle, WA, USA, 2007. [Google Scholar]
- Mutch, R.W. Wildland Fires and Ecosystems—A Hypothesis. Ecology 1970, 51, 1046–1051. [Google Scholar] [CrossRef]
- Mendes-Lopes, J.M.C.; Ventura, J.M.P.; Amaral, J.M.P. Flame characteristics, temperature-time curves, and rate of spread in fires propagating in a bed of Pinus pinaster needles. Int. J. Wildl. Fire 2003, 12, 67–84. [Google Scholar] [CrossRef]
- Madrigal, J.; Guijarro, M.; Hernando, C.; Díez, C.; Marino, E. Estimation of Peak Heat Release Rate of a Forest Fuel Bed in Outdoor Laboratory Conditions. J. Fire Sci. 2011, 29, 53–70. [Google Scholar] [CrossRef]
- Curt, T.; Schaffhauser, A.; Borgniet, L.; Dumas, C.; Estève, R.; Ganteaume, A.; Jappiot, M.; Martin, W.; N’Diaye, A.; Poilvet, B. Litter flammability in oak woodlands and shrublands of southeastern France. For. Ecol. Manag. 2011, 261, 2214–2222. [Google Scholar] [CrossRef]
- Sikkink, P.G.; Jain, T.B.; Reardon, J.; Heinsch, F.A.; Keane, R.E.; Butler, B.; Baggett, L.S. Effect of particle aging on chemical characteristics, smoldering, and fire behavior in mixed-conifer masticated fuel. For. Ecol. Manag. 2017, 405, 150–165. [Google Scholar] [CrossRef]
- Bueis, T.; Bravo, F.; Pando, V.; Turrión, M.B. Local basal area affects needle litterfall, nutrient concentration, and nutrient release during decomposition in Pinus halepensis Mill. plantations in Spain. Ann. For. Sci. 2018, 75, 21. [Google Scholar] [CrossRef]
- Schunk, C.; Leutner, C.; Leuchner, M.; Wastl, C.; Menzel, A. Equilibrium moisture content of dead fine fuels of selected central European tree species. Int. J. Wildl. Fire 2013, 22, 797–809. [Google Scholar] [CrossRef]
- Fonda, R.W. Burning characteristics of needles from eight pine species. For. Sci. 2001, 47, 390–396. [Google Scholar]
- Mola, J.M.; Varner, J.M.; Jules, E.S.; Spector, T. Altered Community Flammability in Florida’s Apalachicola Ravines and Implications for the Persistence of the Endangered Conifer Torreya taxifolia. PLoS ONE 2014, 9, e103933. [Google Scholar] [CrossRef] [PubMed]
- Kane, J.M.; Varner, J.M.; Hiers, J.K. The burning characteristics of southeastern oaks: Discriminating fire facilitators from fire impeders. For. Ecol. Manag. 2008, 256, 2039–2045. [Google Scholar] [CrossRef]
- Olson, J.S. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Alexander, H.D.; Arthur, M.A. Increasing Red Maple Leaf Litter Alters Decomposition Rates and Nitrogen Cycling in Historically Oak-Dominated Forests of the Eastern U.S. Ecosystems 2014, 17, 1371–1383. [Google Scholar] [CrossRef]
- García-Plé, C.; Vanrell, P.; Morey, M. Litter fall and decomposition in a Pinus halepensis forest on Mallorca. J. Veg. Sci. 1995, 6, 17–22. [Google Scholar] [CrossRef]
- Engber, E.A.; Varner, J.M., III; Arguello, L.A.; Sugihara, N.G. The Effects of Conifer Encroachment and Overstory Structure on Fuels and Fire in an Oak Woodland Landscape. Fire Ecol. 2011, 7, 32–50. [Google Scholar] [CrossRef]
- Vander Yacht, A.L.; Keyser, P.D.; Barrioz, S.A.; Kwit, C.; Stambaugh, M.C.; Clatterbuck, W.K.; Simon, D.M. Reversing Mesophication Effects on Understory Woody Vegetation in Mid-Southern Oak Forests. For. Sci. 2019, 65, 289–303. [Google Scholar] [CrossRef]
- Sparks, J.C.; Masters, R.E.; Engle, D.M.; Bukenhofer, G.A. Season of Burn Influences Fire Behavior and Fuel Consumption in Restored Shortleaf Pine-Grassland Communities. Restor. Ecol. 2002, 10, 714–722. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kauf, Z.; Damsohn, W.; Fangmeier, A. How Much Does Fire Behavior of Leaf Litter Beds Change within Two Months? Fire 2019, 2, 33. https://doi.org/10.3390/fire2020033
Kauf Z, Damsohn W, Fangmeier A. How Much Does Fire Behavior of Leaf Litter Beds Change within Two Months? Fire. 2019; 2(2):33. https://doi.org/10.3390/fire2020033
Chicago/Turabian StyleKauf, Zorica, Walter Damsohn, and Andreas Fangmeier. 2019. "How Much Does Fire Behavior of Leaf Litter Beds Change within Two Months?" Fire 2, no. 2: 33. https://doi.org/10.3390/fire2020033
APA StyleKauf, Z., Damsohn, W., & Fangmeier, A. (2019). How Much Does Fire Behavior of Leaf Litter Beds Change within Two Months? Fire, 2(2), 33. https://doi.org/10.3390/fire2020033