1. Introduction
Zinc (Zn) and manganese (Mn) are micronutrients that are inadequately available to plants worldwide [
1], and Zn deficiency is the most often in plants of all the micronutrients [
2]. In plants, Zn and Mn are transported predominantly as a divalent ion (Zn
2+, Mn
2+, respectively) or in organically bound [
3]. Unlike Fe and Cu, Zn is not involved in redox reactions in plants [
4]; instead, it plays a primary role in connection with various enzymes, for example, RNA polymerase, which is crucially involved in protein synthesis, contains two Zn atoms [
5], and carbonic anhydrase, which contains a Zn atom, is required in the carbon cycle of C4 plants to convert dissolved CO
2 to HCO
3− in the mesophyll cells, and allows diffusion of CO
2 to the RUBISCO in C3 plants [
4,
6]. Zn is directly involved in transcription because it is responsible for the finger-like structuring of polypeptide chains of some proteins. These “Zn fingers” enable the binding of DNA or RNA to proteins [
7]. In addition, Zn is central to the structure of ribosomes and also suppresses RNAse [
4]. The Zn-containing CuZnSOD protects plant cells from oxidative stress, which is common under abiotic stresses. The resulting H
2O
2 is subsequently degraded to H
2O by catalases, the function being dependent on an adequate Zn supply [
4].
The transport of Zn and Mn in the soil to the plant roots is mainly by diffusion from the rhizosphere [
8], and their uptake, predominantly as divalent cations (Mn
2+ and Zn
2+) in acidic environments and MnOH
+/ZnOH
+ under alkaline conditions [
4], occurs via ion channels. In addition, the uptake of their sulfate salts and chelates is also possible [
9]. The availability and dynamics of micronutrients in soil depend on several factors such as the total contents and characteristics of soils. The total amount of Zn or Mn in soils is generally composed of five fractions; however, only exchangeable and dissolved fractions are directly available for plant uptake. The exchangeable fraction is reversibly adsorbed to organic or oxidic soil particles and is in exchange with the dissolved fraction [
10].
A decisive factor for the solubility and thus the availability of Zn and Mn to plants is the soil pH: the slightly acidic to acidic range is optimal for the availability and uptake of most micronutrients [
11]. Soil pH not only directly affects the form of the metals but also the CEC of the soil. Thus, Zn and Mn availabilities are also indirectly affected by pH: high pH values cause lower occupancies of the exchangers, which increase the adsorption (reversible) of these metallic ions onto soil particles [
12]. The fixation of the metals to clay minerals and carbonates is also higher with an increase in pH [
13]. Carbonate-rich or alkaline soils, such as those used for agriculture in arid regions of the world, are, therefore, particularly affected by the deficiency of these nutrients [
1].
Soil pH, in turn, is governed by various factors. During pedogenesis, from carbonate-bearing rocks receiving little precipitation, neutral to alkaline soils with high CEC values tend to form. Silicate-rich parent rocks receiving high precipitation, on the other hand, create acidic soils [
12]. Plants also influence the pH of the soil near the roots. For example, depending on plant species, nutrient supply, and other factors, the difference in pH between the rhizosphere and the soil independent of plants can be as much as two pH units [
13].
In addition to these natural factors, agronomic practices affect soil pH: the liming of agricultural acidic soils increases soil pH, whereas the application of acidic mineral fertilizers containing SO
42− can lead to a lowering of the soil pH [
3]. The application of nitrogenous (N) fertilizers, depending upon whether the N form contained in the fertilizer is ammonium (NH
4+) or nitrate (NO
3−), has also been reported to have physiological and/or chemical effects on soil pH, and, in turn, on the availability of Mn [
14]. During NH
4+ uptake, plant roots release protons (H
+) to maintain a constant intracellular charge balance. This process leads to a decrease in rhizospheric pH [
15], which is also called physiological acidification. Fertilization with NH
4+ also has a chemical acidifying effect beyond the rhizosphere through the process of nitrification, which is independent of plant roots [
16]. Protons are released in this process, which acidify the soil [
17].
Nitrate uptake has the effect opposite to NH
4+ nutrition, whereby plants absorb H
+ by co-transportation with nitrate across the plasma membrane of root cells, resulting in higher pH in the root apoplast and rhizosphere [
18]. In addition, when NO
3− is taken up, OH
− is released, which also raises the soil pH in the rhizosphere [
14]. The application of ammonium nitrate (NH
4NO
3) has a “mixed” effect on pH; however, NH
4+ is preferentially taken up in the rhizosphere at equal concentrations of the two N-forms [
19], leading to a decrease in pH, despite NO
3− supply. Although NH
4+ fertilization is known to acidify rhizosphere and enhance nutrient uptake, these effects on a nutrient-sufficient acidic soil amended with lime have not been demonstrated. To investigate the interactions between the form of applied N fertilizer and a liming treatment of neutral soil, a pot experiment was conducted on maize (
Zea mays L.). It was hypothesized that NH
4+ uptake would lead to a lowering of soil pH in both limed and unlimed soil, which, in turn, would increase the solubility, availability, and uptake of micronutrients such as Mn and Zn. Furthermore, as limed soil would have stronger pH buffering, the effect of NH
4+ fertilization on the uptake of Mn or Zn would be weaker in the limed soil than in unlimed soil.
2. Materials and Methods
2.1. Collection, Preparation and Characterization of Soil
The experiment was conducted in the greenhouse of the Institute of Plant Nutrition and Soil Science at Kiel University (54.35° N, 10.11° E), Germany under semi-controlled conditions. The mean air temperature in the greenhouse during the experimental period was 21 °C, and humidity was adjusted between 50–60% with an automatic humidifier. When the sunlight was intense, sunshades were automatically lowered. The lighting duration was 13 h during the day (7–20 h). The experimental soil was collected from an arable field in Ostholstein, Schleswig-Holstein, Germany. The soil was first air dried and then sieved to remove stones and plant residues. The soil was analyzed for pH and plant-available nutrients using the procedure described in
Section 2.4. As plant-available Zn in the original soil was 6.7 mg kg
−1 (
Table 1), which corresponded to a sufficient level for plant growth, no additional Zn was fertilized during the entire course of the experiment. The soil, approximately 120 kg, was fertilized using a soil mixer with the following fertilizers as a basal dose (mg per kg of soil): 348 KH
2PO
4, 743 K
2SO
4, 306 CaCl
2∙2H
2O, 563 MgSO
4∙7H
2O, and 19.8 Fe-EDTA, 6.5 CuSO
4∙5H
2O, 15 MnSO
4∙H
2O, 9.5 H
3BO
3, and 0.67 (NH
4)
6Mo
7O
24∙4 H
2O.
2.2. Treatment Application and Sowing of Maize
Half of the nutrient-added soil was kept unlimed (control), whereas the other half was limed. For this purpose, CaCO3 was added at the rate of 3 g kg−1 soil and mixed well. Adding this amount of CaCO3 raised the soil pH to about 7, which was originally 5.6. After liming and application of the basic nutrients, the unlimed soil (C) and limed soil (CC) was filled in pots (L 20 cm, diameter 20 cm) at the rate of 6 kg soil pot−1 (4.71 dm3) to achieve a bulk density of 1.27 kg dm−3. Four maize kernels (cv KWS Keops, KWS SAAT SE & Co. KGaA, Einbeck, Germany) were sown in each pot to a depth of about 2.5 cm. Keops is an early-maturing silage maize variety (S210) and is suitable for biogas production and cattle feeding. After sowing, both the soils were fertilized with 1g N kg−1 soil using aqueous solutions of either ammonium nitrate or ammonium sulfate fertilizers. This constituted, overall, four treatments (two N source under each of the soil), each having four independent pot replicates. Each pot was applied with 400 mL of respective N fertilizer solution, which moistened the soil to 60% of water holding capacity (WHC).
The soil was kept continuously moist to approximately 60–70% WHC using deionized water. Seven to ten days after sowing, all maize plants had emerged. Only two plants were retained in each pot for final growth. Two further fertilizations of phosphorus (P, 40 mg kg−1 soil) and potassium (K, 50 mg K kg−1) were carried out 35 and 50 days after sowing. Each re-fertilization corresponded to about half the rate of basal fertilization. Similarly, fertilization of Ca, Mg, S, and micronutrients, each equal to the basal fertilizer, was also carried out 50 days after sowing.
2.3. Harvesting, Sample Collection and Preparation
Maize plants were harvested 62 days after sowing. The shoot was cut off near the ground with a scalpel, coarsely chopped, placed in paper bags, and dried in an oven at 65 °C to a constant weight. The post-harvest soil was removed from the pots, and roots were separated as best as possible. The roots were washed and dried in an oven. The dry matters of roots and shoots were recorded on a weighing balance. The dried shoot samples were ground using a mill (Retsch SM300, Retsch GmbH, Haan, Germany) adjusted to 1200 rpm and fitted with 0.5 mm sieve. The root samples were cut into small pieces with scissors and then ground in a fine mill (Foss Cyclotec 1093, Foss GmbH, Hamburg, Germany). The ground root and shoot samples were further ground to fine powder in a vibrating mill (Retsch MM400, Retsch GmbH, Haan, Germany). The root to shoot ratio (WSV) was calculated.
After removing the roots from the pots, the adhering soil was separated from them by gently tapping against a hard surface. The collected soil samples were dried in air, sieved (0.63 mm), and stored in plastic bags for further analysis.
2.4. Chemical Analysis of the Soil
The pH of the original soil samples of both unlimed and limed soils were measured. For this purpose, 20 mL of deionized water was added to a 20 g soil sample in a 50 mL Falcon tube and shaken using an orbital motion shaker (Gerhardt RO5, C. Gerhardt & GmbH, Königswinter, Germany). The suspensions were centrifuged for 7 min at 2000 rpm (Heraeus Multifuge X3R, Thermo Fisher Scientific, Waltham, MA, USA). Subsequently, the pH of the solution was measured using a pH meter (WTW inoLab pH Level 1, Xylem Analytics GmbH, Ingolstadt, Germany).
The concentration of plant-available P in the soil was measured following the method of Olsen et al. [
20]. In brief, 2.5 g of each soil sample was extracted with 50 mL of 0.5
M NaHCO
3 solution (pH 8.5, adjusted with NaOH) for 30 min at 150 rpm for 30 min. The extracts were filtered (Whatman 5 filter paper), and P concentrations in them were measured using an autoanalyzer (SAN++ continuous flow analyzer, Skalar, Netherlands) pre-calibrated with a series of P standards (1–5 mg P L
−1).
The concentrations of micronutrients viz. Zn, Mn, Cu, and Fe in the original soil and the rhizosphere soil samples were determined according to Lindsay and Norvell [
21]. In brief, 15 g of each soil sample was extracted with 30 mL of 0.005
M DTPA solution (made in 1.0
M CaCl
2 and adjusted to pH 7.3 with 1:1 HCl) for 2 h using the above said orbital shaker at 120 rpm min
−1. The suspensions were centrifuged for 10 min at 2000 rpm and filtered using Whatman 42 filter paper. Subsequently, micronutrient concentrations were measured by atomic absorption spectrometer (SOLAAR S Series AA, Thermo Fisher Scientific, Waltham, MA, USA) (
Table 1).
2.5. Plant Analysis
To determine the nutrient concentrations (P, K, Mg, Ca, Fe, Cu, Mn, and Zn) in the plant tissues, the samples were digested in nitric acid (HNO3). Approximately 200 mg of each plant sample was weighed in the digestion tube, and 10 mL of HNO3 (69% Suprapur®) was added. The tubes were then placed in the microwave (MARS 6 Xpress, CEM Corporation, Matthews, NC, USA), where digestion was performed (“Classic” program: 20 min at 190 °C). The digests were cooled to room temperature and diluted to 100 mL with ultrapure water. The obtained aliquots were further diluted 20 times with ultrapure water, and measurements of nutrient concentrations were performed by an inductively coupled plasma mass spectrometer (ICP-MS; Agilent 7700, Agilent Technologies Inc., Santa Clara, CA, USA). The nutrient contents in roots and shoots were calculated by multiplying the concentrations (mg g−1) with dry matters (g pot−1).
2.6. Statistical Analysis
The experimental pots were arranged in a completely randomized design. The analysis of variance (ANOVA) was performed following a two-factor factorial design, whereas the significant differences between the individual treatments were determined by Tukey’s Honestly Significant Difference test (p ≥ 0.5). For this purpose, the programming language “R” (R 4.1.1) was used with the user interface “RStudio” (2021.09.0, © 2009–2021 RStudio, PBC) and the function “gls” therein.
4. Discussion
The purpose of this experiment was to investigate the influence of NH4NO3 and NH4+ forms on the uptake of various nutrients, especially the micronutrients Mn and Zn, and maize growth, with particular reference to liming of an acidic soil. Chemical, biological, and physical conditions in the rhizosphere can differ greatly from the rest of the soil due to the exudation of organic compounds, ions, or water through the roots. The pH of the rhizosphere soil is among the most significantly affected soil properties. Different plant species alter pH differently, and the initial soil pH also determines whether the rhizosphere pH is raised or lowered by root influence.
The application of lime (CaCO
3) by itself has the opposite effects to that of soil NH
4+ fertilization. In this experiment, liming was carried out to raise the soil pH to simulate the widespread alkaline Zn deficient soils, which are common in intensive arable farming system of semiarid and arid regions. After the application of lime, the protons present in the soil are buffered until saturation, i.e., until all lime is used up [
12,
22]. The resultant increase in pH either has a positive or negative effect on plant growth, depending upon the initial soil pH and the plant species. An excessive increase in pH can have negative effects on plant growth. In particular, P dynamics along with the availability and uptake of micronutrients have an optimal soil pH range that depends on the plant species.
4.1. Plant Growth and Dry Matter Production
In general, mixed fertilization with NH
4NO
3 is considered optimal for many crop species such as wheat, as this allows the benefits of both N forms to be exploited under practical conditions [
23]. Pedersen et al. [
24] showed that a mixed application of NH
4+ and NO
3− produced the highest DM yields, whereas pure NH
4+ fertilization resulted in the reduced growth of young maize. However, at neutral to slightly acid soil pH values, plant growth was improved by NH
4+ compared to NH
4NO
3 [
25]. This was consistent with the observations from this study (
Figure 1). It is possible that some of the NH
4+ here was nitrified during the experimental period, so that the plants ultimately took up NO
3−, and the NH
4+ concentration remained below the phytotoxic level. In the present study, most of the observed pH effects of the NH
4+ fertilization could have been due to nitrification-induced acidification. However, acidification in the rhizosphere soil is further stimulated by the ammonium uptake of maize.
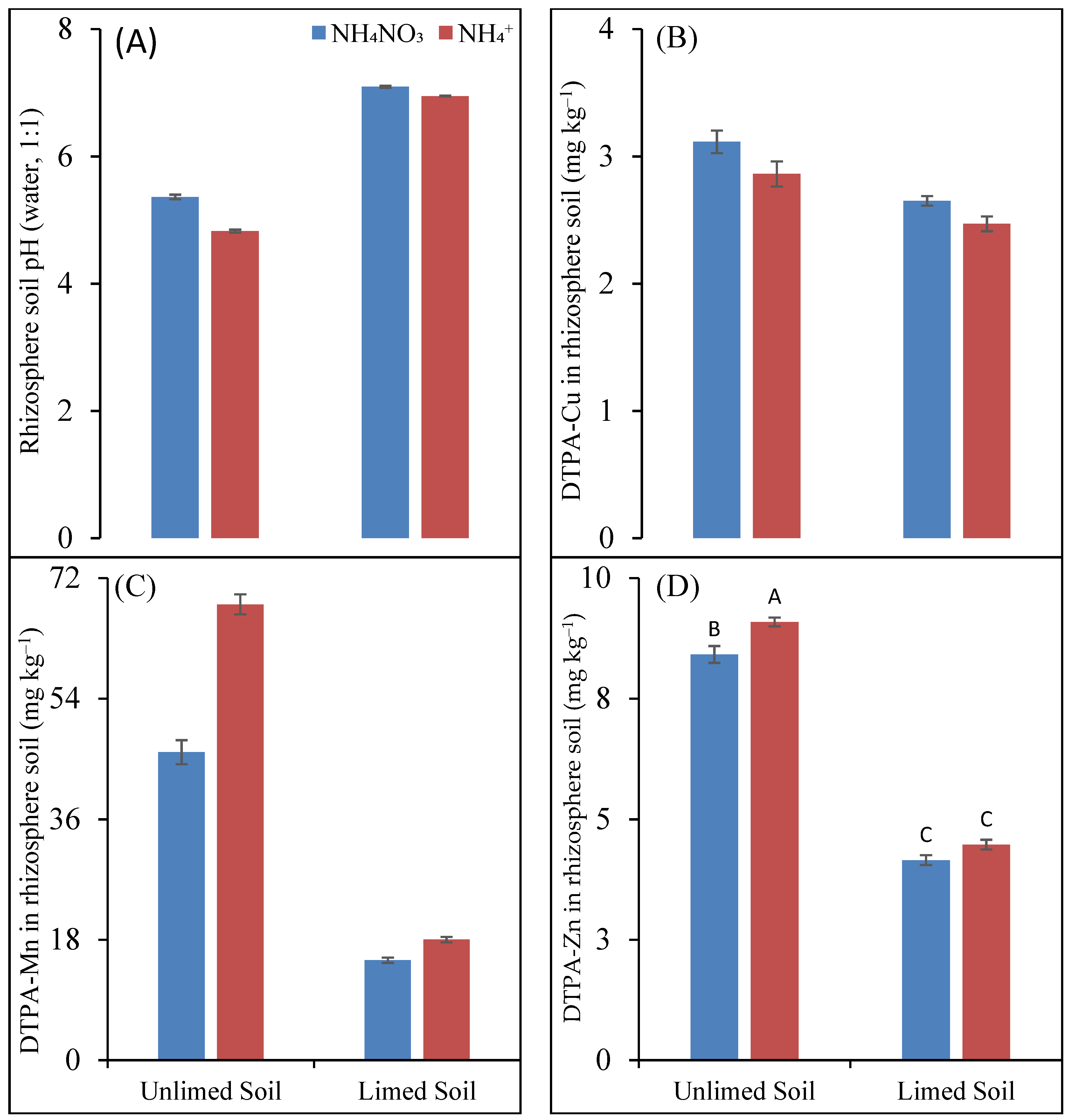
4.2. The Uptake of NH4+ Leads to a Lowering of the Soil pH in Limed and Unlimed Soil
The expected lowering of pH by NH
4+ compared to NH
4NO
3 fertilization was observed in this study (
Figure 2A). The mechanism of the uptake of NH
4+ by plants having an acidifying effect on rhizosphere soil has already been shown by other researchers. For example, Marschner and Römheld [
15] observed a strong acidification of the rhizosphere for various plant species when NH
4+ was in the form of N applied. Acidification of rhizosphere soil by NH
4+ fertilization is associated with both the release of H
+ by plant roots, due to the stimulation of H
+-ATPase, to counterbalance the charge resulting from the NH
4+ absorption by the roots and the nitrification of the NH
4+ itself [
16,
26]. However, the magnitude of the effect is very high in the former case, which is further enhanced by the addition of nitrification inhibitors [
16]. Consistent with this, Taylor and Bloom [
27] observed a significant decrease in pH on the rhizosphere of maize plants when NH
4+ was the only N source.
The lowering of the pH due to the NH
4+ fertilization was recorded in both the limed and unlimed rhizosphere soils; however, the unlimed rhizosphere soil experienced a higher degree of decline (0.54 units) than the limed soil (0.15 units) (
Figure 2A). Only a slight decline in the pH of the limed rhizosphere soil was explained by the buffering effect of the lime, which greatly attenuated the NH
4+-induced acidification by neutralizing the H
+ [
12]. However, it is worth considering that the effect of NH
4+ fertilization was significant despite the pH buffering; thus, the rhizosphere pH of the NH
4+-fertilized soil was lower than the NH
4NO
3-fertilized soil.
The combined fertilization of the NH
4+ and NO
3− as NH
4NO
3 resulted in only a slight lowering of the rhizosphere soil pH compared to the pH of the original soil. The small effect of the NH
4NO
3 fertilization on the pH was explained by the offsetting of the acidifying effect of the ammonium by the H
+/NO
3– cotransport that alkalized the rhizosphere thereafter [
23]. Even a low concentration of NO
3− could partially offset the negative effects of the NH
4+ fertilization [
28]. However, due to the lower energy required for uptake and assimilation, plants preferentially took up NH
4+ when both N forms were present in equal concentrations in the rhizosphere [
23]. This explained the slightly lower rhizosphere pH of the NH
4NO
3-fertilized soil compared to original soil. When maize was fertilized only with NH
4+, it also led to a decrease in rhizospheric soil pH in direct proportion with the quantity of NH
4+ uptake [
24].
4.3. Influence of Liming and N-Form on Soil and Plant Macronutrients
Although soil phosphorus concentration was neither affected by N forms, nor was the difference between liming treatments significant, a significant difference in shoot P concentration was recorded within liming treatments as well as the N forms (
Table 2). This discrepancy may have been due to the test procedure used for analyzing soil P: extraction with NaHCO
3 according to Olsen et al. [
20] mainly detected the potentially available organic P compounds as well as the P bound to the Fe-Al oxides [
10]. These the main fixed forms at the soil pH values < 6.5 [
29]. However, the P fixed to Ca tends to be underestimated by the test method used, which may have been why inaccuracies occurred in the pH range tested here. Furthermore, a reliable statement about the P availability for the plants could only be derived in the case of poor P supply level, which, however, was not present here: the soil used was sufficient in plant-available P at the start of the experiment [
10] (
Table 1).
Thus, although soil P concentration was independent of liming, there was a negative, significant effect of liming on the plant P concentration and content (
Table 2). In addition, the plants with NH
4+ fertilization had significantly higher P contents in shoots than the NH
4NO
3-fertilized plants. In the shoots, there were interactions between the N-forms and liming levels, in that the effect was enhanced by liming (
Table 2). Riley and Barber [
30] showed that NH
4+ fertilization in soybeans (
Glycine max L.) resulted in lower rhizosphere pH levels, which ultimately increased P concentrations in the plants. These observations may be explained by the enhanced solubilization of Ca phosphates by the lowering of pH, which otherwise decreases as a result of liming and the associated higher pH and Ca activity [
10,
24].
Potassium concentrations and contents in maize shoots were significantly decreased by liming and increased by NH
4+ formation (
Table 2 and
Table 3); however, the decline in soil pH seemed to be the governing factor. At lower pH levels, H
+ had stronger competition with the exchangeable K
+ for the soil exchange colloid, thereby moving it into the soil solution and vice versa. Consequently, the NH
4+-uptake-induced acidification of the rhizosphere soil increased both K
+ concentrations and contents in maize shoots, whereas the reverse response was observed with liming. The NH
4+-uptake-induced lowering of the pH might have overwhelmed the frequently described competitive negative effect of the NH
4+ fertilization on the K
+ uptake [
19], and the lime-induced alkalization of the soil had acted the same against the enhancing effect of Ca
2+ on the K
+ uptake.
Liming increased Ca concentration and content in shoots, whereas NH
4+ decreased these attributes compared to NH
4NO
3 (
Table 2 and
Table 3). The increase in shoot Ca concentrations in the limed soil was well-expected, whereas the negative effect of the NH
4+ on the Ca
2+ uptake was very well known [
31]. According to Weil et al. [
32], NH
4+ limits the uptake and accumulation of cations such as Ca
2+. Similarly, Na et al. [
33] observed lower concentrations of Ca
2+ in the shoot with a higher NH
4+ application rate.
Mg concentration was higher with NH
4NO
3 compared to NH
4+ fertilizer, which could be justified by the competitive uptake between the cations Mg
2+ and NH
4+ [
10]. According to Senbayram et al. [
34], low pH levels can increase the availability of Mg in the soil, and its uptake is limited due to higher H
+ concentrations. In our study, shoot Mg content was significantly decreased by liming (
Table 3) because elevated concentrations of Ca due to liming may have also had an antagonistic effect on Mg
2+ uptake [
35].
4.4. NH4+ Uptake-Induced pH Lowering Increases Availability and Uptake of Micronutrients
The solubility, availability, and uptake of all micronutrients did not clearly depend on different N forms. Liming, on the other hand, affected all the studied micronutrients, as expected. The concentration of available Cu in the rhizosphere substrate was negatively affected by liming (
Figure 2B). On the other hand, despite a lowering of rhizosphere soil, NH
4+ fertilization tended to decrease DTPA-extractable Cu in rhizosphere soil. Thus, the two factors differing in their effect on soil pH had the same effect on available Cu in the soil. This was in line with the findings of Jahiruddin et al. [
36], who found no clear influence of soil pH on the DTPA extractability of Cu. On the other hand, however, consistent with this study, according to Fageria et al. [
37], Cu
2+ dissolved in soil is more strongly adsorbed onto clay minerals and organic matter at higher pH values. They explain this by the hydrolysis of hydrated Cu, which occurs at pH values above six. Thus, while availability was pH dependent, Cu uptake was, at the same time, hardly limited by the different pH-dependent Cu binding forms: many of these binding forms, e.g., organometallic complexes (Cu
org), can be taken up by plants [
10]. Therefore, Cu availability is generally assumed to be less pH dependent than is the case for Fe or Mn [
37]. Although availability of a nutrient in the soil is critical for plant uptake, the Cu concentrations in shoots showed a different pattern than the available concentrations in the soils: liming did not affect Cu concentrations in shoots, whereas it was higher when fertilization with NH
4NO
3 was carried out. The lower Cu concentration observed here in the NH
4+-fertilized plants could not be explained by a dilution effect since the Cu content between the NH
4NO
3- and NH
4+-fertilized plants also followed the same trend.
Although Fe is present in sufficient quantities in almost all soils, its availability is a problem on alkaline sites worldwide, mainly due to their high pH levels [
22]. Accordingly, in this experiment, available Fe concentration in soil was reduced from a mean value of about 100 mg kg
−1 soil without liming to 68 mg kg
−1 soil with liming. The Fe concentration in the maize shoot was influenced by liming in this trial, such as the concentration in the soil (
Table 5). However, this did not manifest itself as so-called lime chloroses, probably due to the reason that the plant-available Fe concentration in the limed soil was still enough to provide the required quantity of Fe to plants. Despite the difference in the rhizosphere pH between the N forms, similar to Cu, no positive effect of the NH
4+ fertilization on the concentration of the available Fe in soil and in shoots was observed in this experiment. This contradicted results from other studies, where NH
4+ feeding significantly increased Fe uptake and thus Fe concentrations in the shoot [
33,
38]. This discrepancy could have been explained by the fact that the NH
4+-induced change in the pH was not sufficient to solubilize the Fe in the soil, the concentration of which was already sufficient for plant growth.
Among the studied nutrients, the solubility and availability of Mn was most affected by N forms both in the unlimed and limed soil. As expected, soil Mn concentration was higher with NH
4+ fertilization than recorded with NH
4NO
3 fertilization (
Table 4 and
Table 5). The high pH dependence of Mn availability was also evident in the soil: NH
4+ fertilization caused a rhizosphere pH of below five, highly improved the available Mn concentration in the soil (
Figure 2C), and almost doubled shoot Mn concentration in both the unlimed and limed soils. The concentration of plant-available Mn
2+ was found to increase 100-fold when the soil pH decreased by one unit [
10]. The sharp increase in soil Mn
2+ concentration in an acidic soil (pH < 5.5) could be explained by the dissolution of MnO
2 [
39]. These observations were in agreement with that of Sabir et al. [
40] for maize. It was noteworthy that NH
4+ fertilization brought an additional, positive effect in terms of availability and uptake despite the very good supply of Mn in the soil in this experiment (
Table 1). According to Rengel [
41], this was due to the strong pH dependence of Mn, which has a stronger effect on the availability of this nutrient than the total content in the soil.
Zinc availability is also dependent on soil pH; therefore, a lower soil Zn concentration in the rhizosphere of the limed soil compared to the unlimed soil and a reverse effect were found to be true in the NH
4+- compared to the NH
4NO
3-fertilized unlimed soil only. Accordingly, the increased availability of Zn in the unlimed soil, in turn, reflected in the improved Zn concentrations and uptakes, leading to a conclusion that plant Zn concentration could also be increased using NH
4+ fertilization [
16,
39,
41]. According to Ma et al. [
38], for example, underfoot fertilization with NH
4+ increased shoot Zn concentration and yield of maize. On the one hand, they explained this with altered root growth. Thus, traits such as root length and density were increased due to NH
4+ application. On the other hand, they also observed an acidification of the rhizosphere and indicated this as the cause for the increased Zn concentration in the shoot. Acidification of the rhizosphere was also observed in this experiment. Interestingly, Zn availability was further increased due to pH reduction, even though the soil had an already sufficient concentration of plant-available Zn (
Table 1).








{kind=link}
{kind=link}