Aging Effects on Gut Microbiota in SAMP8 Mice †


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Aging Effect on Microbiome Composition
2.3. Sample Obtention
2.5. Statistics Analysis
3. Results
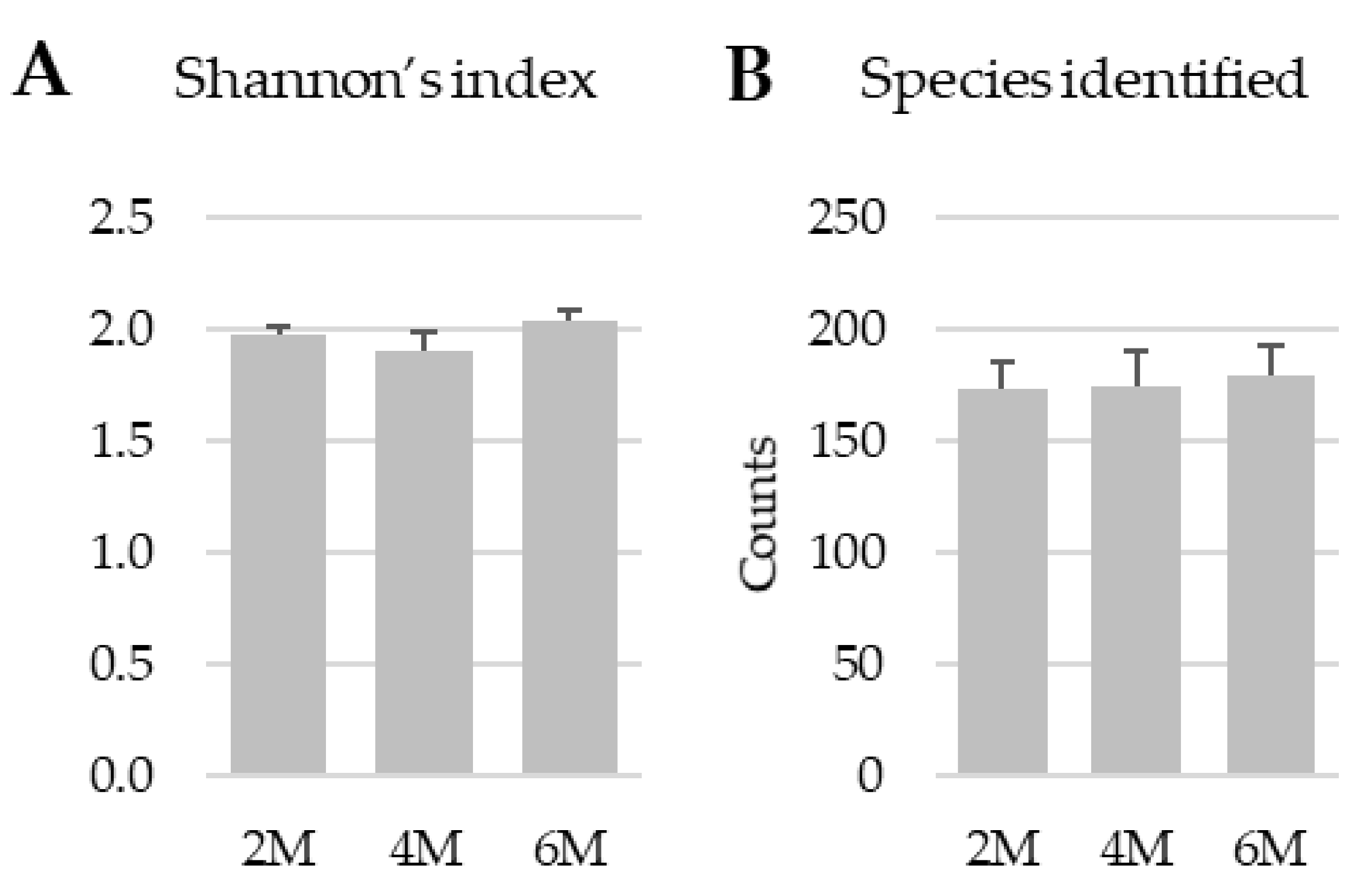
3.1. Shannon Diversity Index and the Number of Species
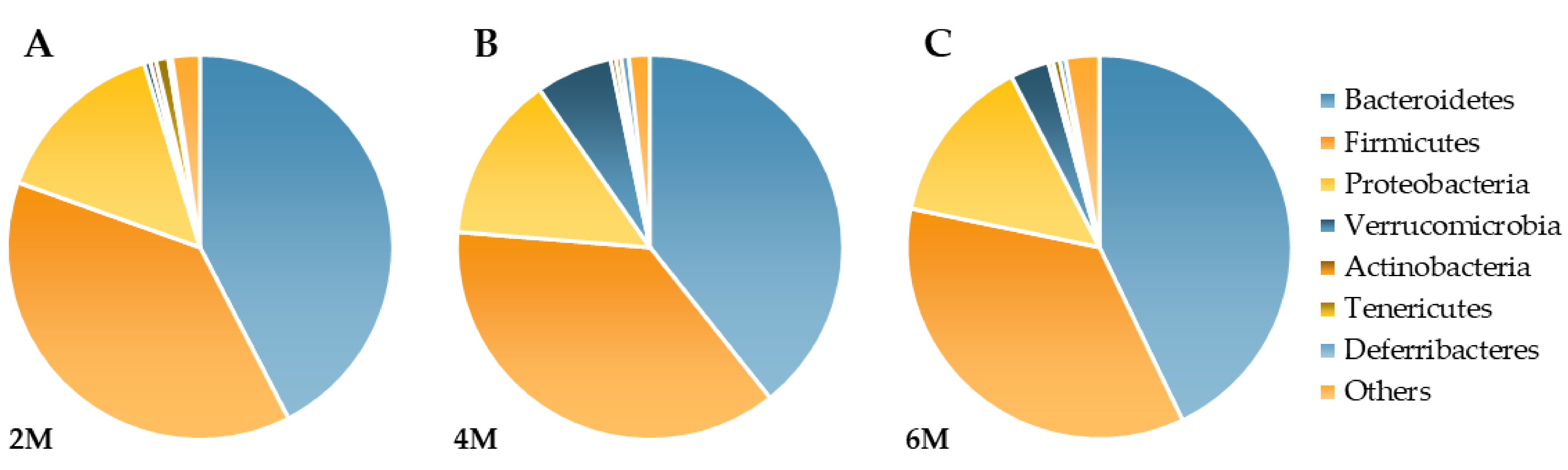
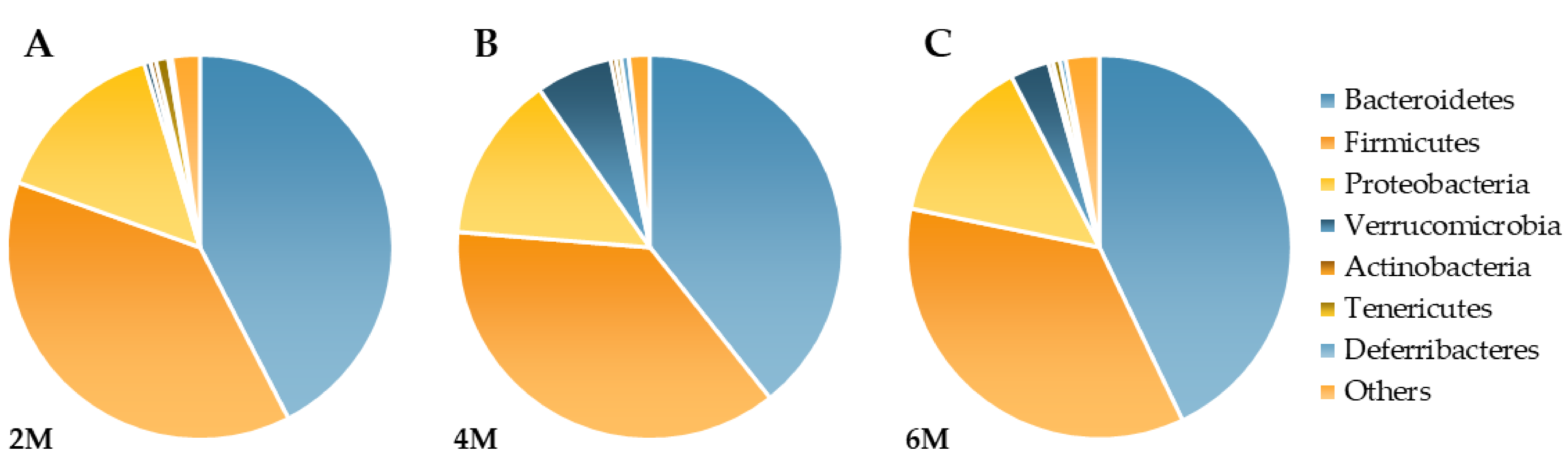
3.2. Phylum Level
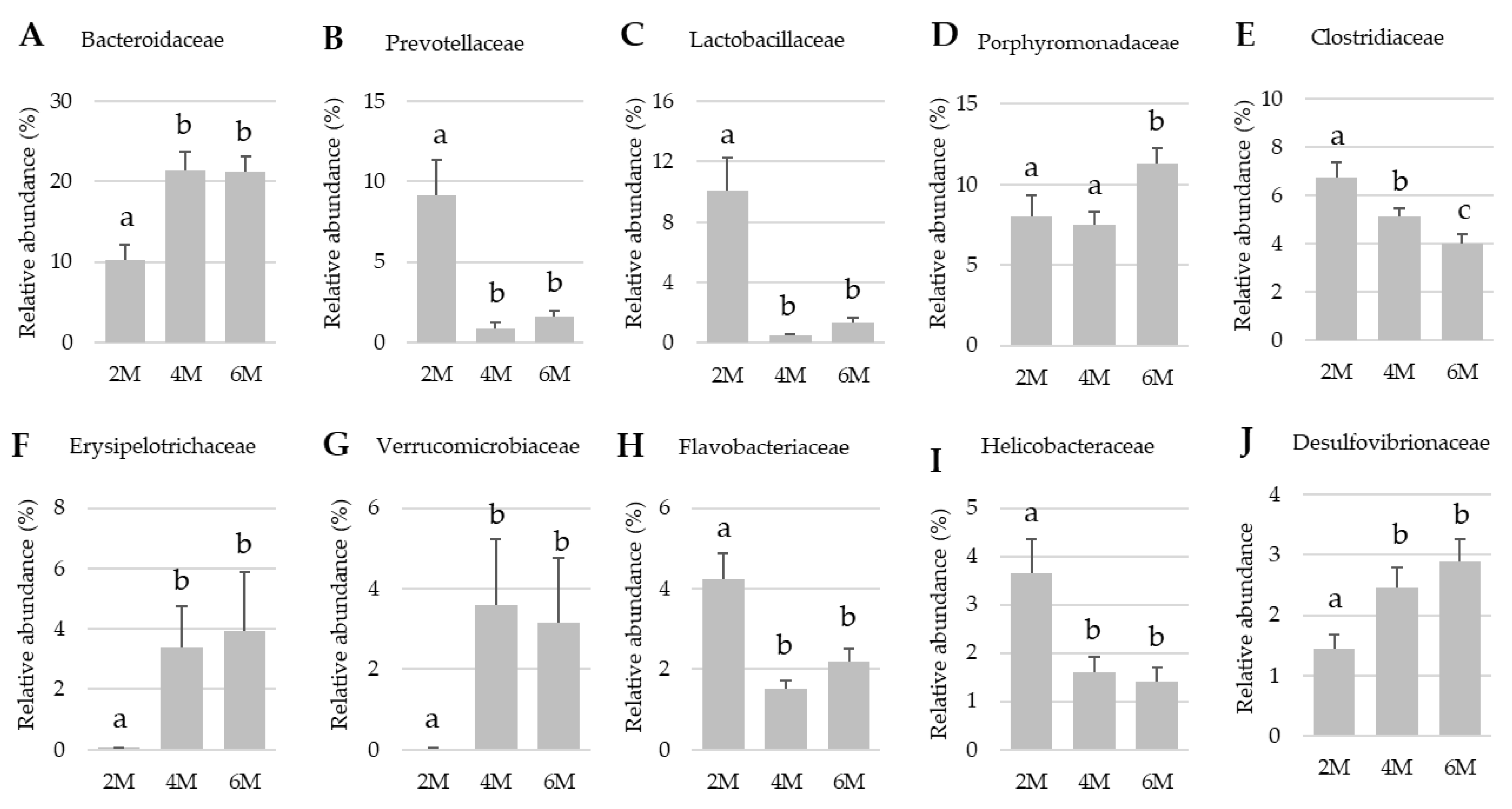
3.3. Family Level Families
3.4. Genus Level
3.5. Species Level
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hooper, L.V.; Littman, D.R.; Macpherson, A.J. Interactions between the microbiota and the immune system. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef] [PubMed]
- Leser, T.D.; Amenuvor, J.Z.; Jensen, T.K.; Lindecrona, R.H.; Boye, M.; Møller, K. Culture-independent analysis of gut bacteria: The pig gastrointestinal tract microbiota revisited. Appl. Environ. Microbiol. 2002, 68, 673–690. [Google Scholar] [CrossRef]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed]
- Rescigno, M. Intestinal microbiota and its effects on the immune system. Cell. Microbiol. 2014, 16, 1004–1013. [Google Scholar] [CrossRef] [PubMed]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef] [PubMed]
- Vaiserman, A.M.; Koliada, A.K.; Marotta, F. Gut microbiota: A player in aging and a target for anti-aging intervention. Ageing Res. Rev. 2017, 35, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Capri, M.; Monti, D.; Giunta, S.; Olivieri, F.; Sevini, F.; Panourgia, M.P.; Invidia, L.; Celani, L.; Scurti, M.; et al. Inflammaging and anti-inflammaging: A systemic perspective on aging and longevity emerged from studies in humans. Mech. Ageing Dev. 2007, 128, 92–105. [Google Scholar] [CrossRef]
- Mullin, J.M.; Valenzano, M.C.; Verrecchio, J.J.; Kothari, R. Age- and diet-related increase in transepithelial colon permeability of Fischer 344 rats. Dig. Dis. Sci. 2002, 47, 2262–2270. [Google Scholar] [CrossRef]
- Tran, L.; Greenwood-Van Meerveld, B. Age-associated remodeling of the intestinal epithelial barrier. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1045–1056. [Google Scholar] [CrossRef]
- Miró, L.; Garcia-Just, A.; Amat, C.; Polo, J.; Moretó, M.; Pérez-Bosque, A. Dietary animal plasma proteins improve the intestinal immune response in senescent mice. Nutrients 2017, 9, 1346. [Google Scholar] [CrossRef]
- Garcia-Just, A.; Miró, L.; Pérez-Bosque, A.; Amat, C.; Polo, J.; Pallàs, M.; Griñán-Ferré, C.; Moretó, M. Dietary Spray-Dried Porcine Plasma Prevents Cognitive Decline in Senescent Mice and Reduces Neuroinflammation and Oxidative Stress. J. Nutr. 2020, 150, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Konturek, P.C.; Haziri, D.; Brzozowski, T.; Hess, T.; Heyman, S.; Kwiecien, S.; Konturek, S.J.; Koziel, J. Emerging role of fecal microbiota therapy in the treatment of gastrointestinal and extra-gastrointestinal diseases. J. Physiol. Pharmacol. 2015, 66, 483–491. [Google Scholar] [PubMed]
- Bischoff, S.C. Microbiota and aging. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Pérez Martínez, G.; Bäuerl, C.; Collado, M.C. Understanding gut microbiota in elderly’s health will enable intervention through probiotics. Benef. Microbes 2014, 5, 235–246. [Google Scholar] [CrossRef]
- Moretó, M.; Miró, L.; Amat, C.; Polo, J.; Manichanh, C.; Pérez-Bosque, A. Dietary supplementation with spray-dried porcine plasma has prebiotic effects on gut microbiota in mice. Sci. Rep. 2020, 10, 2926. [Google Scholar] [CrossRef]
- Larbi, A.; Pawelec, G.; Wong, S.C.; Goldeck, D.; Tai, J.J.; Fulop, T. Impact of age on T cell signaling: A general defect or specific alterations? Ageing Res. Rev. 2011, 10, 370–378. [Google Scholar] [CrossRef]
- Franceschi, C.; Salvioli, S.; Garagnani, P.; de Eguileor, M.; Monti, D.; Capri, M. Immunobiography and the Heterogeneity of Immune Responses in the Elderly: A Focus on Inflammaging and Trained Immunity. Front. Immunol. 2017, 8, 982. [Google Scholar] [CrossRef]
- Larbi, A.; Franceschi, C.; Mazzatti, D.; Solana, R.; Wikby, A.; Pawelec, G. Aging of the immune system as a prognostic factor for human longevity. Physiology 2008, 23, 64–74. [Google Scholar] [CrossRef]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 9, 70–89. [Google Scholar] [CrossRef]
- Claesson, M.J.; Cusack, S.; O’Sullivan, O.; Greene-Diniz, R.; de Weerd, H.; Flannery, E.; Marchesi, J.R.; Falush, D.; Dinan, T.; Fitzgerald, G.; et al. Composition, variability, and temporal stability of the intestinal microbiota of the elderly. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4586–4591. [Google Scholar] [CrossRef]
- Hopkins, M.J.; Sharp, R.; Macfarlane, G.T. Age and disease related changes in intestinal bacterial populations assessed by cell culture, 16S rRNA abundance, and community cellular fatty acid profiles. Gut 2001, 48, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Claesson, M.J.; Jeffery, I.B.; Conde, S.; Power, S.E.; O’Connor, E.M.; Cusack, S.; Harris, H.M.; Coakley, M.; Lakshminarayanan, B.; O’Sullivan, O.; et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012, 488, 178–184. [Google Scholar] [CrossRef] [PubMed]
- McDonald, K.G.; Leach, M.R.; Huang, C.; Wang, C.; Newberry, R.D. Aging impacts isolated lymphoid follicle development and function. Immun. Ageing 2011, 8, 1. [Google Scholar] [CrossRef]
- Rehman, T. Role of the gut microbiota in age-related chronic inflammation. Endocr. Metab. Immune Disord. Drug Targets 2012, 12, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Odamaki, T.; Kato, K.; Sugahara, H.; Hashikura, N.; Takahashi, S.; Xiao, J.Z.; Abe, F.; Osawa, R. Age-related changes in gut microbiota composition from newborn to centenarian: A cross-sectional study. BMC Microbiol. 2016, 16, 90. [Google Scholar] [CrossRef] [PubMed]
- Thevaranjan, N.; Puchta, A.; Schulz, C.; Naidoo, A.; Szamosi, J.C.; Verschoor, C.P.; Loukov, D.; Schenck, L.P.; Jury, J.; Foley, K.P.; et al. Age-Associated Microbial Dysbiosis Promotes Intestinal Permeability, Systemic Inflammation, and Macrophage Dysfunction. Cell Host Microbe 2017, 21, 455–466. [Google Scholar] [CrossRef]
- Fransen, F.; van Beek, A.A.; Borghuis, T.; Aidy, S.E.; Hugenholtz, F.; van der Gaast-de Jongh, C.; Savelkoul, H.F.J.; De Jonge, M.I.; Boekschoten, M.V.; Smidt, H.; et al. Aged Gut Microbiota Contributes to Systemical Inflammaging after Transfer to Germ-Free Mice. Front. Immunol. 2017, 8, 1385. [Google Scholar] [CrossRef]
- Håkansson, Å.; Tormo-Badia, N.; Baridi, A.; Xu, J.; Molin, G.; Hagslätt, M.L.; Karlsson, C.; Jeppsson, B.; Cilio, C.M.; Ahrné, S. Immunological alteration and changes of gut microbiota after dextran sulfate sodium (DSS) administration in mice. Clin. Exp. Med. 2015, 15, 107–120. [Google Scholar] [CrossRef]
- Rowan, F.; Docherty, N.G.; Murphy, M.; Murphy, B.; Calvin Coffey, J.; O’Connell, P.R. Desulfovibrio bacterial species are increased in ulcerative colitis. Dis. Colon Rectum 2010, 53, 1530–1536. [Google Scholar] [CrossRef]
- Mukhopadhya, I.; Hansen, R.; El-Omar, E.M.; Hold, G.L. IBD-what role do Proteobacteria play? Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 219–230. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Rodriguez-Pineiro, A.M.; Schutte, A.; Ermund, A.; Boysen, P.; Bemark, M.; Sommer, F.; Bäckhed, F.; Hansson, G.C.; Johansson, M.E. The composition of the gut microbiota shapes the colon mucus barrier. EMBO Rep. 2014, 16, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Ottman, N.; Davids, M.; Suarez-Diez, M.; Boeren, S.; Schaap, P.J.; Martins Dos Santos, V.A.P.; Smidt, H.; Belzer, C.; de Vos, W.M. Genome-scale model and omics analysis of metabolic capacities of Akkermansia muciniphila reveal a preferential mucin-degrading lifestyle. Appl. Environ. Microbiol. 2017, 83, 1014–1017. [Google Scholar] [CrossRef] [PubMed]
- Walrath, T.; Dyamenahalli, K.U.; Hulsebus, H.J.; McCullough, R.L.; Idrovo, J.P.; Boe, D.M.; McMahan, R.H.; Kovacs, E.J. Age-related changes in intestinal immunity and the microbiome. J. Leukoc. Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.M.; Schafer, M.J.; Sohn, J.; Vincentini, J.; Weiner, H.L.; Ginsberg, S.D.; Blaser, M.J. Calorie restriction slows age-related microbiota changes in an Alzheimer’s disease model in female mice. Sci. Rep. 2019, 9, 17904. [Google Scholar] [CrossRef]
- Berer, K.; Gerdes, L.A.; Cekanaviciute, E.; Jia, X.; Xiao, L.; Xia, Z.; Liu, C.; Klotz, L.; Stauffer, U.; Baranzini, S.E.; et al. Gut microbiota from multiple sclerosis patients enables spontaneous autoimmune encephalomyelitis in mice. Proc. Natl. Acad. Sci. USA 2017, 114, 10719–10724. [Google Scholar] [CrossRef]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V.; et al. Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson’s Disease. Cell 2016, 167, 1469–1480. [Google Scholar] [CrossRef]
- Sharon, G.; Cruz, N.J.; Kang, D.W.; Gandal, M.J.; Wang, B.; Kim, Y.M.; Zink, E.M.; Casey, C.P.; Taylor, B.C.; Lane, C.J.; et al. Human Gut Microbiota from Autism Spectrum Disorder Promote Behavioral Symptoms in Mice. Cell 2019, 177, 1600–1618.e17. [Google Scholar] [CrossRef]
- Cattaneo, A.; Cattane, N.; Galluzzi, S.; Provasi, S.; Lopizzo, N.; Festari, C.; Ferrari, C.; Guerra, U.P.; Paghera, B.; Muscio, C.; et al. Association of brain amyloidosis with pro-inflammatory gut bacterial taxa and peripheral inflammation markers in cognitively impaired elderly. Neurobiol. Aging 2017, 49, 60–68. [Google Scholar] [CrossRef]
- Zhao, Y.; Lukiw, W.J. Bacteroidetes Neurotoxins and Inflammatory Neurodegeneration. Mol. Neurobiol. 2018, 55, 9100–9107. [Google Scholar] [CrossRef]
- Tran, T.T.T.; Corsini, S.; Kellingray, L.; Hegarty, C.; Le Gall, G.; Narbad, A.; Müller, M.; Tejera, N.; O’Toole, P.W.; Minihane, A.M.; et al. APOE genotype influences the gut microbiome structure and function in humans and mice: Relevance for Alzheimer’s disease pathophysiology. FASEB J. 2019, 33, 8221–8231. [Google Scholar] [CrossRef]
- Forsyth, C.B.; Shannon, K.M.; Kordower, J.H.; Voigt, R.M.; Shaikh, M.; Jaglin, J.A.; Estes, J.D.; Dodiya, H.B.; Keshavarzian, A. Increased intestinal permeability correlates with sigmoid mucosa alpha-synuclein staining and endotoxin exposure markers in early Parkinson’sdisease. PLoS ONE 2011, 6, e28032. [Google Scholar] [CrossRef] [PubMed]
- Crews, L.; Tsigelny, I.; Hashimoto, M.; Masliah, E. Role of synucleins in Alzheimer’s disease. Neurotox. Res. 2009, 16, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Albenberg, L.G.; Wu, G.D. Diet and the intestinal microbiome: Associations, functions, and implications for health and disease. Gastroenterology 2014, 146, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.Q.; Shen, L.L.; Li, W.W.; Fu, X.; Zeng, F.; Gui, L.; Lü, Y.; Cai, M.; Zhu, C.; Tan, Y.L.; et al. Gut Microbiota is Altered in Patients with Alzheimer’s Disease. J. Alzheimers Dis. 2018, 63, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Junges, V.M.; Closs, V.E.; Nogueira, G.M.; Gottlieb, M.G.V. Crosstalk between gut microbiota and the central nervous system: A focus for Alzheimer’s disease. Curr. Alzheimer Res. 2018, 15, 1179–1190. [Google Scholar] [CrossRef]
- Bajaj, J.S.; Hylemon, P.B.; Ridlon, J.M.; Heuman, D.M.; Daita, K.; White, M.B.; Monteith, P.; Noble, N.A.; Sikaroodi, M.; Gillevet, P.M. Colonic mucosal microbiome differs from stool microbiome in cirrhosis and hepatic encephalopathy and is linked to cognition and inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G675–G685. [Google Scholar] [CrossRef]
- Askarova, S.; Umbayev, B.; Masoud, A.R.; Kaiyrlykyzy, A.; Safarova, Y.; Tsoy, A.; Olzhayev, F.; Kushugulova, A. The Links Between the Gut Microbiome, Aging, Modern Lifestyle and Alzheimer’s Disease. Front. Cell. Infect. Microbiol. 2020, 10, 104. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Phylum | Family | Genus | 2M | 4 M | 6M |
|---|---|---|---|---|---|
| Bacteroidetes | Bacteroidaceae | Bacteroides | 11.1 ± 2.0 a | 23.0 ± 2.5 b | 22.9 ± 2.1 b |
| Sphingobacteriaceae | Pedobacter | 1.75 ± 0.11 a | 0.93 ± 0.08 b | 1.00 ± 0.10 b | |
| Olivibacter | 1.36 ± 0.19 a | 0.77 ± 0.06 b | 0.73 ± 0.09 b | ||
| Odoribacteraceae | Odoribacter | 1.95 ± 0.31 a | 0.15 ± 0.02 b | 0.14 ± 0.02 b | |
| Butyricimonas | 0.40 ± 0.07 a | 1.32 ± 0.26 b | 1.59 ± 0.28 b | ||
| Prevotellaceae | Prevotella | 9.98 ± 2.40 a | 0.98 ± 0.39 b | 1.75 ± 0.40 b | |
| Porphyromonadaceae | Parabacteroides | 3.48 ± 0.45 a | 3.44 ± 0.47 a | 6.57 ± 0.89 b | |
| Flavobacteriaceae | Flavobacterium | 3.95 ± 0.65 a | 1.10 ± 0.19 b | 1.69 ± 0.24 b | |
| Firmicutes | Lactobacillaceae | Lactobacillus | 8.85 ± 1.99 a | 0.84 ± 0.34 b | 1.15 ± 0.29 b |
| Pediococcus | 0.85 ± 0.18 a | 0.14 ± 0.02 b | 0.16 ± 0.03 b | ||
| Lachnospiraceae | Lachnospira | 0.39 ± 0.06 a | 0.10 ± 0.02 b | 0.11 ± 0.04 b | |
| Erysipelotrichaceae | Erysipelothrix | 0.06 ± 0.01 a | 3.34 ± 1.34 b | 4.05 ± 2.05 b | |
| Clostridiaceae | Clostridium | 1.53 ± 0.16 a | 1.16 ± 0.12 b | 0.92 ± 0.13 c | |
| Natronincola | 2.46 ± 0.30 a | 1.21 ± 0.30 b | 0.92 ± 0.26 b | ||
| Proteobacteria | Helicobacteraceae | Helicobacter | 3.92 ± 0.76 a | 1.71 ± 0.34 b | 1.53 ± 0.32 b |
| Desulfovibrionaceae | Desulfovibrio | 1.53 ± 0.26 a | 2.06 ± 0.36 b | 3.10 ± 0.43 b | |
| Verrucomicrobia | Akkermansiaceae | Akkermansia | 0.01 ± 0.00 a | 2.38 ± 1.06 b | 3.26 ± 1.50 b |
| Phylum | Family | Species | 2M | 4 M | 6M |
|---|---|---|---|---|---|
| Bacteroidetes | Bacteroidaceae | Bacteroides xylanisolvens | 2.90 ± 0.45 a | 7.94 ± 0.90 b | 6.62 ± 1.16 b |
| Bacteroides rodentium | 2.74 ± 0.61 a | 7.49 ± 1.52 b | 3.16 ± 0.90 a | ||
| Bacteroides acidifaciens | 0.54 ± 0.11 a | 5.94 ± 0.96 b | 4.11 ± 0.70 b | ||
| Bacteroides denticanum | 0.86 ± 0.12 a | 3.98 ± 1.36 b | 6.55 ± 1.11 c | ||
| Bacteroides sartorii | 0.31 ± 0.07 a | 1.26 ± 0.25 b | 1.08 ± 0.29 b | ||
| Bacteroides graminisolvens | 0.17 ± 0.03 a | 0.89 ± 0.12 b | 1.02 ± 0.18 b | ||
| Odoribacteraceae | Odoribacter denticanis | 2.57 ± 0.40 a | 0.22 ± 0.06 b | 0.20 ± 0.03 b | |
| Butyricimonas virosa | 0.19 ± 0.03 a | 1.30 ± 0.22 b | 0.85 ± 0.20 c | ||
| Prevotellaceae | Prevotella dentasini | 16.1 ± 3.73 a | 1.12 ± 0.43 b | 1.63 ± 0.40 b | |
| Porphyromonadaceae | Parabacteroides gordonii | 0.00 ± 0.00 a | 0.01 ± 0.01 a | 3.57 ± 0.81 b | |
| Firmicutes | Lactobacillaceae | Lactobacillus hayakitensis | 5.13 ± 1.75 a | 0.33 ± 0.13 b | 0.86 ± 0.21 b |
| Lactobacillus taiwanensis | 1.93 ± 0.67 a | 0.03 ± 0.01 b | 0.06 ± 0.03 b | ||
| Lactobacillus siliginis | 1.77 ± 0.48 a | 0.19 ± 0.07 b | 0.28 ± 0.06 b | ||
| Lactobacillus intermedius | 0.85 ± 0.44 a | 0.06 ± 0.02 b | 0.11 ± 0.03 b | ||
| Pediococcus argentinicus | 0.45 ± 0.14 a | 0.02 ± 0.01 b | 0.02 ± 0.01 b | ||
| Lachnospiraceae | Blautia hansenii | 1.68 ± 0.37 a | 0.65 ± 0.07 b | 0.95 ± 0.23 b | |
| Ruminococcus gnavus | 0.24 ± 0.06 a | 1.08 ± 0.29 b | 0.64 ± 0.22 ab | ||
| Johnsonella ignava | 2.99 ± 0.56 a | 0.31 ± 0.06 b | 0.41 ± 0.11 b | ||
| Erysipelotrichaceae | Erysipelothrix muris | 0.10 ± 0.02 a | 5.54 ± 2.18 b | 1.66 ± 0.83 b | |
| Ruminococcaceae | Oscillospira guilliermondii | 1.03 ± 0.12 a | 1.82 ± 0.29 b | 1.18 ± 0.28 a | |
| Proteobacteria | Helicobacteraceae | Helicobacter mastomyrinus | 4.13 ± 0.64 a | 1.21 ± 0.24 b | 1.65 ± 0.50 b |
| Desulfovibrionaceae | Desulfovibrio psychrotolerans | 0.26 ± 0.06 a | 1.32 ± 0.58 ab | 2.31 ± 0.48 b | |
| Verrucomicrobia | Akkermansiaceae | Akkermansia muciniphila | 0.01 ± 0.00 a | 7.17 ± 2.48 b | 4.75 ± 2.20 b |
| Verrucomicrobiaceae | Luteolibacter algae | 0.01 ± 0.00 a | 1.86 ± 0.68 b | 0.69 ± 0.38 a,b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miró, L.; Moretó, M.; Amat, C.; Polo, J.; Pérez-Bosque, A. Aging Effects on Gut Microbiota in SAMP8 Mice. Proceedings 2020, 61, 25. https://doi.org/10.3390/IECN2020-06995
Miró L, Moretó M, Amat C, Polo J, Pérez-Bosque A. Aging Effects on Gut Microbiota in SAMP8 Mice. Proceedings. 2020; 61(1):25. https://doi.org/10.3390/IECN2020-06995
Chicago/Turabian StyleMiró, Lluïsa, Miquel Moretó, Concepció Amat, Javier Polo, and Anna Pérez-Bosque. 2020. "Aging Effects on Gut Microbiota in SAMP8 Mice" Proceedings 61, no. 1: 25. https://doi.org/10.3390/IECN2020-06995
APA StyleMiró, L., Moretó, M., Amat, C., Polo, J., & Pérez-Bosque, A. (2020). Aging Effects on Gut Microbiota in SAMP8 Mice. Proceedings, 61(1), 25. https://doi.org/10.3390/IECN2020-06995