Flexibility and Hydration of Amphiphilic Hyperbranched Arabinogalactan-Protein from Plant Exudate: A Volumetric Perspective

 , ,
, ,
Abstract

1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Chemical Analyses
2.2.2. Desalting of Acacia Gums and HIC Fractions
2.2.3. Preparation of Acacia Gum Dispersions
2.2.4. Size Exclusion Chromatography (HPSEC)-Multi Angle Light Scattering (MALS)
2.2.5. Density and Sound Velocity Measurements
2.3. Theoretical Treatment of Density and Sound Velocity Parameters
2.3.1. Partial Specific Volume
2.3.2. Isoentropic Compressibility Coefficients
2.3.3. Microscopic Description of Macroscopic Volumetric Data
3. Results
3.1. Theoretical Treatment of Density and Sound Velocity Parameters
3.2. Volumetric Properties
4. Discussion
4.1. Microscopic Description of AGP Volumetric Experimental Data
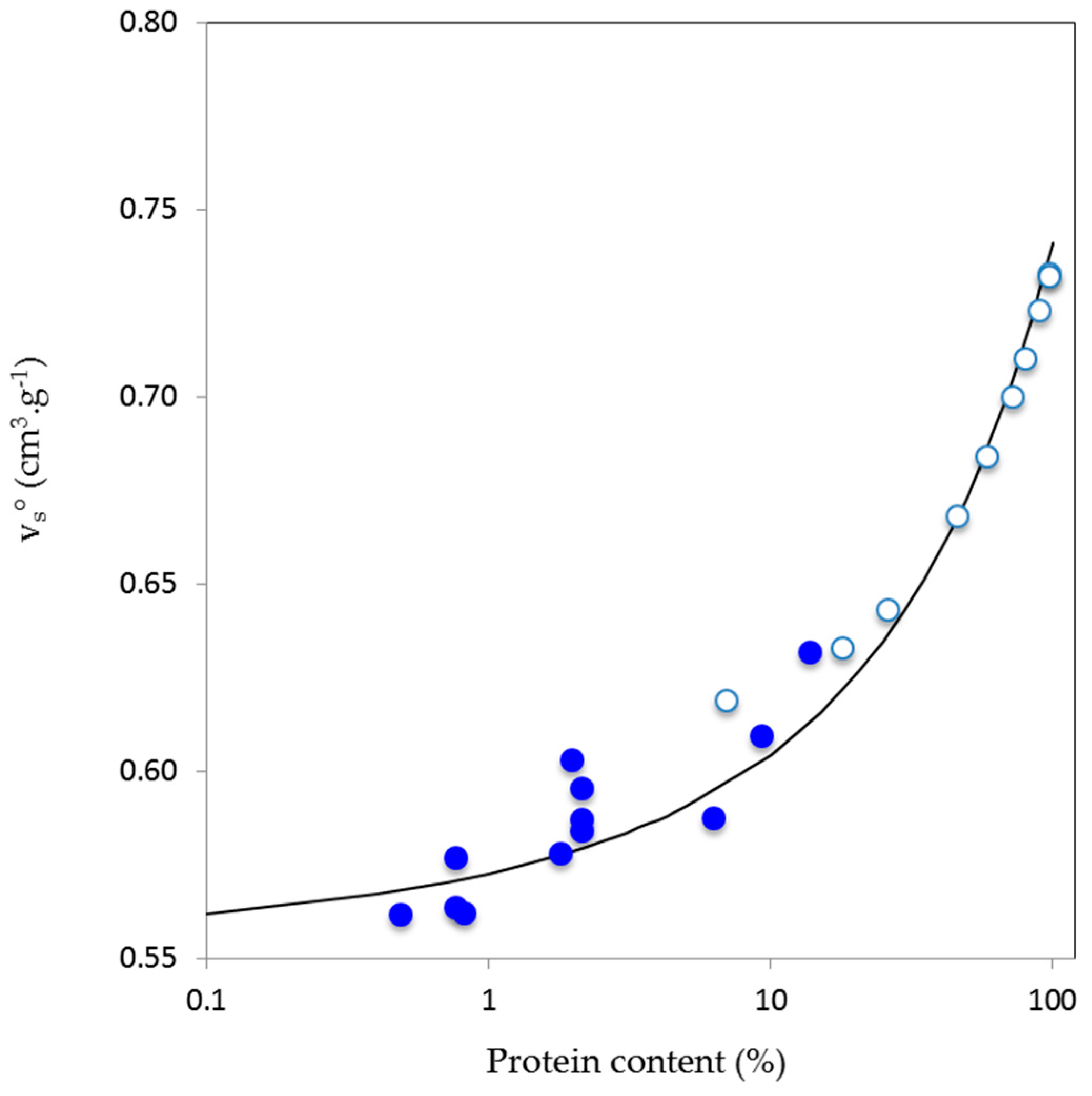
4.2. Partial Molar Volumes of AGPs
4.3. Partial Molar Adiabatic Compressibility of AGPs
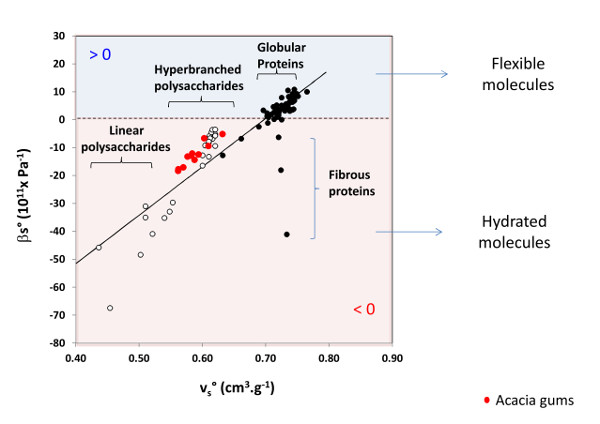
4.4. Additional Comments on the Hydration Properties of AGPs
5. Concluding Remarks
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Williams, P.A.; Philips, G.O. Gum arabic. In Handbook of Hydrocolloids; CRC Press: Boca Raton, MA, USA, 2000; pp. 155–168. [Google Scholar]
- Nussinovitch, A. Plant Gum Exudates of the World: Sources, Distribution, Properties and Aplication; CRC Press: Boca Raton, MA, USA, 2010. [Google Scholar]
- Verbeken, D.; Dierckx, S.; Dewettinck, K. Exudate gums: Occurence, production, and applications. Appl. Microbiol. Biot. 2003, 63, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.; Nigen, M.; Mejia Tamayo, V.; Doco, T.; Williams, P.; Amine, C.; Renard, D. Acacia gum: History of the future. Food Hydrocoll. 2017. [Google Scholar] [CrossRef]
- Akiyama, Y.; Eda, S.; Kato, K. Gum arabic is a kind of arabinogalactan protein. Agric. Biol. Chem. 1984, 48, 235–237. [Google Scholar]
- Tan, L.; Showalter, A.M.; Egelund, J.; Hernandez-Sanchez, A.; Doblin, M.S.; Bacic, A. Arabinogalactan-proteins and the research challenges for these enigmatic plant cell surface proteoglycans. Fronti. Plant Sci. 2012, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lamport, D.T.A.; Várnai, P. Periplasmic arabinogalactan glycoproteins act as a calcium capacitor that regulates plant growth and development. New Phytol. 2013, 197, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.G.; Mehdy, M.C. A nonclassical arabinogalactan protein gene highly expressed in vascular tissues agp31, is transcrptionally repressed by methyl jasmonic acid in arabidopsis. Plant Physiol. 2007, 145, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.; Egelund, J.; Schultz, C.J.; Bacic, A. Arabinogalactan-proteins: Key regulators at the cell surface? Plant Physiol. 2010, 153, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Showalter, A.M. Arabinogalactan-proteins: Structure, expression and function. Cell. Mol. Life Sci. 2001, 58, 1399–1417. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.M.; Egelund, J.; Gilson, P.R.; Houghton, F.; Gleeson, P.A.; Schultz, C.J.; Bacic, A. Identification of a novel group of putative arabidopsis thaliana beta-(1,3)-galactosyltransferases. Plant Mol. Biol. 2008, 68, 43–59. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.W.; Stoddart, J.F. Studies on uronic acid materials. Part xv. The use of molecular-sieve chromatography on acacia senegal gum (gum arabic). Carbohydr. Res. 1966, 2, 104–114. [Google Scholar] [CrossRef]
- Randall, R.C.; Phillips, G.O.; Williams, P.A. Fractionation and characterization of gum from acacia senegal. Food Hydrocoll. 1989, 3, 65–75. [Google Scholar] [CrossRef]
- Islam, A.M.; Phillips, G.O.; Sljivo, A.; Snowden, M.J.; Williams, P.A. A review of recent developments on the regulatory, structural and functional aspects of gum arabic. Food Hydrocoll. 1997, 11, 493–505. [Google Scholar] [CrossRef]
- Renard, D.; Lavenant-Gourgeon, L.; Ralet, M.C.; Sanchez, C. Acacia senegal gum: Continuum of molecular species differing by their protein to sugar ratio; molecular weight; and charges. Biomacromolecules 2006, 2637–2649. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, T.; Williams, P.; Phillips, G.; Al-Assaf, S.; Baldwin, T. New insights into the structural characteristics of the arabinogalactan-protein (agp) fraction of gum arabic. J. Agric. Food Chem. 2008, 56, 9269–9276. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.; Schmitt, C.; Kolodziejczyk, E.; Lapp, A.; Gaillard, C.; Renard, D. The acacia gum arabinogalactan fraction is a thin oblate ellipsoid: A new model based on small-angle neutron scattering and ab initio calculation. Biophys. J. 2008, 94, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Renard, D.; Garnier, C.; Lapp, A.; Schmitt, C.; Sanchez, C. Structure of arabinogalactan-protein from acacia gum: From porous ellipsoids to supramolecular architectures. Carbohydr. Polym. 2012, 90, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Renard, D.; Lavenant-Gourgeon, L.; Lapp, A.; Nigen, M.; Sanchez, C. Enzymatic hydrolysis studies of arabinogalactan-protein structure from acacia gum: The self-similarity hypothesis of assembly from a common builiding block. Carbohydr. Polym. 2014, 112, 648–661. [Google Scholar] [CrossRef] [PubMed]
- Renard, D.; Lepvrier, E.; Garnier, C.; Roblin, P.; Nigen, M.; Sanchez, C. Structure of glycoproteins from acacia gum: An assembly of ring-like glycoproteins modules. Carbohydr. Polym. 2014, 99, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Torrez, L.; Nigen, M.; Williams, P.; Doco, T.; Sanchez, C. Acacia senegal vs. Acacia seyal gums—part 1: Composition and structure of hyperbranched plant exudates. Food Hydrocoll. 2015, 51, 41–53. [Google Scholar] [CrossRef]
- Renard, D.; Garnier, C.; Lapp, A.; Schmitt, C.; Sanchez, C. Corrigendum to “structure of arabinogalactan-protein from acacia gum: From porous ellipsoids to supramolecular architectures” [carbohydr. Polym. 90 (2012) 322–332]. Carbohydr. Polym. 2013, 97, 864–867. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.K.; Bird, P.B.; Iacobucci, G.A.; Clark, B.C. Functionality of gum arabic. Fractionation, characterization and evaluation of gum fractions in citrus oil emulsions and model beverages. Food Hydrocoll. 1995, 9, 123–131. [Google Scholar] [CrossRef]
- Idris, O.H.M.; Williams, P.A.; Phillips, G.O. Characterisation of gum from acacia senegal trees of different age and location using multidetection gel permation chromatography. Food Hydrocoll. 1998, 379–388. [Google Scholar] [CrossRef]
- Dror, Y.; Cohen, Y.; Yerushalmi-Rozen, R. Structure of gum arabic in aqueous solution. J. Polym. Sci. Pol. Phys. 2006, 44, 3265–3271. [Google Scholar] [CrossRef]
- Wang, Q.; Burchard, W.; Cui, S.W.; Huang, X.Q.; Philips, G.O. Solution properties of conventional gum arabic and a matured gum arabic (acacia (sen) super gum). Biomacromolecules 2008, 9, 1163–1169. [Google Scholar] [CrossRef] [PubMed]
- Al-Assaf, S.; Sakata, M.; McKenna, C.; Aoki, H.; Phillips, G.O. Molecular associations in acacia gums. Struct. Chem. 2009, 20, 325–336. [Google Scholar] [CrossRef]
- Kharakoz, D.P. Protein compressibility, dynamics, and pressure. Biophys. J. 2000, 79, 511–525. [Google Scholar] [CrossRef]
- Chalikian, T.V.; Totrov, M.; Abagyan, R.; Breslauer, K.J. The hydration of globular proteins as derived from volume and compressibility measurements: Cross correlating thermodynamic and structural data. J. Mol. Biol. 1996, 260, 588–603. [Google Scholar] [CrossRef] [PubMed]
- Scharnagl, R.M.; Friedrich, J. Local compressibilities of compressibilities: Comparison of optical experiments and simulations for horse heart cytochrome-c. Biophys. J. 2005, 89, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Gavish, B.; Gratton, E.; Hardy, C.J. Adiabatic compressibility of globular proteins. Proc. Natl. Acad. Sci. USA 1983, 80, 750–754. [Google Scholar] [CrossRef] [PubMed]
- Gekko, K.; Hasegawa, Y. Compressibility-structure relationship of globular proteins. Biochemistry 1986, 25, 6563–6571. [Google Scholar] [CrossRef] [PubMed]
- Durchschlag, H. Specific volumes of biological macromolecules and some other molecules of biological interest. In Thermodynamic Data for Biochemistry and Biotechnology; Hinz, H.-J., Ed.; Springer: Berlin, Germany, 1986; pp. 45–128. [Google Scholar]
- Gekko, K. Hydration-structure-function relationships of polysaccharides and proteins. Food Hydrocoll. 1989, 3, 289–299. [Google Scholar] [CrossRef]
- Sarvazyan, A.P. Ultrasonic velocimetry of biological compounds. Annu. Rev. Biophys. Biophys. Chem. 1991, 20, 321–342. [Google Scholar] [CrossRef] [PubMed]
- Kharakoz, D.P.; Sarvazyan, A.P. Hydrational and intrinsic compressibilities of globular proteins. Biopolymers 1993, 33, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Chalikian, T.V.; Sarvazyan, A.P.; Breslauer, K.J. Hydration and partial compressibility of biological compounds. Biophys. Chem. 1994, 51, 89–109. [Google Scholar] [CrossRef]
- Chalikian, T.V.; Filfil, R. How large are the volume changes accompanying protein transitions and binding? Biophys. Chem. 2003, 104, 489–499. [Google Scholar] [CrossRef]
- Dadarlat, V.M.; Post, C.B. Decomposition of protein experimental compressibility into intrinsic and hydration shell contributions. Biophys. J. 2006, 91, 4544–4554. [Google Scholar] [CrossRef] [PubMed]
- Gekko, K. Volume and compressibility of proteins. In High Pressure Bioscience: Basic Concepts, Applications and Frontiers; Akasaka, K., Matsuki, H., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 75–108. [Google Scholar]
- Buckin, V.A.; Kankiya, B.I.; Sarvazyan, A.P.; Uedaira, H. Acoustical investigation of poly(da)-poly(dt), poly[d(a-t)]-poly[d(at)], poly (a).Polyflj) and DNA hydration in dilute aqueous solutions. Nucleic Acids Res. 1989, 17, 4189–4203. [Google Scholar] [CrossRef] [PubMed]
- Chalikian, T.V.; Sarvazyan, A.P.; Plum, S.G.E.; Breslauer, K.J. Influence of base composition, base sequence, and duplex structure on DNA hydration: Apparent molar volumes and apparent molar adiabatic compressibilities of synthetic and natural DNA duplexes at 25 °C. Biochemistry 1994, 33, 2394–2401. [Google Scholar] [CrossRef] [PubMed]
- Chalikian, T.V.; McGregor, J.R.B. Nucleic acid hydration: A volumetric perspective. Phys. Life Rev. 2007, 4, 91–115. [Google Scholar] [CrossRef]
- Myahara, Y.; Shio, H. The adiabatic compressibility of starch sols. Nippon Kagaku Zasshi 1952, 73, 1–2. [Google Scholar] [CrossRef][Green Version]
- Shiio, H.; Yoshihashi, H. Measurement of the amount of bound water by ultrasonic interferometer. Ii. Polyvinyl alcohol and its partially substituted acetates. J. Phys. Chem. 1956, 60, 1049–1051. [Google Scholar] [CrossRef]
- Itoh, K. Adiabatic compressibility of polysaccharides. Nippon Kagaku Zasshi 1956, 77, 1594–1595. [Google Scholar] [CrossRef]
- Nomura, H.; Miyahara, Y. Partial specific compressibility of polystyrene. J. Appl. Polym. Sci. 1964, 8, 1643–1649. [Google Scholar] [CrossRef]
- Suzuki, Y.; Uedaira, H. Hydration of potassium hyaluronate. Bull. Chem. Soc. Jpn. 1970, 43, 1892–1894. [Google Scholar] [CrossRef]
- Gekko, K.; Noguchi, H. Physicochemical studies of oligodextran. I. Molecular weight dependence of intrinsic viscosity, partial specific compressibility and hydrated water. Biopolymers 1971, 10, 1513–1524. [Google Scholar] [CrossRef] [PubMed]
- Gekko, K.; Noguchi, H. Hydration behavior of ionic dextran derivatives. Macromolecules 1974, 7, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Kawaizumi, F.; Nishio, N.; Nomura, H.; Miyahara, Y. Calorimetric and compressibility study of aqueous solutions of dextran with special reference to hydration and structural change of water. Polym. J. 1981, 13, 209–213. [Google Scholar] [CrossRef]
- Nomura, H.; Onoda, M.; Miyahara, Y. Preferential solvation of dextran in water—ethanol mixtures. Polym. J. 1982, 14, 249–253. [Google Scholar] [CrossRef]
- Davies, A.; Gormally, J.; Wyn-Jones, E.; Wedlock, D.J.; Phillips, G.O. A study of factors influencing hydration of sodium hyaluronate from compressibility and high precision densimetric measurements. Biochem. J. 1983, 213, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Gekko, K.; Mugishima, H.; Koga, S. Compressibility, densimetric and calorimetric studies of hydration of carrageenans in the random form. Int. J. Biol. Macromol. 1985, 7, 57–63. [Google Scholar] [CrossRef]
- Kupke, D.W. Density and volume change measurements. In Physical Principles and Techniques of Protein Chemistry, Part C; Leach, S.J., Ed.; Academic Press: Cambridge, MA, USA, 1973; pp. 3–75. [Google Scholar]
- Hoiland, H. Partial molar volumes of biochemical model compounds in aqueous solution. In Thermodynamic Data for Biochemistry and Biotechnology; Hinz, H.-J., Ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 1986; pp. 17–44. [Google Scholar]
- Gekko, K.; Yamagami, K. Flexibility of food proteins as revealed by compressibility. J. Agric. Food Chem. 1991, 39, 57–62. [Google Scholar] [CrossRef]
- Chalikian, T.V.; Sarvazyan, A.P.; Breslauer, K.J. Partial molar volumes, expansibilities, and compressibilities of.Alpha.,.Omega.-aminocarboxylic acids in aqueous solutions between 18 and 55 °C. J. Phys. Chem. 1993, 97, 13017–13026. [Google Scholar] [CrossRef]
- Taulier, N.; Chalikian, T.V. Compressibility of protein transitions. BBA-Protein Struct. Mol. 2002, 1595, 48–70. [Google Scholar] [CrossRef]
- Gekko, K.; Noguchi, H. Compressibility of globular proteins in water at 25 °C. J. Phys. Chem. 1979, 83, 2706–2714. [Google Scholar] [CrossRef]
- Pierotti, R.A. A scaled particle theory of aqueous non aqueous solutions. Chem. Rev. 1976, 76, 717–726. [Google Scholar] [CrossRef]
- Reiss, H. Scaled particle methodsin the statistical thermodynamics of fluids. Adv. Chem. Phys. 1965, 9, 1–84. [Google Scholar]
- Kharakoz, D.P. Volumetric properties of proteins and their analogs in diluted water solutions. Biophys. Chem. 1989, 34, 115–125. [Google Scholar] [CrossRef]
- Kharakoz, D.P. Partial volumes of molecules of arbitrary shape and the effect of hydrogen bonding with water. J. Solut. Chem. 1992, 21, 569–595. [Google Scholar] [CrossRef]
- Pierotti, R.A. The solubility of gases in liquids. J. Phys. Chem. 1963, 67, 1840–1845. [Google Scholar] [CrossRef]
- Kauzmann, W. Some factors in the interpretation of protein denaturation. Adv. Protein Chem. 1959, 14, 1–63. [Google Scholar] [PubMed]
- Bánó, M.; Marek, J. How thick is the layer of thermal volume surrounding the protein? Biophys. Chem. 2006, 120, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Edward, J.T.; Farrell, P.G. Relation between van der waals and partial molal volumes of organic molecules in water. Can. J. Chem. 1975, 53, 2965–2970. [Google Scholar] [CrossRef]
- Voloshin, V.P.; Medvedev, N.N.; Smolin, N.; Geiger, A.; Winter, R. Exploring volume, compressibility and hydration changes of folded proteins upon compression. Phys. Chem. Chem. Phys. 2015, 17, 8499–8508. [Google Scholar] [CrossRef] [PubMed]
- Chalikian, T.V. Structural thermodynamics of hydration. J. Phys. Chem. B 2001, 105, 12566–12578. [Google Scholar] [CrossRef]
- Kharakoz, D.P. Volumetric properties of proteins and their analogues in diluted water solutions. 2. Partial adiabatic compressibilities of amino acids at 15–70.Degree.C. J. Phys. Chem. 1991, 95, 5634–5642. [Google Scholar] [CrossRef]
- Anderson, D.M.W.; Bridgeman, M.M.E.; Farquhar, J.G.K.; McNab, C.G.A. The chemical characterization of the test article used in toxicological studies of gum arabic (acacia senegal (l.) willd). Int. Tree Crops J. 1983, 2, 245–254. [Google Scholar] [CrossRef]
- Flindt, C.; Alassaf, S.; Phillips, G.; Williams, P. Studies on acacia exudate gums. Part v. Structural features of acacia seyal. Food Hydrocoll. 2005, 19, 687–701. [Google Scholar] [CrossRef]
- Hölter, D.; Burgath, A.; Frey, H. Degree of branching in hyperbranched polymers. Acta Polym. 1997, 48, 30–35. [Google Scholar] [CrossRef]
- Gashua, I.B.; Williams, P.A.; Yadav, M.P.; Baldwin, T.C. Characterisation and molecular association of nigerian and sudanese acacia gum exudates. Food Hydrocoll. 2015, 51, 405–413. [Google Scholar] [CrossRef]
- Osman, M.E.; Menzies, A.R.; Williams, P.A.; Phillips, G.O.; Baldwin, T.C. The molecular characterisation of the polysaccharide gum from acacia senegal. Carbohydr. Res. 1993, 246, 303–318. [Google Scholar] [CrossRef]
- Chen, C.-G.; Pu, Z.-Y.; Moritz, R.L.; Simpson, R.J.; Bacic, A.; Clarke, A.E.; Mau, S.-L. Molecular cloning of a gene encoding an arabinogalactan-protein from pear (pyrus communis) cell suspension culture. Proc. Natl. Aca. Sci. USA 1994, 91, 10305–10309. [Google Scholar] [CrossRef]
- Goodrum, L.J.; Patel, A.; Leykam, J.F.; Kieliszewski, M.J. Gum arabic glycoprotein contains glycomodules of both extensin and arabinogalactan-glycoproteins. Phytochemistry 2000, 54, 99–106. [Google Scholar] [CrossRef]
- Al-Assaf, S.; Phillips, G.; Williams, P. Studies on acacia exudate gums: Part ii. Molecular weight comparison of the vulgares and gummiferae series of acacia gums. Food Hydrocoll. 2005, 19, 661–667. [Google Scholar] [CrossRef]
- Sanchez, C.; Renard, D.; Robert, P.; Schmitt, C.; Lefebvre, J. Structure and rheological properties of acacia gum dispersions. Food Hydrocoll. 2002, 16, 257–267. [Google Scholar] [CrossRef]
- Zhu, C.; Gao, Y.; Li, H.; Meng, S.; Li, L.; Francisco, J.S.; Cheng Zeng, X. Characterizing hydrophobicity of amino acid side chains in a protein environment via measuring contact angle of a water nonodroplet on planar peptide network. PNAS 2016, 113, 12946–12951. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Farrell, P.G.; Edward, J.T. Partial molar volumes of organic compounds in water. Iii. Carbohydrates. J. Solut. Chem. 1976, 5, 807–816. [Google Scholar] [CrossRef]
- Banipal, P.K.; Banipal, T.S.; Lark, B.S.; Ahluwalia, J.C. Partial molar heat capacities and volumes of some mono-, di- and tri-saccharides in water at 298.15, 308.15 and 318.15 k. J. Chem. Soc. Faraday Trans. 1997, 93, 81–87. [Google Scholar] [CrossRef]
- Perkins, S.J.; Miller, A.; Hardingham, T.E.; Muir, H. Physical properties of the hyaluronate binding region of proteoglycan from pig laryngeal cartilage: Densitometric and small-angle neutron scattering studies of carbohydrates and carbohydrate-protein macromolecules. J. Mol. Biol. 1981, 150, 69–95. [Google Scholar] [CrossRef]
- Durchschlag, H. Determination of the partial specific volume of conjugated proteins. Colloid Polym. Sci. 1989, 267, 1139–1150. [Google Scholar] [CrossRef]
- Tamura, Y.; Gekko, K.; Yoshioka, K.; Vonderviszt, F.; Namba, K. Adiabatic compressibility of flagellin and flagelar filament of salmonella typhimurium. Biochim. Biophys. Acta 1997, 1335, 120–126. [Google Scholar] [CrossRef]
- Chalikian, T.V.; Völker, J.; Srinivasan, A.R.; Olson, W.; Breslauer, K.J. The hydration of nucleic acids duplexes as assessed by a combination of volumetric and structural techniques. Biopolymers 1999, 50, 459–471. [Google Scholar] [CrossRef]
- Vandevelde, M.C.; Fenyo, J.C. Estimation of the charge density of arabic acid by potentiometry and dye binding. Polym. Bull. 1987, 18, 47–51. [Google Scholar] [CrossRef]
- Chalikian, T.V. On the molecular origins of volumetric data. J. Phys. Chem. B 2008, 112, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Dubins, D.N.; Pomes, R.; Chalikian, T.V. Size dependence of cavity volume: A molecular dynamics study. Biophys. Chem. 2012, 161, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.R.; Makhatadze, G.I. Protein volume: Calculating molecular van der waals and void volume in proteins. BMC Bioinf. 2015, 16, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Bondi, A. Van der waals volumes and radii. J. Phys. Chem. 1964, 68, 441–451. [Google Scholar] [CrossRef]
- Spillane, W.J.; Birch, G.G.; Drew, M.G.B.; Bartolo, I. Correlation of computed van der waals and molecular volumeswith apparent molar volumes (amw) for amino acid, carbohydrate and sulfamate tastant molecules. Relationship between corey-pauling-koltun volumes (vcpk) and computed volumes. Chem. Soc. Perkin Trans. 1992, 2, 497–523. [Google Scholar] [CrossRef]
- Batsanov, S.S. Van der waals radii of elements. Inorg. Mater. 2001, 37, 871–885. [Google Scholar] [CrossRef]
- Carl, W. A monte carlo study of model dendrimers. J. Chem. Soc. Faraday Trans. 1996, 92, 4151–4154. [Google Scholar] [CrossRef]
- Maiti, P.K.; Cagin, T.; Lin, S.-T.; Williamm, A.G. Effect of solvent and ph on the structure of pamam dendrimers. Macromolecules 2005, 38, 979–991. [Google Scholar] [CrossRef]
- Li, T.; Honk, K.; Porcar, L.; Verduzco, R.; Butler, P.D.; Smith, G.S.; Liu, Y.; Wei-Ren Chen, W.R. Assess the intramolecular cavity of a pamam dendrimer in aqueous solution by small-angle neutron scattering. Macromolecules 2008, 41, 8916–8920. [Google Scholar] [CrossRef]
- Maiti, P.K. Pamam dendrimer: A ph controlled nanosponge. Can. J. Chem. 2017, 95, 991–998. [Google Scholar] [CrossRef]
- Kaliannan, P.; Gromiha, M.M.; Elanthiraiyan, M. Solvent accesibility studies on polysaccharides. Int. J. Biol. Macromol. 2001, 28, 135–141. [Google Scholar] [CrossRef]
- Kaliannan, P.; Gromiha, M.M.; Ramamurthi, K.; Elanthiraiyan, M. Solvent accesibility studies on glycosaminoglycans. Biophys. Chem. 1998, 74, 135–141. [Google Scholar] [CrossRef]
- Miller, S.; Lesk, A.M.; Janin, J.; Chothia, C. The accesible surface area and stability of oligomeric proteins. Nature 1987, 328, 834–836. [Google Scholar] [CrossRef] [PubMed]
- Janin, J.; Miller, S.; Chothia, C. Surface, subunit interfaces and interior of oligomeric proteins. Mol. Biol. 1988, 204, 155–164. [Google Scholar] [CrossRef]
- Chalikian, T.V.; Breslauer, K.J. Thermodynamic analysis of biomolecules: A volumetric approach. Curr. Opin. Struct. Biol. 1998, 8, 657–664. [Google Scholar] [CrossRef]
- Svergun, D.I.; Richard, S.; Koch, M.H.J.; Sayers, Z.; Kuprin, S.; Zaccai, G. Protein hydration in solution: Experimental observation by x-ray and neutron scattering. PNAS 1998, 95, 2267–2272. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Fong, C.; Lamport, T.A. Gum arabic glycoprotein is a twisted hairy rope. A new model based on o-galactosylhydroxyproline as the polysaccharide attachment site. Plant Physiol. 1991, 96, 848–855. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Maiti, P.K.; Cagin, T.; Wang, G.; Goddard, W.A. Structure of pamam dendrimers: Generations 1 through 11. Macromolecules 2004, 37, 6236–6254. [Google Scholar] [CrossRef]
- Sarvazyan, A.P.; Hemmes, P. Relaxational contributios to protein compressibility from ultasonic data. Biopolymers 1979, 18, 3015–3024. [Google Scholar] [CrossRef]
- Marchi, M. Compressibility of cavities and biological water from voronoi volumes in hydrated proteins. J. Phys. Chem. B 2003, 107, 834–836. [Google Scholar] [CrossRef]
- Smolin, N.; Winter, R. A molecular dynamics simulation of snase and its hydration shell at high temperature and high pressure. Biochim. Biophys. Acta 2006, 1764, 522–534. [Google Scholar] [CrossRef] [PubMed]
- Grossutti, M.; Dutcher, J.R. Correlation between chain architecture and hydration water structure in polysaccharides. Biomacromolecules 2016, 17, 1198–1204. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, L.A.; Siani, M.A.; Pique, M.E.; Fisher, C.L.; Getzoff, E.D.; Tainer, J.A. The interdependence of protein surface topography and bound water molecules revealed by surface accessibility and fractal density measures. J. Mol. Biol. 1992, 228, 13–22. [Google Scholar] [CrossRef]
- Smith, A.H.; Lawson, A.W. The velocity of sound in water as a function of temperature and pressur. J. Chem. Phys. 1954, 22, 351–359. [Google Scholar] [CrossRef]
- Leadbetter, A.J. The thermodynamic and vibrational properties of h2o ice and d2o ice. Proc. R. Soc. Lond. 1965, 287, 403–425. [Google Scholar] [CrossRef]
- Chen, Y.-C. Review of Thermal Properties of Snow, Ice and Sea Ice; Cold Regions Research and Engineering Laboratory: Hanover, NH, USA, 1981; pp. 1–35. [Google Scholar]
- Jacobson, B. On the adiabatic compressibility of aqueous solutions. Ark. Kemi 1950, 2, 177–210. [Google Scholar]
- Priev, A.; Almagor, A.; Yedgar, S.; Gavish, B. Glycerol decreases the volume and compressibility of protein interior. Biochemistry 1996, 35, 2061–2066. [Google Scholar] [CrossRef] [PubMed]
- Edwards, C.; Palmer, S.B.; Helliwell, J.R.; Glover, D.; Harris, G.W.; Moss, D.S. Thermal motion in proteins estimated using laser-generated ultrasound and young’s modulus measurements. Acta Crystallogr. 1990, A46, 315–320. [Google Scholar] [CrossRef]
- Morozov, V.N.; Morozova, T.Y. Elasticity of globular proteins. The relation between mechanics, thermodynamics and mobility. J. Biomol. Struct. Dyn. 1993, 11, 459–481. [Google Scholar] [CrossRef] [PubMed]
- Paci, E.; Velikson, B. On the volume of macromolecules. Biopolymers 1996, 41, 785–797. [Google Scholar] [CrossRef]
- Mori, K.; Seki, Y.; Yamada, Y.; Matsumoto, H.; Soda, K. Evaluation of intrinsic compressibility of proteins by molecular dynamics simulations. J. Chem. Phys. 2006, 125. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yamada, H.; Akasaka, K. Effect of pressure on individual hydrogen bonds in proteins: Basic pancreatic trypsin inhibitor. Biochemistry 1998, 37, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Wormald, M.R.; Petrescu, A.J.; Pao, Y.-L.; Glithero, A.; Elliot, T.; Dwek, R.A. Conformational studies of oligosaccharides and glycopeptides: Complementarity of nmr, x-ray crystallography, and molecular modelling. Chem. Rev. 2002, 102, 371–386. [Google Scholar] [CrossRef] [PubMed]
- Phillips, G.O.; Takigami, S.; Takigami, M. Hydration characteristics of the gum exudate from acacia senegal. Food Hydrocoll. 1996, 10, 11–19. [Google Scholar] [CrossRef]
- Nickels, J.D.; Atkinson, J.; Papp-Szabo, E.; Stanley, C.; Diallo, S.O.; Perticaroli, S.; Baylis, B.; Mahon, P.; Ehlers, G.; Katsaras, J.; et al. Structure and hydration of highly-branched, monodisperse phytoglycogen nanoparticles. Biomacromolecules 2016, 17, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Ramadugu, S.K.; Chung, Y.-H.; Xia, J.; Margulis, C.J. When sugars get wet. A comprehensive study of the behavior of water on the surface of oligosaccharides. J. Phys. Chem. B 2009, 113, 11003–11015. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, T.; Uetake, T.; Hatakeayama, H. Frezeeing bound water restrained by gum arabic. In Gums and Stabilizers for the Food Industry; RSC Publishing: Cambridge, UK, 2010; Volume 15, pp. 69–75. [Google Scholar]
- Lupi, L.; Comez, L.; Paolantoni, M.; Perticaroli, S.; Sassi, P.; Morresi, A.; Ladanyi, B.M.; Fioretto, D. Hydration and aggregation in mono- and disaccharide aqueous solutions by gigahertz-to-terahertz light scattering and molecular dynamics simulations. J. Phys. Chem. B 2012, 116, 14760–14767. [Google Scholar] [CrossRef] [PubMed]
- Perticaroli, S.; Nakanishi, M.; Pashkovski, E.; Sokolov, A.P. Dynamics of hydration water in sugars and peptides solutions. J. Phys. Chem. B 2013, 117, 7729–7736. [Google Scholar] [CrossRef] [PubMed]
- Castellani, O.; Al-Assaf, S.; Axelos, M.; Phillips, G.O.; Anton, M. Hydrocolloids with emulsifying capacity. Part 2—Adsorption properties at the n-hexadecane–water interface. Food Hydrocoll. 2010, 24, 121–130. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Component (mg·g−1) | A. senegal | HIC-F1 | HIC-F2 | HIC-F3 | A. seyal |
|---|---|---|---|---|---|
| Total Dry Matter | 893.4 ± 4.0 | 921.6 ± 0.1 | 926.2 ± 1.0 | 921.9 ± 2.0 | 966.9 ± 2.5 |
| Sugars a | 944.4 | 961.3 | 918.3 | 813.0 | 978.0 |
| Arabinose (%) b | 30.2 ± 0.6 | 26.8 ± 1.3 | 35.6 ± 1.0 | 38.3 ± 2.1 | 48.5 ± 1.7 |
| Galactose (%) b | 40.5 ± 1.7 | 39.0 ± 0.8 | 34.4 ± 0.8 | 33.3 ± 1.8 | 34.2 ± 2.0 |
| Rhamnose (%) b | 12.4 ± 0.4 | 12.5 ± 0.1 | 13.7 ± 0.4 | 13.9 ± 1.0 | 3.2 ± 0.7 |
| Glucuronic Acid (%) b | 17.8 ± 1.7 | 20.3 ± 0.6 | 15.6 ± 0.6 | 13.7 ± 2.1 | 7.7 ± 0.4 |
| 4-O-Me-Glucuronic Acid (%) b | 1.0 ± 0.1 | 1.4 ± 0.1 | 0.6 ± 0.1 | 0.7 ± 0.1 | 6.4 ± 0.6 |
| Branching degree d | 0.78 | 0.77 | 0.75 | 0.75 | 0.59 |
| Proteins c | 21.5 ± 0.9 | 4.9 ± 0.1 | 63.1 ± 1.2 | 137.7 ± 2.7 | 7.7 ± 0.0 |
| (27) e | (19) e | (27) e | (32) e | (29) e | |
| Minerals | 34.1 ± 0.1 | 30.5 ± 1.1 | 19.3 ± 1.1 | 49.3 ± 2.6 | 14.3 ± 2.5 |
| A. senegal | HIC-F1 | HIC-F2 | HIC-F3 | A. seyal | |
|---|---|---|---|---|---|
| Mw (g·mol−1) | 6.8 × 105 | 3.5 × 105 | 1.5 × 106 | 1.6 × 106 | 7.1 × 105 |
| Mn (g·mol−1) | 3.1 × 105 | 2.3 × 105 | 1.1 × 106 | 9.0 × 105 | 4.2 × 105 |
| Mw/Mn | 2.0 | 1.4 | 1.3 | 1.9 | 1.5 |
| Mw < 7.5 × 105 g·mol−1 (%) | 86 | 93.0 | 12.3 | 22.7 | 80 |
| Mw > 7.5 × 105 g·mol−1 (%) | 14 | 7.0 | 87.7 | 67.3 | 20 |
| Density (g·cm−3) | 0.99766 | 0.99775 | 0.99759 | 0.99743 | 0.99747 |
| Basic Molecular Characteristics | HIC-F1 | HIC-F2 | HIC-F3 |
|---|---|---|---|
| AGP Mw (g·mol−1) | 348,300 | 1,495,000 | 1,643,000 |
| Polysaccharide moiety Mw (g·mol−1) | 346,593 | 1,400,666 | 1,416,759 |
| Average sugar residue Mw (g·mol−1) | 173.2 | 169.3 | 168.2 |
| Average sugar partial molar volume (cm3·mol−1) | 105.9 | 104.3 | 103.9 |
| Average sugar van der Waals volume (Å3) | 136.9 | 133.8 | 133.0 |
| Number of sugar residues | 2001 | 8441 | 9375 |
| Potential number of charged and polar interacting sites (Polysaccharide moiety) | 6273 | 29,183 | 28,789 |
| Protein moiety Mw (g·mol−1) | 1707 | 94,335 | 226,241 |
| Average aminoacid residue Mw (g·mol−1) | 127.3 | 128.2 | 129.5 |
| Number of aminoacid residues | 13 | 736 | 1747 |
| Charged and polar aminoacids (%) | 80 | 72 | 67 |
| Hydrophobicity index a | −1.46 | −1.01 | −1.14 |
| Potential number of charged and polar interacting sites (Protein moiety) | 12 | 621 | 1391 |
| Type of Acacia Gum or Fraction | vs° (cm3·g−1) | (1011 × cm3·g−1·Pa−1) | βs° (1011 × Pa−1) |
|---|---|---|---|
| A. seyal | 0.5767 | −7.6 | −13.2 |
| A. senegal a | 0.5870 | −7.2 | −12.2 |
| A. senegal | 0.5842 | −7.1 | −12.2 |
| A. senegal b | 0.5940 | −7.5 | −12.5 |
| A. Senegal c | 0.5880 | −7.3 | −12.3 |
| A. Senegal d | 0.5850 | −7.0 | −12.0 |
| HIC-F1 | 0.5616 | −10.3 | −18.3 |
| HIC-F2 | 0.5876 | −8.5 | −14.4 |
| HIC-F3 | 0.6500 | −0.7 | −1.0 |
| Volumetric Properties | HIC-F1 | HIC-F2 | HIC-F3 |
|---|---|---|---|
| Partial molar volumes and related parameters | |||
| Vs° Experimental partial molar volume (cm3·mol−1) | 195,715 | 878,462 | 1,038,047 |
| VM Intrinsic partial molar volume (cm3·mol−1) | 176,296 | 790,616 | 934,243 |
| Vvdw vdW partial molar volume (cm3·mol−1) | 143,530 | 566,467 | 548,311 |
| Vvoid void partial molar volume (cm3·mol−1) | 32,766 | 224,149 | 385,932 |
| VT thermal partial molar volume (cm3·mol−1) | 50,440 | 191,573 | 192,283 |
| Vl interaction partial molar volume (cm3·mol−1) | −30,852 | −103,727 | −90,771 |
| Vsh Partial molar volume hydration water (cm3·mol−1) | 16.298 | 16.320 | 16.333 |
| Decrease of partial molar volume of hydration water (%) | 9.5 | 9.3 | 9.3 |
| Packing density (Vvdw/VM) | 0.81 | 0.72 | 0.60 |
| Void volume (%) | 18.6 | 28.4 | 40.2 |
| Hydration number nh (mole H2O/mole AGP) | 18,128 | 61,748 | 54,444 |
| Hydration number nh (gH2O/gAGP) | 0.85 | 0.68 | 0.54 |
| Hydration number nh (molecule H2O/per residue) | 9.0 | 6.8 | 5.1 |
| Hydration number nh per polysaccharide moiety (gH2O/g AGP) a | 0.88 | 0.72 | 0.62 |
| Hydration number nh per sugar residue of the Polysaccharide moiety (gH2O/gsugar residue) | 8.5 | 6.6 | 5.8 |
| Hydration number nh per protein moiety (gH2O/gAGP) a | 0.44 | 0.36 | 0.31 |
| Hydration number nh per amino acid residue of the Protein moiety (gH2O/gamino acid residue) | 3.9 | 3.6 | 3.4 |
| Partial molar adiabatic compressibility and related parameters | |||
| KM (cm3·mol−1·Pa−1) Intrinsic molar adiabatic compressibility | 1.88 × 10−5 | 1.00 × 10−4 | 1.49 × 10−4 |
| βM (Pa−1) Intrinsic coefficient of adiabatic compressibility | 1.06 × 10−10 | 1.27 × 10−10 | 1.60 × 10−10 |
| Partial molar compressibility hydration water (cm3·mol−1·Pa−1) | 5.20 × 10−9 | 4.60 × 10−9 | 5.30 × 10−9 |
| Partial specific compressibility hydration water (Pa−1) | 2.91 × 10−10 | 2.54 × 10−10 | 2.95 × 10−10 |
| Decrease of partial molar adiabatic compressibility of Hydration water (%) | 37 | 45 | 36 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mejia Tamayo, V.; Nigen, M.; Apolinar-Valiente, R.; Doco, T.; Williams, P.; Renard, D.; Sanchez, C. Flexibility and Hydration of Amphiphilic Hyperbranched Arabinogalactan-Protein from Plant Exudate: A Volumetric Perspective. Colloids Interfaces 2018, 2, 11. https://doi.org/10.3390/colloids2010011
Mejia Tamayo V, Nigen M, Apolinar-Valiente R, Doco T, Williams P, Renard D, Sanchez C. Flexibility and Hydration of Amphiphilic Hyperbranched Arabinogalactan-Protein from Plant Exudate: A Volumetric Perspective. Colloids and Interfaces. 2018; 2(1):11. https://doi.org/10.3390/colloids2010011
Chicago/Turabian StyleMejia Tamayo, Verónica, Michaël Nigen, Rafael Apolinar-Valiente, Thierry Doco, Pascale Williams, Denis Renard, and Christian Sanchez. 2018. "Flexibility and Hydration of Amphiphilic Hyperbranched Arabinogalactan-Protein from Plant Exudate: A Volumetric Perspective" Colloids and Interfaces 2, no. 1: 11. https://doi.org/10.3390/colloids2010011
APA StyleMejia Tamayo, V., Nigen, M., Apolinar-Valiente, R., Doco, T., Williams, P., Renard, D., & Sanchez, C. (2018). Flexibility and Hydration of Amphiphilic Hyperbranched Arabinogalactan-Protein from Plant Exudate: A Volumetric Perspective. Colloids and Interfaces, 2(1), 11. https://doi.org/10.3390/colloids2010011