
Negligible Response of Transpiration to Late-Summer Nitrogen Fertilization in Japanese Oak (Quercus crispula)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
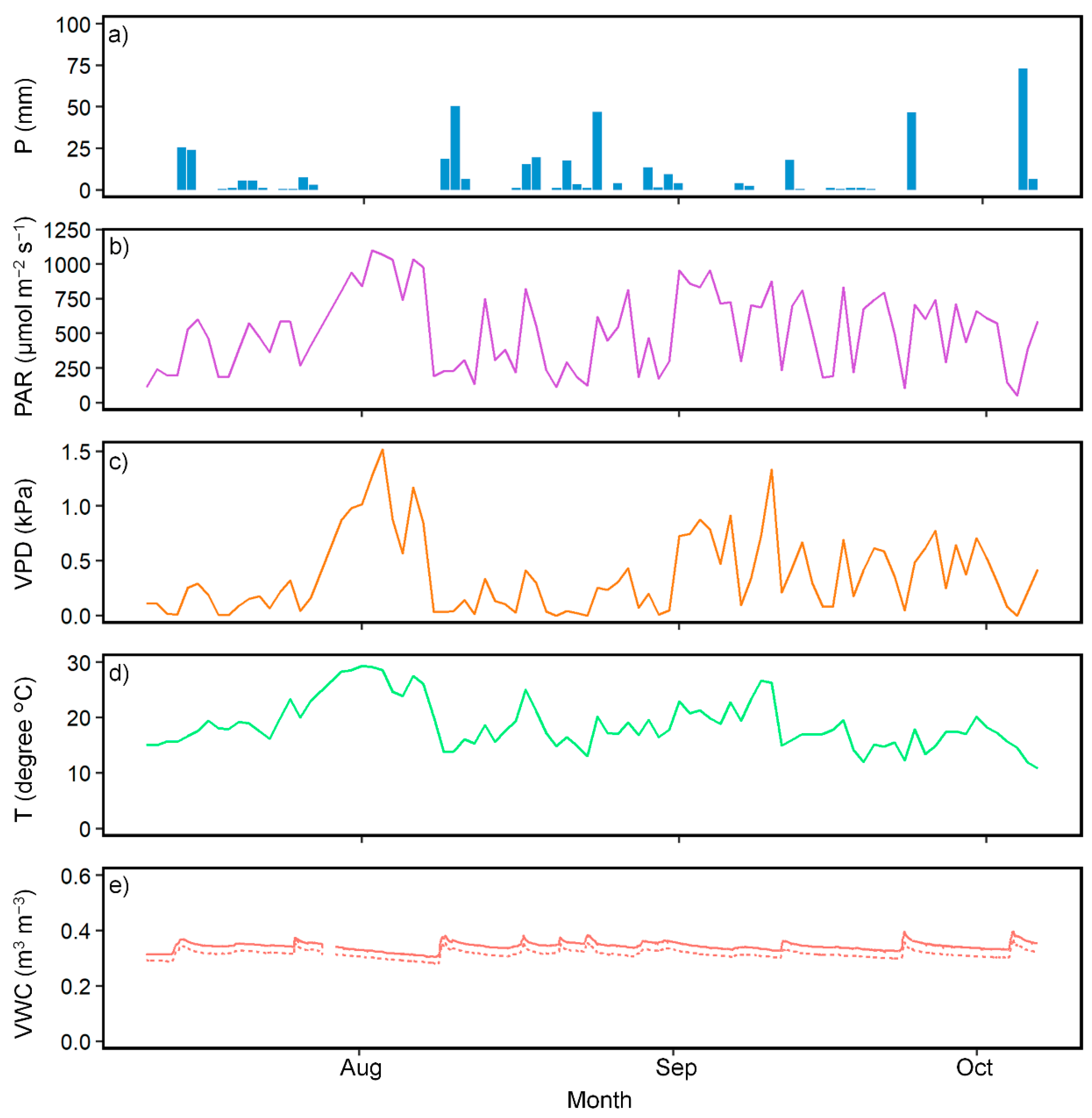
2.2. Meteorological and Soil Moisture Measurements
2.3. Sap Flow Measurements
2.4. N Fertilization
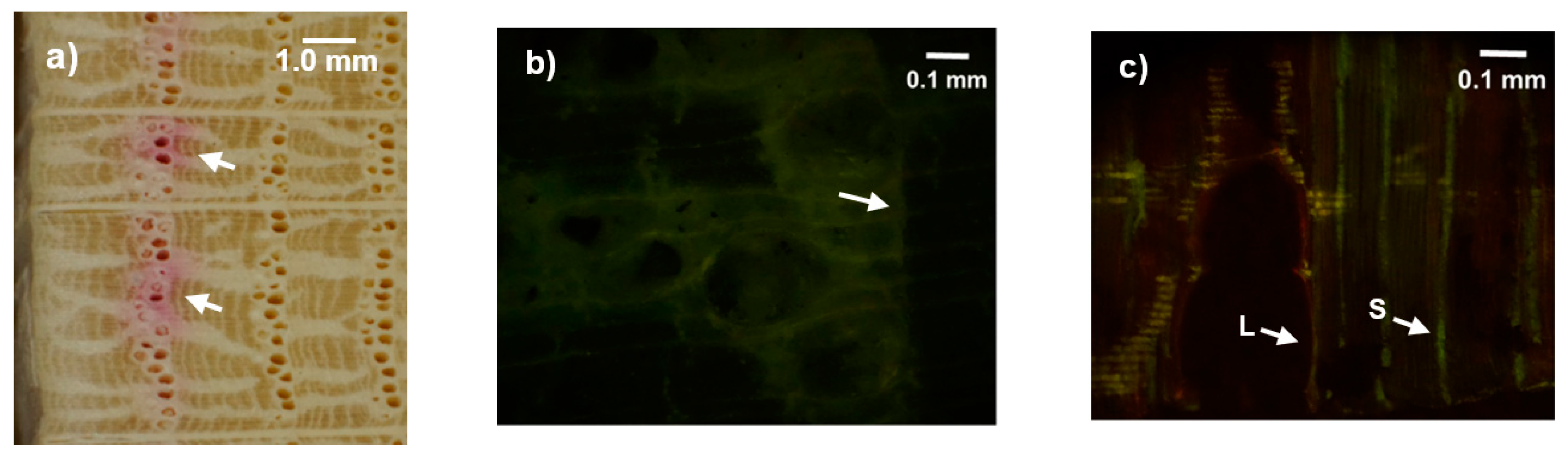
2.5. Dye Injection Experiments
2.6. Leaf N Contents
2.7. Calculations and Statistical Analysis
3. Results
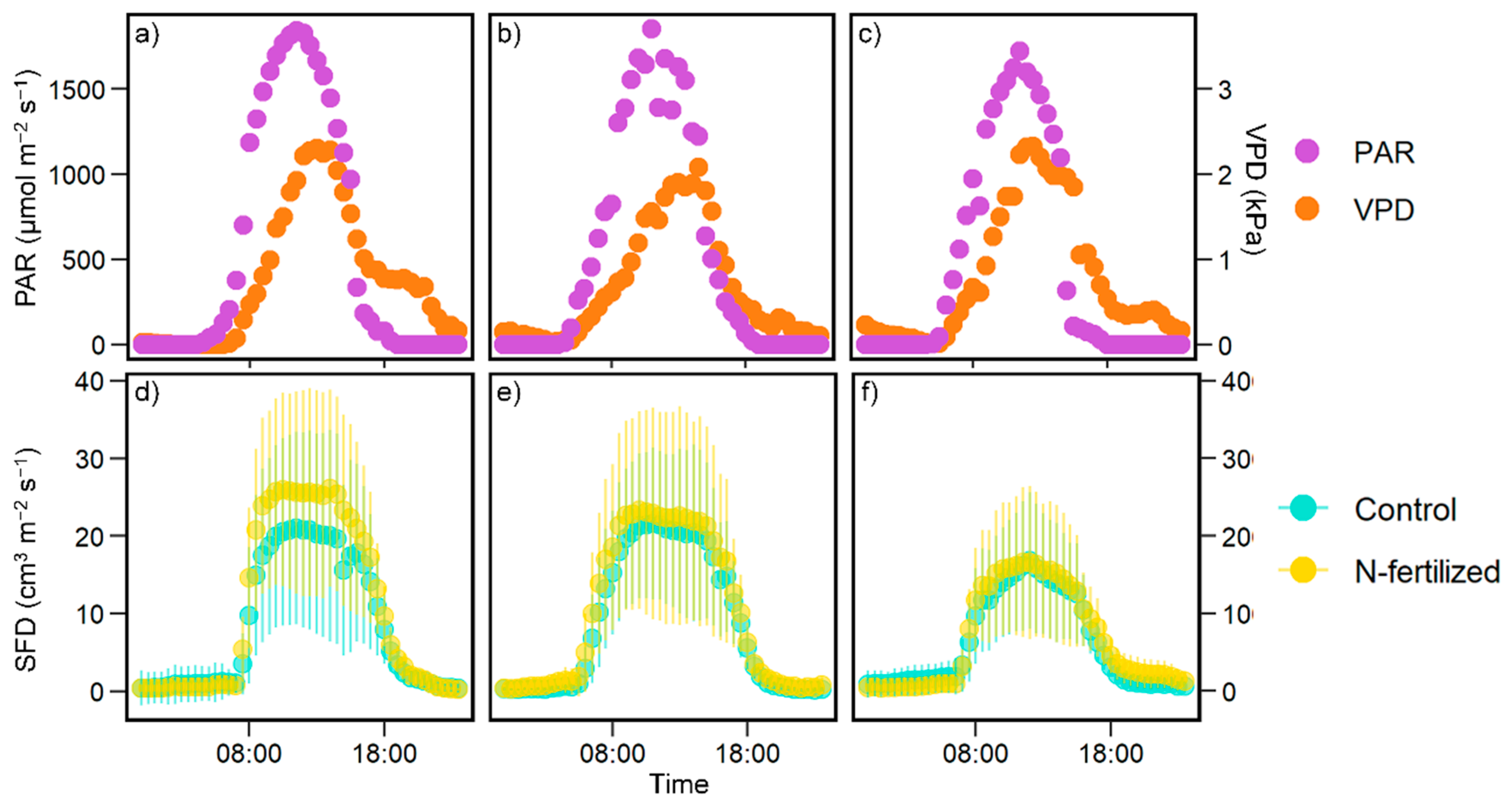
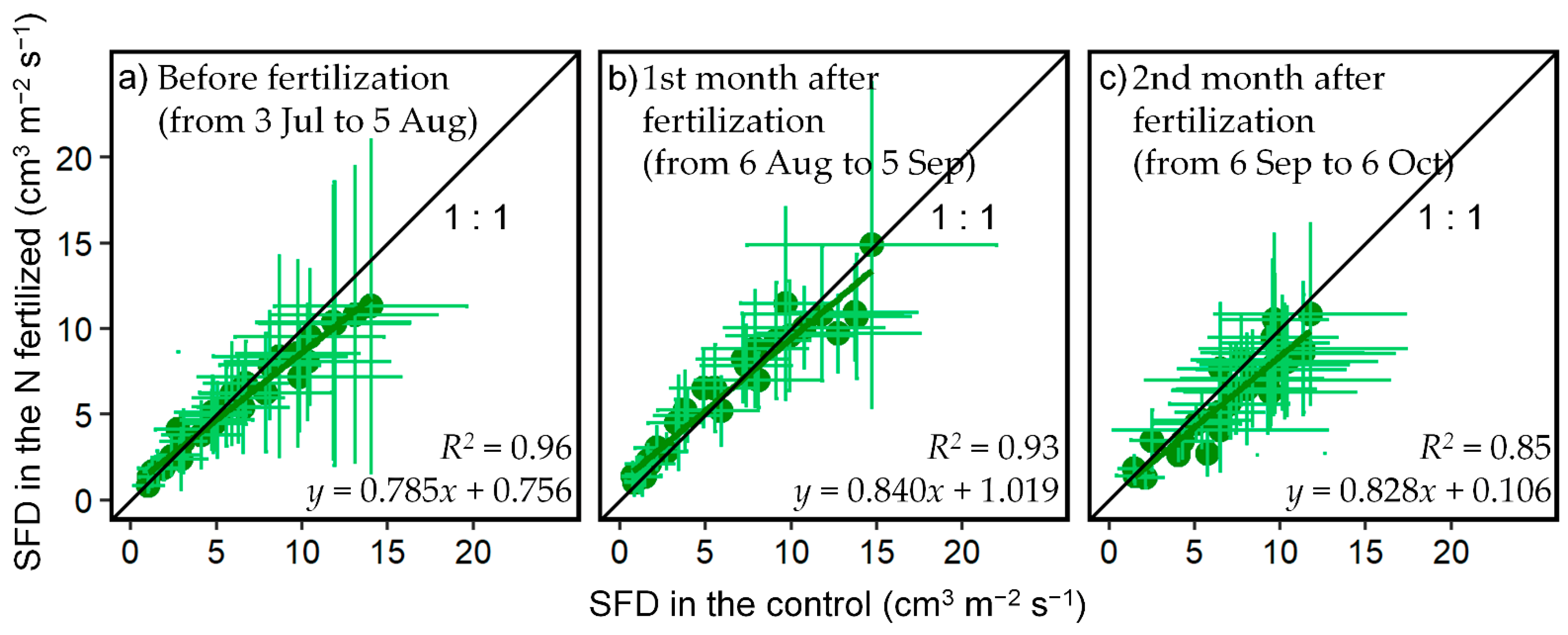
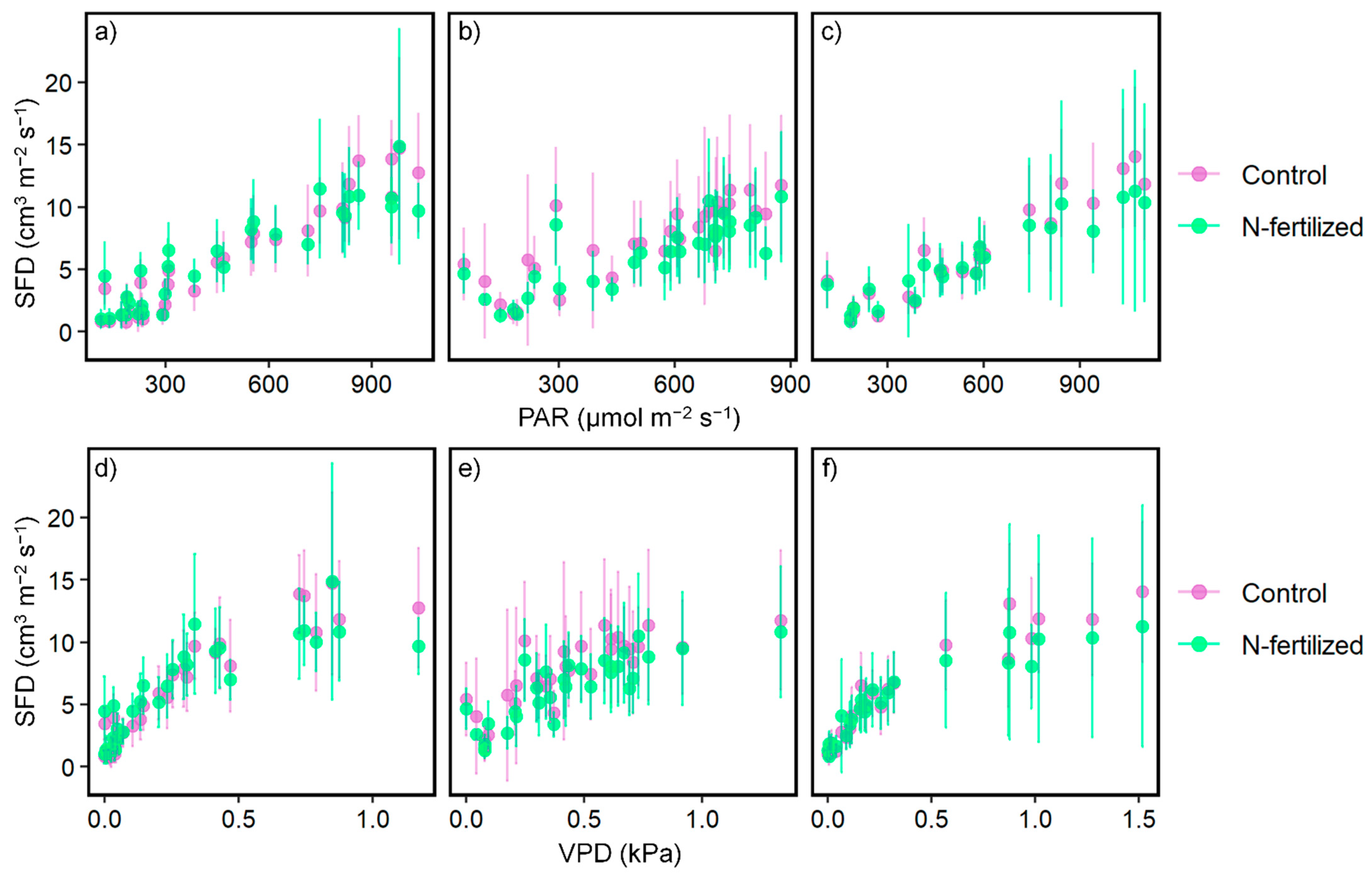
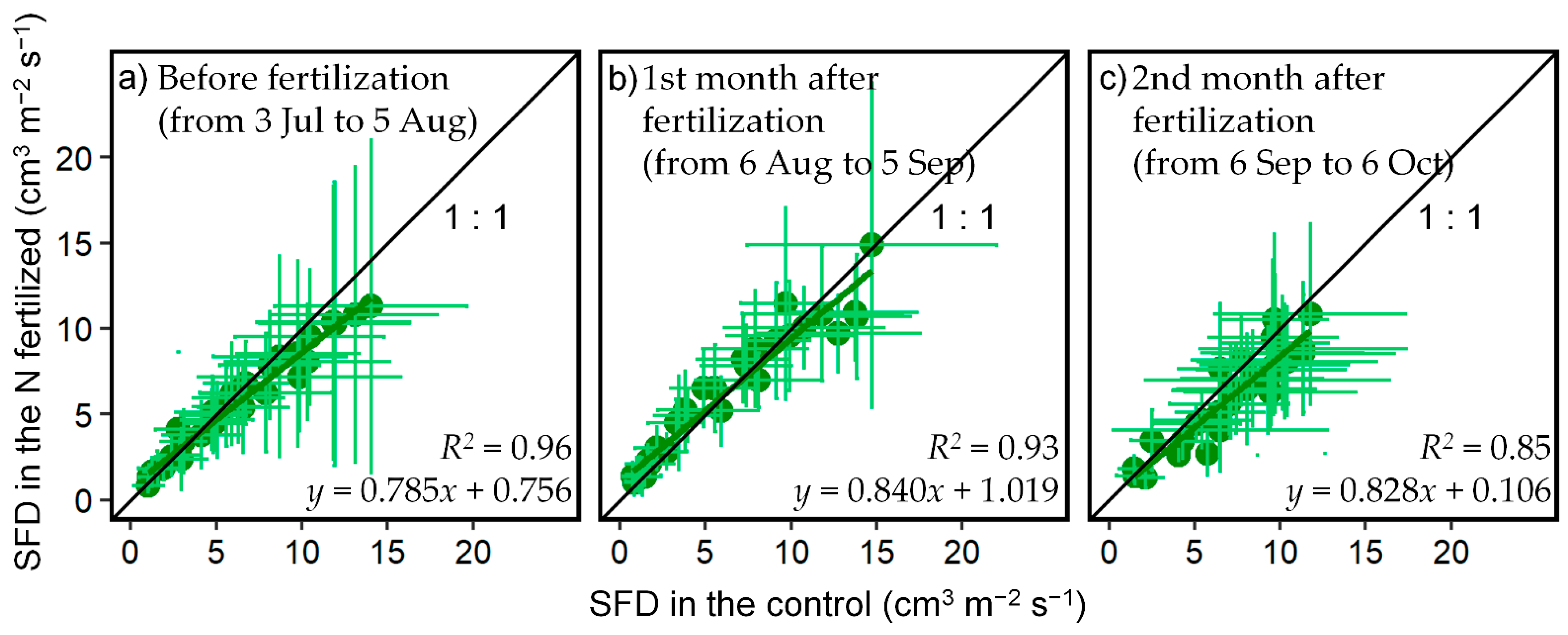
3.1. Effect of N Fertilization on SFD
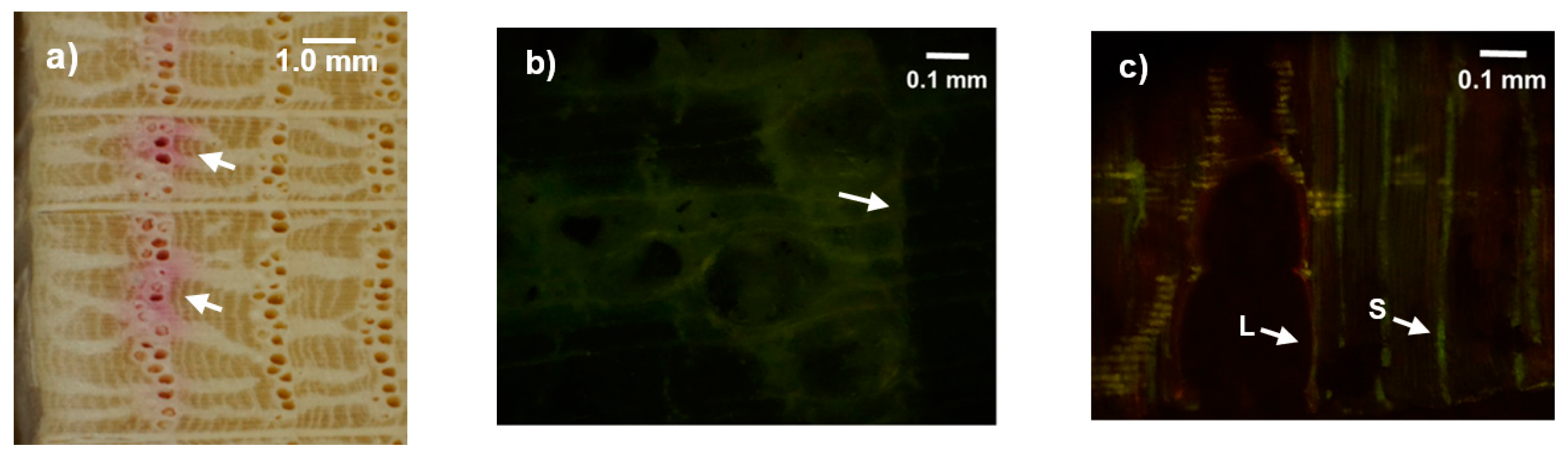
3.2. Dye Injection Experiments
3.3. Leaf N Contents
4. Discussion
4.1. Utility of Sap Flux Measurements in N Addition Experiments
4.2. Response of Tree Transpiration to Increased N Deposition
4.3. Response of Leaf N Contents to Increased N Deposition
4.4. Nonsignificance of the Short-Term Response
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ackerman, D.; Millet, D.B.; Chen, X. Global Estimates of Inorganic Nitrogen Deposition AcrossFour Decades. Glob. Biogeochem. Cycles 2018, 33, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Galloway, J.N.; Dentene, F.J.; Capone, D.G.; Boyer, E.W.; Howarth, R.W.; Seitzinger, S.P.; Asner, G.P.; Cleveland, C.C.; Green, P.A.; Holland, E.A.; et al. Nitrogen cycles: Past, present, and future. Biogeochemistry 2004, 70, 153–226. [Google Scholar] [CrossRef]
- Gao, Q.; Hasselquist, N.J.; Palmroth, S.; Zheng, Z.; You, W. Short-term response of soil respiration to nitrogen fertilization in a subtropical evergreen forest. Soil Biol. Biochem. 2014, 76, 297–300. [Google Scholar] [CrossRef]
- Aber, J.; McDowell, W.; Nadelhoffer, K.; Magill, A.; Berntson, G.; Kamakea, M.; McNulty, S.; Currie, W.; Rustad, L.; Fernandez, I. Nitrogen saturation in temperate forest ecosystem. hypotheses rivisited. Bioscience 1998, 48, 921–934. [Google Scholar] [CrossRef]
- Högberg, P.; Fan, H.; Quist, M.; Binkley, D.; Tamm, C. Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest. Glob. Chang. Biol. 2006, 12, 489–499. [Google Scholar] [CrossRef]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Zhang, T.; Lu, X.; Ellsworth, D.S.; BassiriRad, H.; You, C.; Wang, D.; He, P.; Deng, Q.; Liu, H.; et al. Global response patterns of plant photosynthesis to nitrogen addition: A meta-analysis. Glob. Chang. Biol. 2020, 26, 3585–3600. [Google Scholar] [CrossRef] [PubMed]
- Nagakura, J.; Kaneko, S.; Takahashi, M.; Tange, T. Nitrogen promotes water consumption in seedlings of Cryptomeria japonica but not in Chamaecyparis obtusa. For. Ecol. Manag. 2008, 255, 2533–2541. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, F.; Li, F. Effect of different drip irrigation methods and fertilization on growth, physiology and water use of young apple tree. Sci. Hortic. 2011, 129, 119–126. [Google Scholar] [CrossRef]
- Nilsen, P. Effect of nitrogen on drought strain and nutrient uptake in Norway spruce Picea abies (L.) Karst.) trees. Plant Soil 1995, 172, 73–85. [Google Scholar] [CrossRef]
- Welander, N.T.; Ottosson, B. The influence of low light, drought and fertilization on transpiration and growth in young seedlings of Quercus robur L. For. Ecol. Manag. 2000, 127, 139–151. [Google Scholar] [CrossRef]
- Harvey, H.P.; Van Den Driessche, R. Nitrogen and potassium effects on xylem cavitation and water-use efficiency in poplars. Tree Physiol. 1999, 19, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhao, P.; Zhu, L.; Zhao, X.; Ni, G.; Ouyang, L.; Schäfer, K.V.R.; Shen, W. Responses of sap flux and intrinsic water use efficiency to canopy and understory nitrogen addition in a temperate broadleaved deciduous forest. Sci. Total Environ. 2019, 648, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Forrester, D.I.; Collopy, J.J.; Beadle, C.L.; Warren, C.R.; Baker, T.G. Effect of thinning, pruning and nitrogen fertiliser application on transpiration, photosynthesis and water-use efficiency in a young Eucalyptus nitens plantation. For. Ecol. Manag. 2012, 266, 286–300. [Google Scholar] [CrossRef]
- Ewers, B.E.; Oren, R.; Albaugh, T.J.; Dougherty, P.M. Carry-over effects of water and nutrient supply on water use of pinus taeda. Ecol. Appl. 1999, 9, 513–525. [Google Scholar] [CrossRef]
- Hubbard, R.M.; Ryan, M.G.; Giardina, C.P.; Barnard, H. The effect of fertilization on sap flux and canopy conductance in a Eucalyptus saligna experimental forest. Glob. Chang. Biol. 2004, 10, 427–436. [Google Scholar] [CrossRef]
- Lu, X.; Liu, Z.; An, S.; Miralls, D.G.; Maes, W.; Liu, Y.; Tang, J. Potential of solar-induced chlorophyll fluorescence to estimate transpiration in a temperate forest. Agric. For. Meteorol. 2018, 252, 75–87. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, F.; Gou, X.; Fonti, P.; Xia, J.; Gao, Z.; Liu, J.; Wang, Y.; Zhang, J. Seasonal variations in leaf-level photosynthesis and water use efficiency of three isohydric to anisohydric conifers on the Tibetan Plateau. Agric. For. Meteorol. 2021, 308–309, 108581. [Google Scholar] [CrossRef]
- Chiwa, M. Long-term changes in atmospheric nitrogen deposition and stream water nitrate leaching from forested watersheds in western Japan. Environ. Pollut. 2021, 287, 117634. [Google Scholar] [CrossRef] [PubMed]
- Japan Meteorological Agency. Available online: http://www.jma.go.jp/jma/ (accessed on 13 August 2021).
- Imada, M. Study on the high forest system of Mizunara (Quercus crispula Blume). Bull. Kyushu Univ. For. 1972, 45, 81–225. [Google Scholar]
- Chiwa, M.; Nakamura, T. Temporal variation of air temperature, snow depth, and soil temperature at different slopes of Ashoro Research Forest, Kyushu University. Bull. Kyushu Univ. For. 2020, 101, 7–11, (In Japanese with English Summary). [Google Scholar]
- Frazer, G.W.; Canham, C.D.; Lertzman, K.P. Gap light analyzer (GLA), Version 2.0: Image-processing software to analyze true-color, hemispherical canopy photographs. Bull. Ecol. Soc. Am. 2000, 81, 191–197. [Google Scholar]
- Buck, A.L. New Equations for Computing Vapor Pressure and Enhancement Factor. J. Appl. Meteorol. Climatol. 1981, 20, 1527–1532. [Google Scholar] [CrossRef] [Green Version]
- Granier, A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiol. 1987, 3, 309–320. [Google Scholar] [CrossRef] [PubMed]
- James, S.A.; Clearwater, M.J.; Meinzer, F.C.; Goldstein, G. Heat dissipation sensors of variable length for the measurement of sap flow in trees with deep sapwood. Tree Physiol. 2002, 22, 277–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, P.; Urban, L.; Ping, Z. Granier’s Thermal Dissipation Probe (TDP) Method for Measuring Sap Flow in Trees: Theory and Practice. Acta Bot. Sin. 2004, 46, 631–646. [Google Scholar]
- Shen, Y.; Fukatsu, E.; Muraoka, H.; Saitoh, T.M.; Hirano, Y.; Yasue, K. Climate responses of ring widths and radial growth phenology of Betula ermanii, Fagus crenata and Quercus crispula in a cool temperate forest in central Japan. Trees 2020, 34, 679–692. [Google Scholar] [CrossRef]
- Song, L.; Kuang, F.; Skiba, U.; Zhu, B.; Liu, X.; Levy, P.; Dore, A.; Fowler, D. Bulk deposition of organic and inorganic nitrogen in southwest China from 2008 to 2013. Environ. Pollut. 2017, 227, 157–166. [Google Scholar] [CrossRef]
- Wang, X.; Wu, Z.; Shao, M.; Fang, Y.; Zhang, L.; Chen, F.; Chan, P.W.; Fan, Q.; Wang, Q.; Zhu, S.; et al. Atmospheric nitrogen deposition to forest and estuary environments in the Pearl River Delta region, southern China. Tellus B 2013, 65, 20480. [Google Scholar] [CrossRef] [Green Version]
- Stevens, C.J.; Duprè, C.; Dorland, E.; Gaudnik, C.; Gowing, D.J.G.; Bleeker, A.; Diekmann, M.; Alard, D.; Bobbink, R.; Fowler, D.; et al. Nitrogen deposition threatens species richness of grasslands across Europe. Environ. Pollut. 2010, 158, 2940–2945. [Google Scholar] [CrossRef] [Green Version]
- Umebayashi, T.; Utsumi, Y.; Koga, S.; Inoue, S.; Shiiba, Y.; Arakawa, K.; Matsumura, J.; Oda, K. Optimal conditions for visualizing water-conducting pathways in a living tree by the dye injection method. Tree Physiol. 2007, 27, 993–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- RStudio Inc. Available online: https://rstudio.com/ (accessed on 13 August 2021).
- Ocheltree, T.W.; Nippert, J.B.; Prasad, P.V.V. Stomatal responses to changes in vapor pressure deficit reflect tissue-specific differences in hydraulic conductance. Plant Cell Environ. 2014, 37, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Kume, T.; Otsuki, K.; Du, S.; Yamanaka, N.; Wang, Y.-L.; Liu, G.-B. Spatial variation in sap flow velocity in semiarid region trees: Its impact on stand-scale transpiration estimates. Hydrol. Processes 2012, 26, 1161–1168. [Google Scholar] [CrossRef]
- Tseng, H.; Chiu, C.W.; Laplace, S.; Kume, T. Can we assume conservative temporal change in spatial variation of sap flux for year-round tree transpiration estimates? A case study in a Cryptomeria Japonica, central Taiwan. Trees 2017, 31, 1239–1251. [Google Scholar] [CrossRef]
- Tsuruta, K.; Kume, T.; Komatsu, H.; Higashi, N.; Umebayashi, T.; Kumagai, T.; Otsuki, K. Azimuthal variations of sap flux density within Japanese cypress xylem trunks and their effects on tree transpiration estimates. J. For. Res. 2010, 15, 398–403. [Google Scholar] [CrossRef]
- Umebayashi, T.; Utsumi, Y.; Koga, S.; Inoue, S.; Matsumura, J.; Oda, K.; Fujikawa, S.; Arakawa, K.; Otsuki, K. Xylem water-conducting patterns of 34 broadleaved evergreen trees in southern Japan. Trees 2010, 24, 571–583. [Google Scholar] [CrossRef]
- Umebayashi, T.; Fukuda, K. Seasonal changes in the occurrence of embolisms among broadleaved trees in a temperate region. Botany 2018, 96, 873–881. [Google Scholar] [CrossRef]
- Phillips, N.; Oren, R.; Zimmermann, R. Radial patterns of xylem sap flow in non-, diffuse- and ring-porous tree species. Plant Cell Environ. 1996, 19, 983–990. [Google Scholar] [CrossRef]
- Tateishi, M.; Kumagai, T.; Utsumi, Y.; Umebayashi, T.; Shiiba, Y.; Inoue, K.; Cho, K.; Otsuki, K. Spatial variations in xylem sap flux density in evergreen oak trees with radial-porous wood: Comparisons with anatomical observations. Trees 2008, 22, 23–30. [Google Scholar] [CrossRef]
- Komatsu, H.; Shinohara, Y.; Kume, T.; Tsuruta, K.; Otsuki, K. Does measuring azimuthal variations in sap flux lead to more reliable stand transpiration estimates? Hydrol. Process 2016, 30, 2129–2137. [Google Scholar] [CrossRef]
- Komatsu, H.; Kume, T.; Shinohara, Y. Optimal sap flux sensor allocation for stand transpiration estimates: A non-dimensional analysis. Ann. For. Sci. 2017, 74, 38. [Google Scholar] [CrossRef]
- Kume, T.; Tsuruta, K.; Komatsu, H.; Kumagai, T.; Higashi, N.; Shinohara, Y.; Otsuki, K. Effects of sample size on sap flux-based stand-scale transpiration estimates. Tree Physiol. 2010, 30, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kume, T.; Onozawa, Y.; Komatsu, H.; Tsuruta, K.; Shonohara, Y.; Umebayashi, T.; Otsuki, K. Stand-scale transpiration estimates in a Moso bamboo forest: (I) Applicability of sap flux measurements. For. Ecol. Manag. 2010, 260, 1287–1294. [Google Scholar] [CrossRef]
- Gobin, R.; Korboulewsky, N.; Dumas, Y.; Balandier, P. Transpiration of four common understorey plant species according to drought intensity in temperate forests. Ann. For. Sci. 2015, 72, 1053–1064. [Google Scholar] [CrossRef] [Green Version]
- Pietsch, S.A.; Hasenauer, H.; Kučera, J.; Čermák, J. Modeling effects of hydrological changes on the carbon and nitrogen balance of oak in floodplains. Tree Physiol. 2003, 23, 735–746. [Google Scholar] [CrossRef] [Green Version]
- Okano, T.; Nakai, T. Seasonal patterns of seedling growth and dry matter partitioning of deciduous broad-leaved trees in Hokkaido. Bull. Kyushu Univ. For. 1992, 65, 1–14, (In Japanese with English summary). [Google Scholar]
- Wirabuana, P.Y.A.P.; Sadono, R.; Juniarso, S. Fertilization Effects on Early Growth, Aboveground Biomass, Carbon Storage, and Leaf Characteristics of Eucalyptus pellita F. Muell. in South Sumatera. J. Manaj. Hutan Trop. 2019, 25, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Guehl, J.M.; Fort, C.; Fehri, A. Differential response of leaf conductance, carbon isotope discrimination and water-use efficiency to nitrogen deficiency in maritime pine and pedunculate oak plants. New Phytol. 1995, 131, 149–157. [Google Scholar] [CrossRef]
- Adachi, M.; Bekku, Y.S.; Konuma, A.; Kadir, W.R.; Okuda, T.; Koizumi, H. Required sample size for estimating soil respiration rates in large areas of two tropical forests and of two types of plantation in Malaysia. For. Ecol. Manag. 2005, 210, 455–459. [Google Scholar] [CrossRef]
- Hikosaka, K.; Nabeshima, E.; Hiura, T. Seasonal changes in the temperature response of photosynthesis in canopy leaves of Quercus crispula in a cool-temperate forest. Tree Physiol. 2007, 27, 1035–1041. [Google Scholar] [CrossRef]
- Ueda, M.U.; Mizumachi, E.; Tokuchi, N. Foliage nitrogen turnover: Differences among nitrogen absorbed at different times by Quercus serrata saplings. Ann. Bot. 2011, 108, 169–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, G.H.M.; Höckel, F.E. Long-term effects of ozone on Fagus sylvatica L.—An open-top chamber exposure study. Water Air Soil Pollut. 1995, 85, 1337–1342. [Google Scholar] [CrossRef]
- Lucas, P.W.; Diggle, P.J. The use of longitudinal data analysis to study the multi-seasonal growth responses of Norway and Sitka spruce to summer exposure to ozone: Implications for the determination of critical levels. New Phytol. 1997, 137, 315–323. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Density (SD, Stems ha−1) | Leaf Area Index (LAI, m2 m−2) | Diameter at Breast Height (DBH, cm) | |
|---|---|---|---|
| Control | 4188 | 3.95 | 13 ± 1.1 |
| N-Fertilized | 4250 | 4.02 | 12 ± 1.5 |
| p values | - | 0.62 | 0.37 |
| Tree Sample | T1 | T2 | T3 | T4 |
|---|---|---|---|---|
| DBH (cm) | 9.3 | 11.5 | 13.4 | 14.5 |
| Tree Height (m) | 10.8 | 10.5 | 12.2 | 12.3 |
| Before (from 3 July 2019 to 5 August 2019) | 1st Month after (from 6 August 2019 to 5 September 2019) | 2nd Month after (from 6 September 2019 to 6 October 2019) | p Value | |
|---|---|---|---|---|
| a 1 | 0.785 | 0.840 | 0.828 | >0.05 |
| b | 0.756 | 1.019 | 0.106 | >0.05 |
| R2 | 0.96 | 0.93 | 0.85 |
| Before (from 3 July 2019 to 5 August 2019) | 1st Month after (from 6 August 2019 to 5 September 2019) | 2nd Month after (from 6 September 2019 to 6 October 2019) | 1 p Value | |||
|---|---|---|---|---|---|---|
| PAR | Control | a | 0.011 | 0.014 | 0.0099 | ns |
| b | −0.25 | −0.94 | 2.18 | <0.05 | ||
| R2 | 0.79 | 0.93 | 0.67 | - | ||
| N-fertilized | a | 0.0091 | 0.012 | 0.0091 | ns | |
| b | 0.50 | 0.30 | 1.46 | <0.05 | ||
| R2 | 0.80 | 0.84 | 0.70 | - | ||
| p | ns | ns | ns | - | ||
| VPD | Control | a | 7.24 | 12.91 | 8.03 | <0.05 |
| b | 3.03 | 2.24 | 3.92 | ns | ||
| R2 | 0.69 | 0.86 | 0.65 | - | ||
| N-fertilized | a | 5.72 | 10.22 | 7.42 | <0.05 | |
| b | 3.20 | 3.08 | 3.02 | ns | ||
| R2 | 0.67 | 0.71 | 0.67 | - | ||
| 2 p value | p | 3 ns | ns | ns | - | |
| July | September | |||
|---|---|---|---|---|
| Control | N-Fertilized | Control | N-Fertilized | |
| Sample number | 10 | 10 | 10 | 10 |
| Mean value (%) | 2.25 | 2.17 | 2.12 | 2.20 |
| Standard deviation (%) | 0.18 | 0.17 | 0.18 | 0.21 |
| Required sample size (±10%) | 3 | 3 | 4 | 5 |
| Required sample size (±20%) | 1 | 1 | 1 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagano, N.; Kume, T.; Utsumi, Y.; Tashiro, N.; Otsuki, K.; Chiwa, M. Negligible Response of Transpiration to Late-Summer Nitrogen Fertilization in Japanese Oak (Quercus crispula). Nitrogen 2022, 3, 76-89. https://doi.org/10.3390/nitrogen3010006
Nagano N, Kume T, Utsumi Y, Tashiro N, Otsuki K, Chiwa M. Negligible Response of Transpiration to Late-Summer Nitrogen Fertilization in Japanese Oak (Quercus crispula). Nitrogen. 2022; 3(1):76-89. https://doi.org/10.3390/nitrogen3010006
Chicago/Turabian StyleNagano, Nao, Tomonori Kume, Yasuhiro Utsumi, Naoaki Tashiro, Kyoichi Otsuki, and Masaaki Chiwa. 2022. "Negligible Response of Transpiration to Late-Summer Nitrogen Fertilization in Japanese Oak (Quercus crispula)" Nitrogen 3, no. 1: 76-89. https://doi.org/10.3390/nitrogen3010006
APA StyleNagano, N., Kume, T., Utsumi, Y., Tashiro, N., Otsuki, K., & Chiwa, M. (2022). Negligible Response of Transpiration to Late-Summer Nitrogen Fertilization in Japanese Oak (Quercus crispula). Nitrogen, 3(1), 76-89. https://doi.org/10.3390/nitrogen3010006