
Drones, Gulls and Urbanity: Interaction between New Technologies and Human Subsidized Species in Coastal Areas


Abstract
1. Introduction
2. Material and Methods
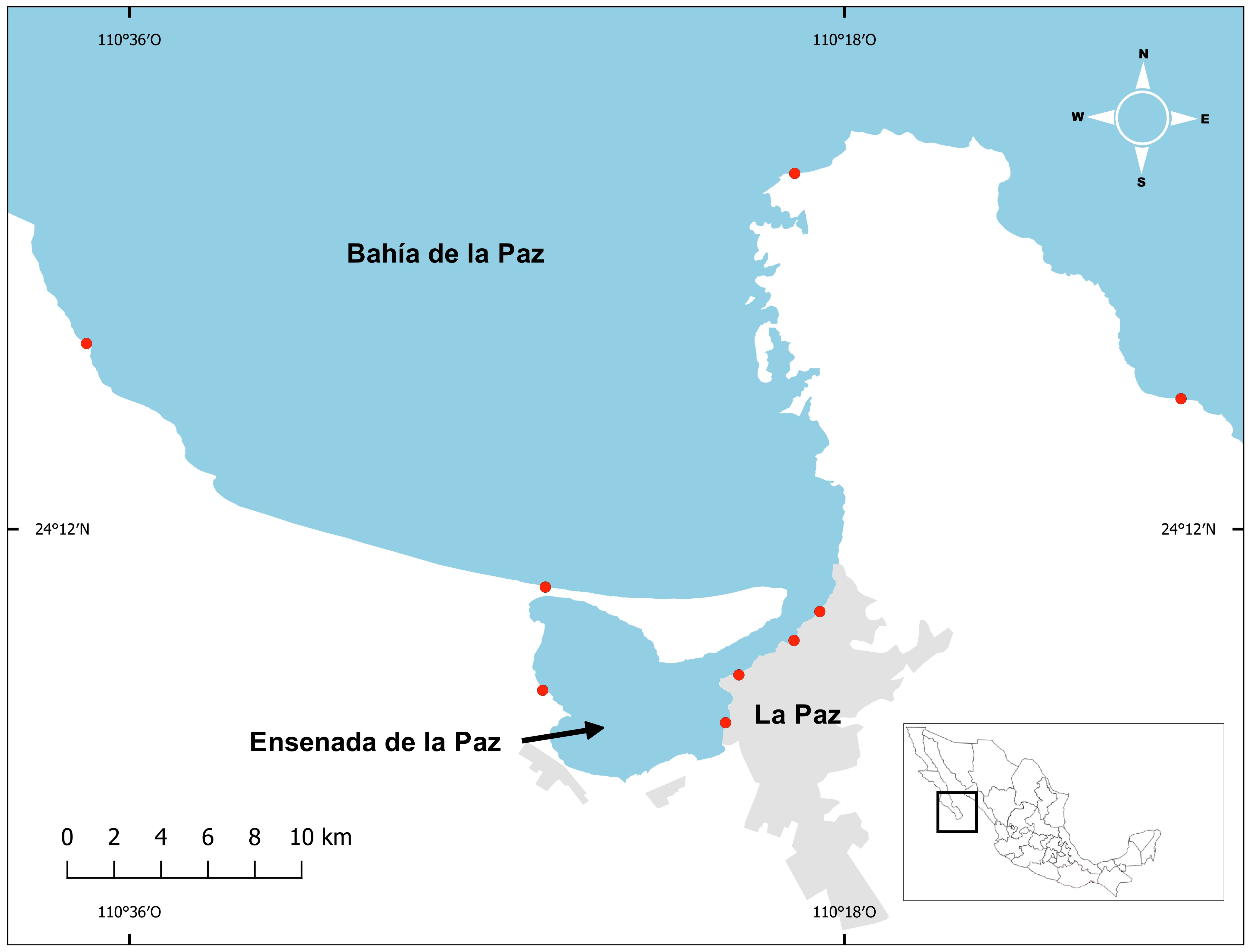
2.1. Study Area
2.2. Field Work and Analysis
2.3. Statistical Analysis
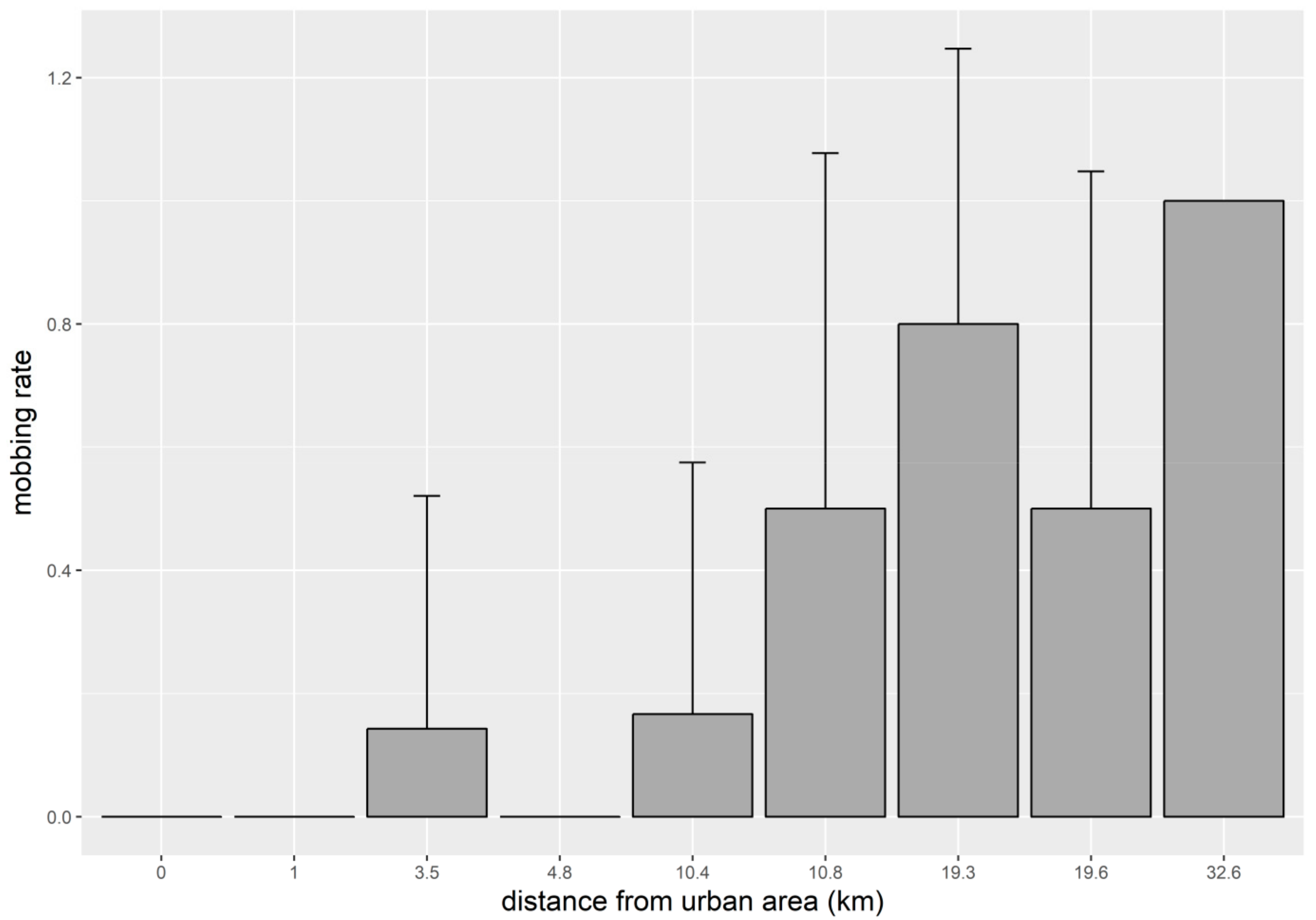
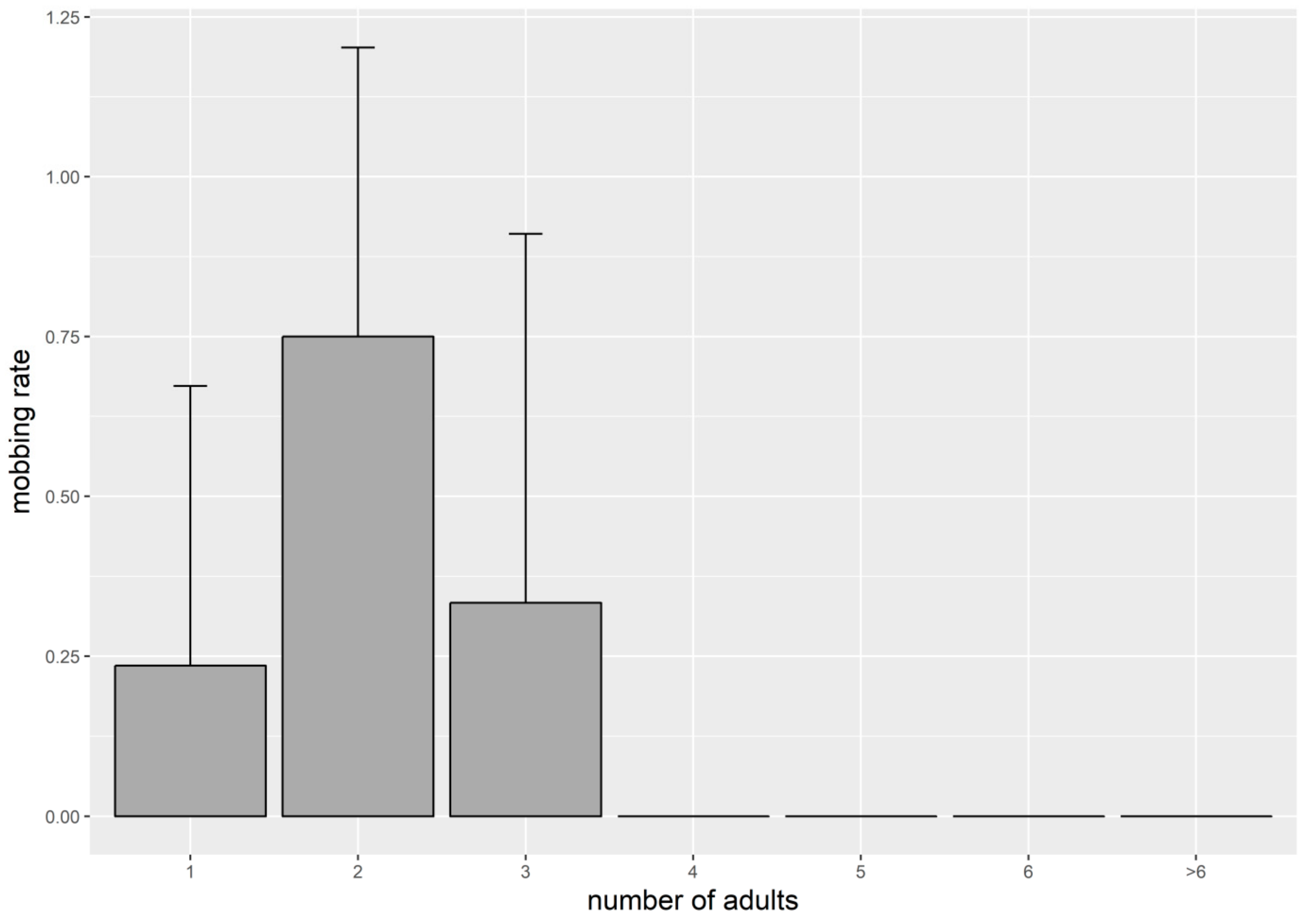
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical Standards
References
- Anderson, K.; Gaston, K.J. Lightweight Unmanned Aerial Vehicles Will Revolutionize Spatial Ecology. Front. Ecol. Environ. 2013, 11, 138–146. [Google Scholar] [CrossRef]
- Park, J.; Kim, S.; Suh, K. A Comparative Analysis of the Environmental Benefits of Drone-Based Delivery Services in Urban and Rural Areas. Sustainability 2018, 10, 888. [Google Scholar] [CrossRef]
- Wang, Z.; Griffin, A.; Lucas, A.; Wong, K.C. Psychological warfare in vineyard: Using drones and bird psychology to control bird damage to wine grapes. Crop Prot. 2019, 120, 163–170. [Google Scholar] [CrossRef]
- Joshi, E.; Sasode, D.S.; Singh, N.; Chouhan, N. Revolution of Indian Agriculture through Drone Technology. Biot. Res. Today 2020, 2, 174–176. [Google Scholar]
- Hodgson, J.C.; Baylis, S.M.; Mott, R.; Herrody, A.; Clarke, R.H. Precision wildlife monitoring using unmanned aerial vehicles. Sci. Rep. 2016, 6, 22574. [Google Scholar] [CrossRef] [PubMed]
- Butcher, P.A.; Colefax, A.P.; Gorkin, R.A.; Kajiura, S.M.; López, N.A.; Mourier, J.; Purcell, C.R.; Skomal, G.B.; Tucker, J.P.; Walsh, A.J.; et al. The Drone Revolution of Shark Science: A Review. Drones 2021, 5, 8. [Google Scholar] [CrossRef]
- Stankov, U.; Kennell, J.; Morrison, A.M.; Vujičić, M.D. The view from above: The relevance of shared aerial drone videos for destination marketing. J. Travel Tour. Mark. 2019, 36, 808–822. [Google Scholar] [CrossRef]
- Provost, E.J.; Butcher, P.A.; Coleman, M.A.; Kelaher, B.P. Assessing the viability of small aerial drones to quantify recreational fishers. Fish. Manag. Ecol. 2020, 27, 615–621. [Google Scholar] [CrossRef]
- Maza, I.; Caballero, F.; Capitán, J.; Martínez-de-Dios, J.; Ollero, A. Experimental results in multi-UAV coordination for disaster management and civil security applications. J. Intell. Robot. Syst. 2011, 61, 563–585. [Google Scholar] [CrossRef]
- Shavarani, S.M.; Nejad, M.G.; Rismanchian, F.; Izbirak, G. Application of hierarchical facility location problem for optimization of a drone delivery system: A case study of Amazon prime air in the city of San Francisco. Int. J. Adv. Manuf. Technol. 2018, 95, 3141–3153. [Google Scholar] [CrossRef]
- Yakushiji, K.; Fujita, H.; Murata, M.; Hiroi, N.; Hamabe, Y.; Yakushiji, F. Short-Range Transportation Using Unmanned Aerial Vehicles (UAVs) during Disasters in Japan. Drones 2020, 4, 68. [Google Scholar] [CrossRef]
- Sedov, L.; Krasnochub, A.; Polishchuk, V. Modeling quarantine during epidemics and mass-testing using drones. PLoS ONE 2020, 15, e0235307. [Google Scholar] [CrossRef]
- Muller, C.G.; Chilvers, B.L.; Barker, Z.; Barnsdale, K.P.; Battley, P.F.; French, R.K.; McCullough, J.; Samandari, F. Aerial VHF tracking of wildlife using an unmanned aerial vehicle (UAV): Comparing efficiency of yellow-eyed penguin (Megadyptes antipodes) nest location methods. Wildl. Res. 2019, 46, 145–153. [Google Scholar] [CrossRef]
- Girault, C. Between naturalness and urbanity, how are protected areas integrated into cities? The case of Helsinki (Finland). J. Urban Res. 2017, 16. [Google Scholar] [CrossRef]
- Floreano, D.; Wood, R.J. Science, technology and the future of small autonomous drones. Nature 2015, 521, 460–466. [Google Scholar] [CrossRef]
- Barr, J.R.; Green, M.C.; De Maso, S.J.; Hardy, T.B. Drone surveys do not increase colony-wide flight behaviour at waterbird nesting sites, but sensitivity varies among species. Sci. Rep. 2020, 10, 3781. [Google Scholar] [CrossRef]
- Rümmler, M.; Mustafa, O.; Maercker, J.; Peter, H.; Esefeld, J. Measuring the influence of unmanned aerial vehicles on Adélie penguins. Polar Biol. 2016, 39, 1329–1334. [Google Scholar] [CrossRef]
- Brisson-Curadeau, É.; Bird, D.; Burke, C.; Fifield, D.A.; Pace, P.; Sherley, R.B.; Elliott, K.H. Seabird species vary in behavioral response to drone census. Sci. Rep. 2017, 7, 17884. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Lim, G.J.; Cho, J.; Côté, M.J. Drone-Aided Healthcare Services for Patients with Chronic Diseases in Rural Areas. J. Intell. Robot. Syst. 2017, 88, 163–180. [Google Scholar] [CrossRef]
- Afán, I.; Máñez, M.; Díaz-Delgado, R. Drone Monitoring of Breeding Waterbird Populations: The Case of the Glossy Ibis. Drones 2018, 2, 42. [Google Scholar] [CrossRef]
- Francis, R.J.; Lyons, M.B.; Kingsford, R.T.; Brandis, K.J. Counting mixed breeding aggregations of animal species using drones: Lessons from waterbirds on semi-automation. Remote Sens. 2020, 12, 1185. [Google Scholar] [CrossRef]
- Mesquita, G.P.; Rodríguez-Teijeiro, J.D.; Wich, S.A.; Mulero-Pázmány, M. Measuring disturbance at a swift breeding colonies due to the visual aspects of a drone: A quasi-experiment study. Curr. Zool. 2020, 67, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Spear, L.B.; Anderson, D. Nest-site selection by Yelow-footed Gulls. Condor 1989, 91, 91–99. [Google Scholar] [CrossRef]
- Lozano, E.; Carmona, R.; Brabata, G. Reproductive success of Heermann’s (Larus heermanni) and Yellow-footed (L livens) gulls in the south of the gulf of California, Mexico. Ornitol. Neotrop. 2004, 15, 237–246. [Google Scholar]
- Hand, J.L.; Hunt, G.L.; Warner, M. Thermal stress and predation: Influences on the structure of gull colony and possibly on breeding distributions. Condor 1981, 83, 193–203. [Google Scholar] [CrossRef]
- Hand, J.L. Human disturbance in Western Gulls Larus occidentalis livens colonies and possible implication by intraespecific predation. Biol. Conserv. 1980, 18, 59–63. [Google Scholar] [CrossRef]
- Baird, R.W. Yellow-footed gull (Larus livens) preys on black storm-petrel (Oceanodroma melania). Colonial Waterbirds 1996, 19, 260–261. [Google Scholar] [CrossRef]
- Flores-Martínez, J.; Gerardo-Herrera, L.; Arroyo-Cabrales, J.; Alarcón, I.; Ruiz, E.A. Seasonal dietary differences of the Yellow-footed Gull (Charadriiformes: Laridae) in Isla Partida Norte, Gulf of California, Mexico. Rev. Mex. Biodivers. 2015, 86, 412–418. [Google Scholar] [CrossRef]
- Adame, K.; Pardo, M.A.; Salvadeo, C.; Beier, E.; Elorriaga-Verplancken, F.R. Detectability and categorization of California sea lions ussing an anmanned aerial vehicle. Mar. Mammal Sci. 2017, 33, 913–925. [Google Scholar] [CrossRef]
- Lowry, H.; Lill, A.; Wong, B.B. Behavioural responses of wildlife to urban environments. Biol. Rev. Camb. Philos. Soc. 2013, 88, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Salgado, R.A.; Bermejo, N.M.; Playa, C.; de Santa Rita, P.; La Paz, B.; de La Paz, H.M.; Esta, F.I.; Vértice, X.Y. Ficha Informativa de los Humedales de RAMSAR (FIR). 2008. Available online: https://rsis.ramsar.org/RISapp/files/RISrep/MX1335RIS.pdf (accessed on 26 March 2021).
- Akaike, H. Information Theory and an Extension of the Maximum Likelihood Principle. In International Symposium on Information Theory; Petrov, B.N., Csaki, F., Eds.; Springer: New York, NY, USA, 1973; pp. 267–281. [Google Scholar]
- Egan, C.C.; Blackwell, B.F.; Fernández-Juricic, E.; Klug, P.E. Testing a key assumption of using drones as frightening devices: Do birds perceive drones as risky? Condor 2020, 122. [Google Scholar] [CrossRef]
- Spelt, A.; Williamson, C.; Shamoun-Baranes, J.; Shepard, E.; Rock, P.; Windsor, S. Habitat use of urban-nesting lesser black-backed gulls during the breeding season. Sci. Rep. 2019, 9, 10527. [Google Scholar] [CrossRef] [PubMed]
- Kasinsky, T.; Suárez, N.; Marinao, C.; Yorio, P. Kelp Gull (Larus dominicanus) Use of Alternative Feeding Habitats at the Bahía San Blas Protected Area, Argentina. Waterbirds 2018, 41, 285–294. [Google Scholar] [CrossRef]
- Marinao, C.; Kasinsky, T.; Suárez, N.; Yorio, P. Contribution of recreational fisheries to the diet of the opportunistic Kelp Gull. Austral Ecol. 2018, 43, 861–875. [Google Scholar] [CrossRef]
- Nunes, L.J.; Raposo, M.A.; Gomes, C.J. The Impact of Tourism Activity on Coastal Biodiversity: A Case Study at Praia da Cova Redonda (Algarve-Portugal). Environments 2020, 7, 88. [Google Scholar] [CrossRef]
- Marrón, G.; Carmona, R.; Ayala, V.; Gutiérrez, G. Noteworthy records of waterbirds in La Paz Bay, Baja California Sur, Mexico. Huit. Rev. Mex. Ornitol. 2014, 15, 57–63. [Google Scholar]
- Silverthorne, K.M.; Perrig, P.L.; Lambertucci, S.A.; Pauli, J.N. Isotopic Niche Partitioning in Avian Scavenger Communities of South America. J. Raptor Res. 2020, 54, 440–446. [Google Scholar] [CrossRef]
- Lisnizer, N.; Yorio, P. Trophic niche expansion during the non-breeding season in kelp gulls of known breeding colony. Mar. Biol. 2019, 166, 12. [Google Scholar] [CrossRef]
- Sol, D.; González-Lagos, C.; Moreira, D.; Maspons, J.; Lapiedra, O. Urbanisation tolerance and the loss of avian diversity. Ecol. Lett. 2014, 17, 942–950. [Google Scholar] [CrossRef]
- Birnie-Gauvin, K.; Peiman, K.S.; Gallagher, A.J.; de Bruijn, R.; Cooke, S.J. Sublethal consequences of urban life for wild vertebrates. Environ. Rev. 2016, 24, 416–425. [Google Scholar] [CrossRef]
- Brousseau, P.; Lefebvre, J.; Giroux, J.F. Diet of ring-billed gull chicks in urban and non-urban colonies in Quebec. Colonial Waterbirds 1996, 19, 22–30. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Model | df | logLik | AICc | ∆ AICc | Weight |
|---|---|---|---|---|---|
| 1) DC + AGS × TGS | 5 | −10.4 | 32.8 | 0 | 0.238 |
| 2) DC + TGS | 3 | −13.6 | 34.1 | 1.27 | 0.126 |
| 2) DC × AGS | 4 | −12.6 | 34.6 | 1.81 | 0.097 |
| 2) DC + TGS + JGS | 4 | −13 | 35.4 | 2.59 | 0.065 |
| 3) DC + AGS | 3 | −14.3 | 35.5 | 2.68 | 0.062 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frixione, M.G.; Salvadeo, C. Drones, Gulls and Urbanity: Interaction between New Technologies and Human Subsidized Species in Coastal Areas. Drones 2021, 5, 30. https://doi.org/10.3390/drones5020030
Frixione MG, Salvadeo C. Drones, Gulls and Urbanity: Interaction between New Technologies and Human Subsidized Species in Coastal Areas. Drones. 2021; 5(2):30. https://doi.org/10.3390/drones5020030
Chicago/Turabian StyleFrixione, Martín G., and Christian Salvadeo. 2021. "Drones, Gulls and Urbanity: Interaction between New Technologies and Human Subsidized Species in Coastal Areas" Drones 5, no. 2: 30. https://doi.org/10.3390/drones5020030
APA StyleFrixione, M. G., & Salvadeo, C. (2021). Drones, Gulls and Urbanity: Interaction between New Technologies and Human Subsidized Species in Coastal Areas. Drones, 5(2), 30. https://doi.org/10.3390/drones5020030