2. Results
2.1. Testudinidae
Remarks: Prominent costal wedging is a trait often associated with Testudinidae. We use this in the same way Hay did [
2]; it is associated with the proximal widening of costals 3 and 5 and the proximal narrowing of costals 2, 4, and 6.
2.2. Hadrianus corsoni
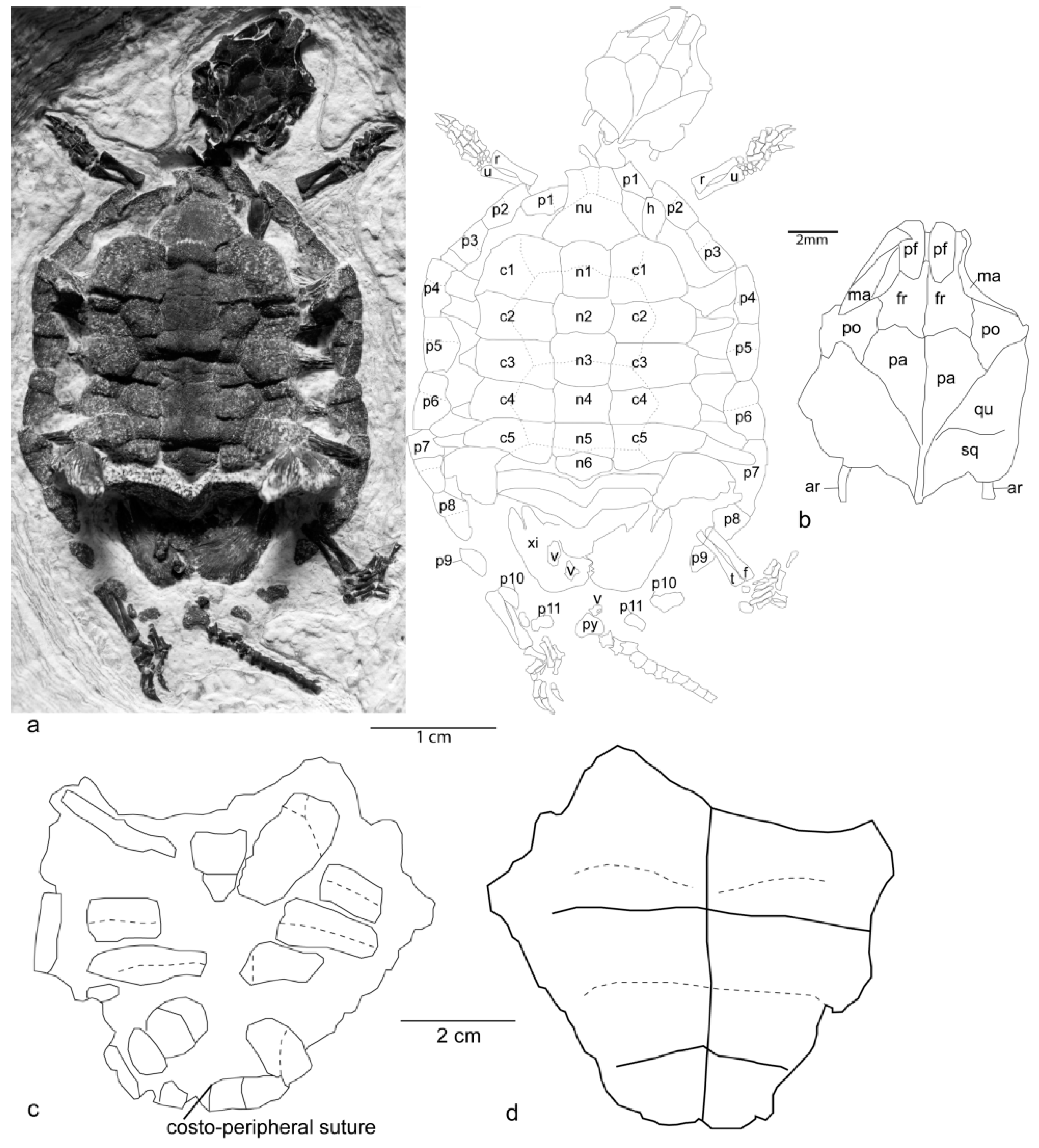
New material: Reconstruction of a hatchling turtle skeleton identified as
Hadrianus corsoni (FOBU 14015,
Figure 1) is nearly complete, including the skull, carapace, posterior two-thirds of the plastron, and incomplete limbs.
Skull: The skull (
Figure 1) is 13 mm long and 9.7 mm wide. The frontal forms the posterior half of the dorsal margin of the orbit. The frontals have a distinct anteromedial projection between the prefrontals. The orbits are anteriorly placed just behind the anterior tip of the snout. The parietals are large, forming most of the posterior portion of the skull roof. The postorbital is large and expanded laterally, is triangular, and is sutured to the parietal and frontal. It forms the entire posterior margin of the orbit. The prefrontals are roughly rectangular, forming the dorsal anterior portion of the orbital margin. The maxilla forms much of the lateral margin of the nasal passage and the floor of the orbit. The squamosal and quadrate suture is about halfway between the posterior margin of the postorbital and the posterior extremity of the supraoccipital. The articular of each side of the mandible is below the skull, about 3 mm lateral to either side of the supraoccipital.
Carapace: The carapace (
Figure 1) is 40 mm long and 31.3 mm wide. It has the distinct medial ridge typical of many juvenile testudinoids. The posterior three peripherals and the pygal are poorly ossified, and do not contact each other. Costal wedging is present, and the neurals and proximal portions of the costals show the typical striated surface sculpture of Hadrianus. The sulci are deeply incised into the carapace. The nuchal is well-developed, contacting neural 1, both first peripherals, and both first costals. Neural 1 is rectangular, 6.3 mm long and 5.8 mm wide, contacts costal 1, and is crossed by the vertebral 1–2 sulcus. Neural 2 is hexagonal, 2.5 mm long and 8.1 mm wide, contacts costals 1 and 2, and is not crossed by sulci. Neural 3 is hexagonal, with short sides paralleling the midline near the crossing of the vertebral 2–3 sulcus. It is 5 mm long and 4.4 mm wide. Neural 4 is hexagonal, with extremely short anterior lateral sides, and is 5 mm long and 5 mm wide. It lacks any crossing sulcus and contacts costals 3, 4, and 5, although the contact with costal 5 is small. Neural 5 is rectangular, and very slightly elongated anteriorly and posteriorly along the midline. It is crossed by the vertebral 3–4 sulcus. All three vertebral–vertebral sulci bow anteriorly as they cross the midline, creating a slight “omega” shape. Neural 6 is tilted nearly vertical, so some of its features are obscured; however, it appears as rectangular, with no division of the lateral sides. The neural formula up to neural 6 is 4-6A-6A-6A-4-4. The lateral margin of the carapace is slightly concave between peripherals 5 and 6. The greatest width is at the peripheral 6–7 suture.
Plastron: Both the hyoplastron–hypoplastron and the hypoplastron–xiphiplastron fontanelles are open. The midline suture is loose and unsutured in some areas. The inguinal buttress is well ossified between the distal third of costals 5 and 6, and was pushed through the carapace taphonomically.
Limbs: The forelimbs are nearly complete, but the humerus is partly covered by the anterior peripherals. Both ulnae are 4.4 mm long, and the hands are 6 mm long. The distal phalanges have a small flexor tubercle. The phalangeal formula is 0-3-3-3-3. The left hind limb is relatively complete, including a 5 mm long tibia and a 6 mm long pes. The distal phalanges of the pes lack a well-defined flexor tubercle. Neither pes preserves full digits 1 or 2. The phalangeal formula is ?-?-3-3-3.
2.3. Hadrianus majusculus
New material: UCB 76816; incomplete carapace and plastron, missing the anterior and posterior ends of the plastron, as well as most peripherals; the nuchal; and the pygal. Distribution: Wasatchian (Wa-5) of the Willwood Formation of Wyoming, USA.
Description: An incomplete shell (
Figure 1) identified as
Hadrianus majusculus based on: (1) lack of costal wedging; (2) posteriorly concave pectoral–abdominal sulcus; and (3) narrow base of anterior lobe of plastron relative to base of posterior lobe. The hyoplastron measures 20 mm long and the hypoplastron is 21 mm long. The hyoplastron has four scute annuli, indicating an individual approximately 4 years old [
4].
2.4. Echmatemys lativertebralis (Family Unconfirmed)
Distribution: Early Eocene (Wasatchian), San Jose Formation, New Mexico; Wasatch Formation, Wyoming; DeBeque Formation, Colorado.
Remarks:
Echmatemys lativertebralis is more distinct from the other species of Echmatemys than they are from each other.
E. lativertebralis averages 250 mm carapace length. Vlachos [
5] recognizes
Echmatemys cibollensis as distinct from E. lativertebralis, but this is only based on the extent of the gulars overlapping the entoplastron, which varies in all testudinoid species.
2.5. Echmatemys haydeni
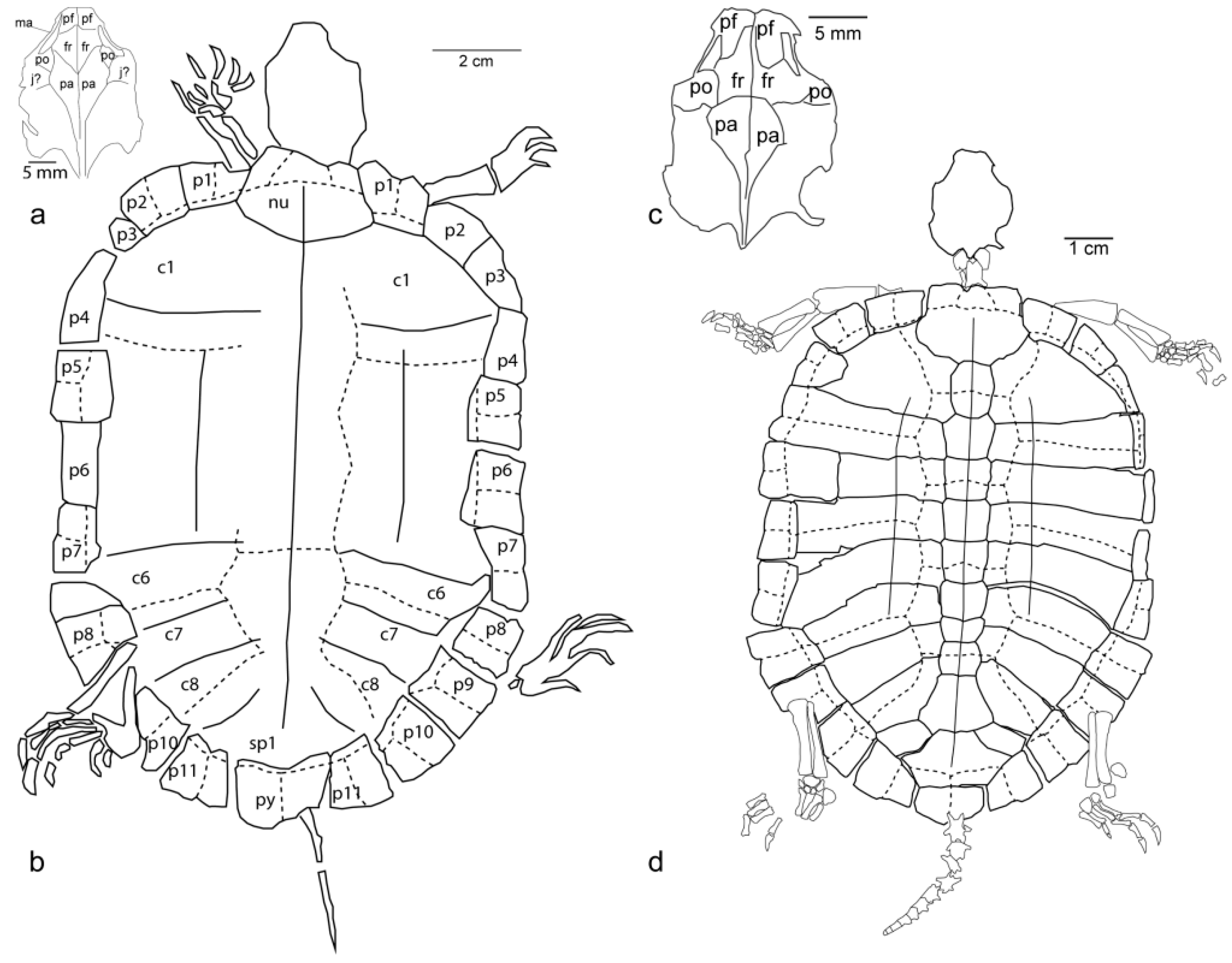
Referred specimen: An articulated skeleton from the Green River Formation (TMP 2008.00.14,
Figure 2) includes the first known skull of Echmatemys haydeni. Identification is based on the long contact of marginal 1 and pleural 1, as well as the overlap of the posterior sulcus of vertebral 5 over the pygal. The large medial extent of the inguinal buttresses due to the costals separating them nearly to the costal–neural sutures is unusual.
Description: The skull is 31 mm long, 21 mm wide, and dorsoventrally compressed, with most elements below the skull table obscured. The postorbitals are reduced, with another bone (possibly jugal) forming much of the ventral posterior margin of the orbit. The anterior process of the frontal is small relative to that of
Echmatemys naomi. The frontals lie rostrally, mainly medial to the orbits, and extend caudally slightly beyond the caudal edge of the orbital rim. The frontals in this specimen project posteromedially between the parietals, allowing the parietals to project anterolaterally toward the orbital rim, resulting in reduced postorbitals. This also makes the parietals prominent, extending between the orbits beyond their posterior rim, distinctly farther than in
Hadrianus corsoni and
E. naomi. The carapace is complete, 142 mm long, with the normal complement of scutes and bones seen in Echmatemys [
2]. Five or six growth annuli are present on costal 1, suggesting this individual was at least five years old when it died [
4].
2.6. Echmatemys naomi
Referred specimen: A nearly complete skeleton of
Echmatemys naomi from the Green River Formation (FOBU 14014,
Figure 2). The identification is based on the lack of overlap of the nuchal by the pleurals, neural 1 being much longer than is it wide, the presence of the vertebral 4–5 sulcus over the last neural, and the wide suprapygal.
Description: FOBU 14014 has a carapace length of 110 mm. The first costal has four growth lines, suggesting this turtle's age at death was ~4 years [
4]. The carapace is similar to other
Echmatemys naomi specimens [
2]. The skull is more robust (22.7 mm long, 16 mm wide) than
E. haydeni, with more pronounced prefrontals. The frontals have a larger anteromedial process along the midline. The right prefrontal is dislocated, covering part of the anterior process of the frontals. The postorbital excludes the jugal from the dorsal margin of the orbit. The parietals form a posteriorly concave suture with the frontals, rather than anteriorly concave with a posteromedial projection of the frontals between them.
3. Discussion
Following the end-Cretaceous mass extinction, no terrestrial families of turtles survived in the northern continents [
6]. This left terrestrial turtle niches open, possibly until the Eocene. Basal testudinids have long been assumed to have an ecology similar to extant testudinids. This assumption is challenged by
Manouria emys, the most basal extant testudinid, which has a distinct ecology [
7], including a unique mode of food apprehension, breeding behavior, and a greater willingness to attempt subaqueous food capture. The histology of
Hadrianus is that of a semi-aquatic to aquatic turtle [
8]. It is near the base of a long stem leading to crown testudinids, so we need not assume they were similar to extant or even fossil crown testudinids in their ecology.
The oldest tortoises are stem-Testudinidae from the early Eocene of North America. The first record of these tortoises is
Hadrianus majusculus [
9] from the late Wasatchian (Wa-5) of North America (the oldest reported pantestudinid). Purported stem-Testudinidae and testudinids have been reported from the Eocene of Asia. A recent revision of
Anhuichelys [
10] concluded that it is a stem-testudinid, rather than an emydid, as previously proposed [
11]. However, the phylogenetic analysis [
10] is suspect, given that if either of the two characters are subtracted, the analysis finds a sister group relationship of
Anhuichelys with
Platysternon [
9]. This sister clade was found to be sister to both basal testudinoids included in the analysis (
Lindholemys and
Mongolemys). This agrees with the original placement [
11]. Purported stem-Testudinidae mentioned from Mongolia [
12] have never been published. These come from the "Naran Bulak Formation of Khaichin Ula IV and the Tsagan Khushu localities" (Igor Danilov pers. com., 2016). Tsagan Khushu is of late Paleocene to early Eocene age in the Naran Bulak Svita, whereas Khaichin Ula IV is of middle Eocene age in the Khaychin Svita [
13].
Perez-Garcia et al. [
14] revised Europe’s oldest tortoises, naming
Fontainichelon for the species “
Achilemys cassouleti” [
15]. They suggested the specimen is early Eocene (MP 8–9), roughly equivalent to the late Wasatchian NALMA. This is contrary to the original description, which placed it in MP 10, or approximately equivalent to the Bridgerian NALMA. There is some confusion between the sites in that region of France—the application of the age of the “La Borie” locality to these specimens, which are from a different locality (“Saint-Papoul”), is not justified [
16]. Thus,
Fontainichelon is younger than North American
Hadrianus.
Fontainichelon is also not the most basal testudinid, an assessment based on “the presence of narrower and longer gular scutes and a “wavy” humero–pectoral sulcus, with the medial part being "convex anteriorly” [
14].
Fontainichelon and
Hadrianus have similar-sized gular scutes, each occupying approximately one-third of the length of the midline. The “wavy” sulcus is present in a wide variety of taxa in the Emydidae, Geoemydidae, and Testudinidae, and encompasses a number of distinct morphologies. The closest comparison is
Rhinoclemmys pulcherima, a basal geoemydid. While this character is distinct in mature individuals, it is completely absent in a three-month-old hatchling of
R. pulcherima, which has a transverse humeral pectoral sulcus (which develops by 15 months).
Hutchison [
17] proposed that Testudinidae entered North America from Asia during the second thermal maximum of the Eocene (~52 Ma). Hutchison [
17] and Holroyd et al. [
18] identify three important events: (1)
Echmatemys sensu lato appearing in North America by Wa-0; (2)
Baptemys appearing, likely an immigrant from south of Texas, at Wa-5; and (3) at this same horizon in the Willwood Formation, Testudinidae make an abrupt first appearance. Instead, we conclude that
Echmatemys [
2] is a combination of at least three groups that entered North America at about the Paleocene-Eocene thermal maximum and emigrated from North America to Europe and Asia by the earliest Bridgerian. The oldest and most basal tortoises are from the western United States.
| Institutional Abbreviation | Location |
| FOBU | Fossil Butte National Monument, Kemmerer, Wyoming, USA |
| TMP | Royal Tyrrell Museum of Palaeontology, Drumheller, Alberta, Canada |
| UCB | University of Colorado Museum of Natural History, Boulder, Colorado, USA |



{kind=link}
{kind=link}