Nitrogen Cycling and Mass Balance in the World’s Mangrove Forests


Abstract
1. Introduction
2. Nitrogen Concentrations and Standing Stocks
2.1. Dissolved N Concentrations in Tidal Waters and Porewater
2.2. Total N Concentrations and C/N Ratios in Forest Components and Soil
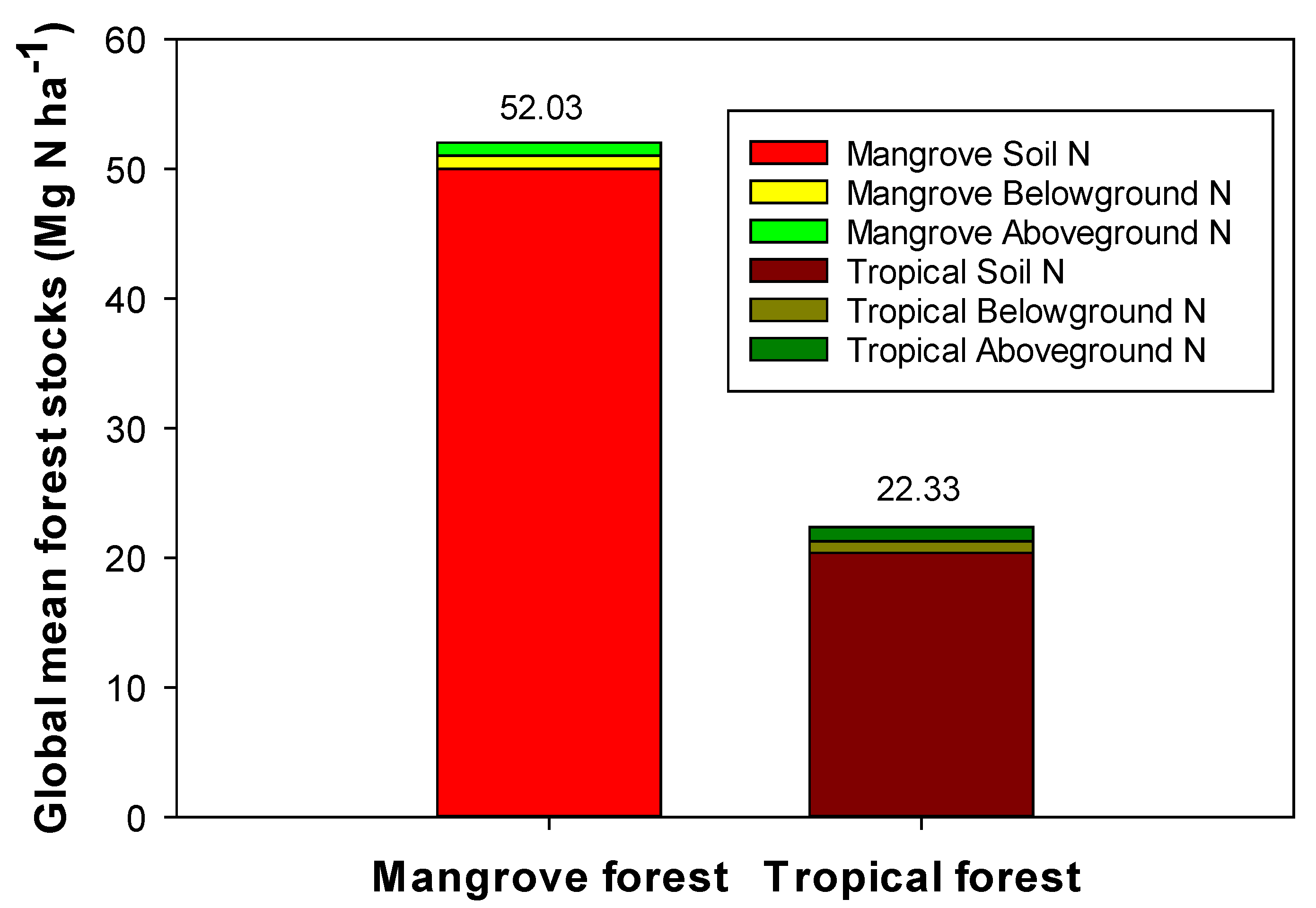
2.3. Global Mean N Stocks
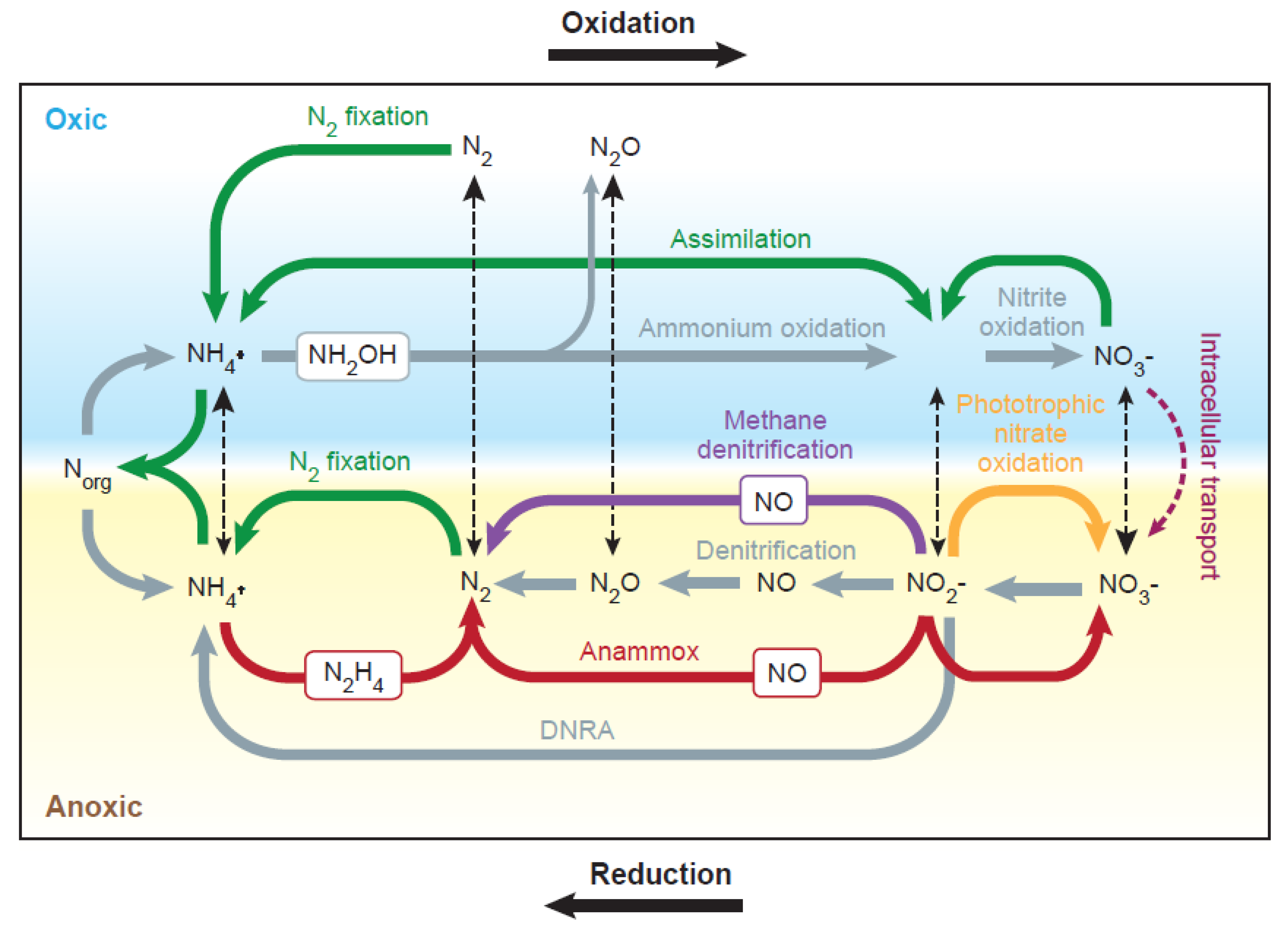
3. The Nitrogen Cycle in Soils
3.1. Forest Soil N Transformations
3.1.1. Nitrogen Fixation
3.1.2. Within-Soil Transformations
3.1.3. NO, N2O and DN Fluxes from Surface Soils
4. Tidal Water N Processes
4.1. N2O Fluxes
4.2. Tidal Exchange
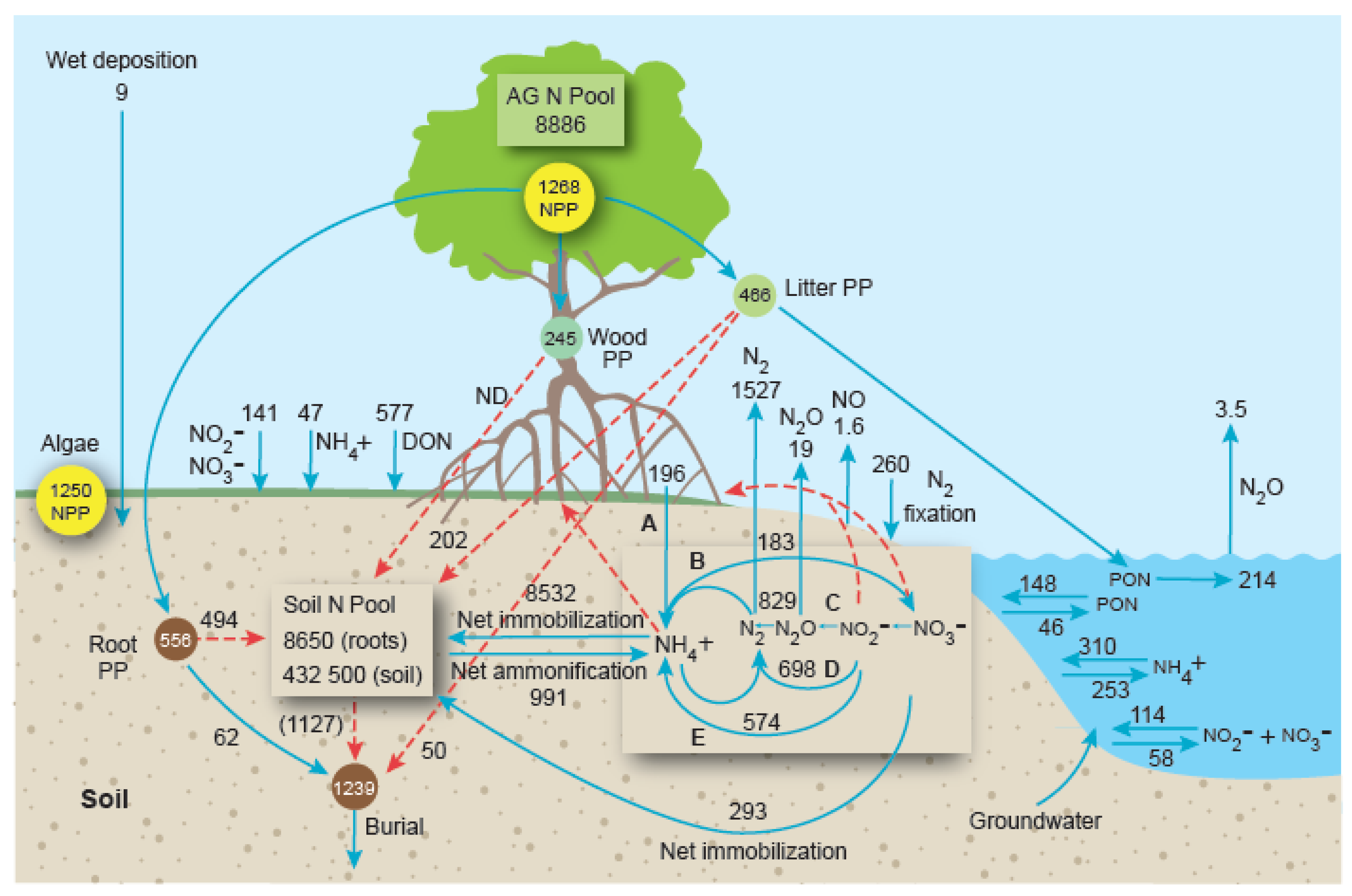
5. An Ecosystem-Level View of Mangrove Forests: A N Budget
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Alongi, D.M. The Energetics of Mangrove Forests; Springer Science: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Feller, I.C.; Lovelock, C.E.; Berger, U.; McKee, K.L.; Joye, S.B.; Ball, M.C. Biocomplexity in mangrove ecosystems. Annu. Rev. Mar. Sci. 2010, 2, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Alongi, D.M. Cycling and global fluxes of nitrogen in mangroves. Glob. Environ. Res. 2013, 17, 173–182. [Google Scholar]
- Alongi, D.M.; Boto, K.G.; Robertson, A.I. Nitrogen and Phosphorus Cycles. In Tropical Mangrove Ecosystems; Robertson, A.I., Alongi, D.M., Eds.; American Geophysical Union: Washington, DC, USA, 1992; pp. 251–292. [Google Scholar]
- Reef, R.; Feller, I.C.; Lovelock, C.E. Nutrition of mangroves. Tree Physiol. 2010, 30, 1148–1160. [Google Scholar] [CrossRef]
- Boto, K.G. Nutrients and mangroves. In Pollution in Tropical Aquatic Systems; Connell, D.M., Hawker, D.W., Eds.; CRC Press: Boca Raton, FL, USA, 1991; pp. 129–145. [Google Scholar]
- Lovelock, C.E.; Feller, I.C.; Ball, M.C.; Engelbrecht, B.M.J.; Ewe, M.L. Differences in plant function in phosphorus- and nitrogen-limited mangrove ecosystems. New Phytol. 2006, 172, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, A.; Tsutsuki, K.; Inoue, Y.; Maie, N.; Melling, L.; Jaffé, R. Composition of dissolved organic nitrogen in rivers associated with wetlands. Sci. Total Environ. 2014, 493, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Alongi, D.M.; Pfitzner, J.; Trott, L.A.; Tirendi, F.; Dixon, P.; Klumpp, D.W. Rapid sediment accumulation and microbial mineralization in forests of the mangrove Kandelia candel in the Jiulongjiang Estuary, China. Estuar. Coast. Shelf Sci. 2005, 63, 605–618. [Google Scholar] [CrossRef]
- Boto, K.G.; Wellington, J.T. Soil characteristics and nutrient status in northern Australian mangrove forests. Estuaries 1984, 7, 61–69. [Google Scholar] [CrossRef]
- Alongi, D.M. The dynamics of benthic nutrient pools and fluxes in tropical mangrove forests. J. Mar. Res. 1996, 54, 123–148. [Google Scholar] [CrossRef]
- Naidoo, G. Effects of salinity and nitrogen on growth and water relations in the mangrove, Avicennia marina (Forsk.) Vierh. New Phytol. 1987, 107, 317–325. [Google Scholar] [CrossRef]
- Naidoo, G. Effects of nitrate, ammonium and salinity on growth of the mangrove Bruguiera gymnorrhiza (L.) Lam. Aquat. Bot. 1990, 8, 209–219. [Google Scholar] [CrossRef]
- Shiau, Y.J.; Lee, S.C.; Chen, T.H.; Tian, G.; Chiu, C.Y. Water salinity effects on growth and nitrogen assimilation rate of mangrove (Kandelia candel) seedlings. Aquat. Bot. 2017, 137, 50–55. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, X.; Wang, Y.; Jiang, Z.; Ma, X.; Inyang, A.I.; Cheng, H. Effects of salt on root aeration, nitrification, and nitrogen uptake in mangroves. Forests 2019, 10, 1131. [Google Scholar] [CrossRef]
- Lalnunzira, C.; Tripathi, S.K. Leaf and root production, decomposition and carbon and nitrogen fluxes during stand development in tropical moist forests, north-east India. Soil Res. 2018, 56, 306–317. [Google Scholar] [CrossRef]
- Leuschner, C.; Harteveld, M.; Hertel, D. Consequences of increasing forest use intensity for biomass, morphology, and growth of fine roots in a tropical moist forest on Sulawesi, Indonesia. Agric. Ecosyst. Environ. 2009, 129, 474–481. [Google Scholar] [CrossRef]
- Valverde-Barrantes, O.J.; Raich, J.W.; Russell, A.E. Fine-root mass, growth and nitrogen content for six tropical tree species. Plant Soil 2007, 290, 357–370. [Google Scholar] [CrossRef]
- Asante, W.; Jengre, N. Carbon Stocks and Soil Nutrient Dynamics in the Peat Swamp Forests of the Amanzule Wetlands and Ankobra River Basin. In USAID Integrated Coastal and Fisheries Governance Program for the Western Region of Ghana; Nature Conservation and Research Centre: Accra, Ghana, 2012; p. 45. [Google Scholar]
- Vitousek, P.M.; Sanford, R.L., Jr. Nutrient cycling in moist tropical forest. Annu. Rev. Ecol. Syst. 1986, 17, 137–167. [Google Scholar] [CrossRef]
- Martin, A.R.; Erickson, D.L.; Kress, W.J.; Thomas, S.C. Wood nitrogen concentrations in tropical trees: Phylogenetic patterns and ecological correlates. New Phytol. 2014, 204, 484–495. [Google Scholar] [CrossRef]
- Herbohn, J.L.; Congdon, R.A. Ecosystem dynamics at disturbed and undisturbed sites in North Queensland wet tropical rain forest. III. Nutrient returns to the forest floor through litterfall. J. Trop. Ecol. 1998, 14, 217–229. [Google Scholar] [CrossRef]
- Msunaga, T.; Kubota, D.; Hotta, M.; Wakatsuki, T. Nutritional characteristics of mineral elements in tree species of tropical rain forest, West Sumatra, Indonesia. Soil Sci. Plant Nutr. 1997, 43, 405–418. [Google Scholar] [CrossRef]
- Paoli, G.D.; Curran, L.M. Soil nutrients limit fine litter production and tree growth in mature lowland forest of southwestern Borneo. Ecosystems 2007, 10, 503–518. [Google Scholar] [CrossRef]
- Sugihara, S.; Shibata, M.; Mvondo Ze, A.D.; Araki, S.; Funakawa, S. Effect of vegetation on soil C, N, P and other minerals in oxisols at the forest-savanna transition zone on central Africa. Soil Sci. Plant Nutr. 2014, 60, 45–59. [Google Scholar] [CrossRef]
- Martinelli, L.A.; Piccolo, M.C.; Townsend, A.R.; Vitousek, P.M.; Cuevas, E.; McDowell, W.; Robertson, G.P.; Santos, O.C.; Treseder, K. Nitrogen stable isotopic composition of leaves and soil: Tropical versus temperate forests. Biogeochemistry 1999, 46, 45–65. [Google Scholar] [CrossRef]
- Schurr, E.A.G.; Matson, P.A. Net primary productivity and nutrient cycling across a mesic to wet precipitation gradient in Hawaiian montane forest. Oecologia 2001, 128, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Proctor, J.; Anderson, J.; Chai, P.; Vallack, H.W. Ecological studies in four contrasting lowland rain forests in Gunung Mulu National Park, Sarawak. I. Forest environment, structure and floristics. J. Ecol. 1983, 71, 237–260. [Google Scholar] [CrossRef]
- Shibata, M.; Sugihara, S.; Mvondo-Ze, A.D.; Taraki, S.; Funakawa, S. Nitrogen flux patterns through oxisols and ultisols in tropical forests of Cameroon, central Africa. Soil Sci. Plant Nutr. 2017, 63, 306–317. [Google Scholar] [CrossRef]
- Weintraub, S.R.; Taylor, P.G.; Porder, S.; Asner, G.P. Topographic controls on soil nitrogen availability in a lowland tropical forest. Ecology 2015, 96, 1561–1574. [Google Scholar] [CrossRef]
- Reed, S.C.; Cleveland, C.C.; Townsend, A.R. Tree species control rates of free-living nitrogen fixation in a tropical rain forest. Ecology 2008, 89, 2924–2934. [Google Scholar] [CrossRef]
- Tuah, S.J.; Jamal, Y.M.; Limin, S.H. Nutritional characteristics in leaves of plants native to tropical peat swamps and heath forests of central Kalimantan, Indonesia. Tropics 2003, 12, 224–232. [Google Scholar] [CrossRef]
- Powers, J.S.; Schlesinger, W.H. Relationships among soil carbon distributions and biophysical factors at nested spatial scales in rain forests of northeastern Costa Rica. Geoderma 2002, 109, 165–190. [Google Scholar] [CrossRef]
- Werner, C.; Kiese, R.; Butterbach-Bahl, K. Soil-atmosphere exchange of N2O, CH4, and CO2 and controlling environmental factors for tropical rain forest sites in western Kenya. J. Geophys. Res. 2007, 112, D03308. [Google Scholar] [CrossRef]
- Koehler, B.; Corre, M.D.; Veldkamp, E.; Wullaert, H.; Wright, S.J. Immediate and long-term nitrogen oxide emissions from tropical forest soils exposed to elevated nitrogen input. Glob. Chang. Biol. 2009, 15, 2049–2066. [Google Scholar] [CrossRef]
- Neto, E.S.; Carmo, J.B.; Keller, M.; Martins, S.C.; Alves, L.F.; Vieira, S.A.; Piccolo, M.C.; Camargo, P.; Couto, H.T.Z.; Joly, C.A.; et al. Soil-atmosphere exchange of nitrous oxide, methane and carbon dioxide in a gradient of elevation in the coastal Brazilian Atlantic forest. Biogeosciences 2011, 8, 733–742. [Google Scholar] [CrossRef]
- Palmiotto, P.A.; Davies, S.J.; Vogt, K.A.; Ashton, M.S.; Vogt, D.J.; Ashton, P.S. Soil-related habitat specialization in dipterocarp rain forest tree species in Borneo. J. Ecol. 2004, 92, 609–623. [Google Scholar] [CrossRef]
- Vance, E.D.; Nadkarni, N.M. Microbial biomass and activity in canopy organic matter and the forest floor of a tropical cloud forest. Soil Biol. Biochem. 1990, 22, 677–684. [Google Scholar] [CrossRef]
- Alongi, D.M. Global significance of mangrove blue carbon in climate change mitigation. Sci 2020, 2, 67. [Google Scholar] [CrossRef]
- Naidoo, G. Differential effects of nitrogen and phosphorus enrichment on growth of dwarf Avicennia marina mangroves. Aquat. Bot. 2009, 90, 184–190. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Ruess, R.W.; Feller, I.C. Fine root respiration in the mangrove Rhizophora mangle over variation in forest stature and nutrient availability. Tree Physiol. 2006, 26, 1601–1606. [Google Scholar] [CrossRef]
- Lee, R.Y.; Porubsky, W.P.; Feller, I.C.; Mc Kee, K.L.; Joye, S.B. Porewater biogeochemistry and soil metabolism in dwarf red mangrove habitats (Twin Cays, Belize). Biogeochemistry 2008, 87, 181–198. [Google Scholar] [CrossRef]
- Lacerda, L.D.; Ittekkot, V.; Patchineelam, S.R. Biogeochemistry of mangrove soil organic matter: A comparison between Rhizophora and Avicennia soils in south-eastern Brazil. Estuar. Coast. Shelf Sci. 1995, 40, 713–720. [Google Scholar] [CrossRef]
- Chen, R.; Twilley, R.R. A simulation model of organic matter and nutrient accumulation in mangrove wetland soils. Biogeochemistry 1999, 44, 93–118. [Google Scholar] [CrossRef]
- Krauss, K.W.; Doyle, T.W.; Twilley, R.R.; Rivera-Monroy, V.H.; Sullivan, J.K. Evaluating the relative contributions of hydroperiod and soil fertility on growth of south Florida mangroves. Hydrobiologia 2006, 569, 311–324. [Google Scholar] [CrossRef]
- Rovai, A.S.; Twilley, R.R.; Castañeda-Moya, E.; Riul, P.; Cifuentes-Jara, M.; Manrow-Villalobos, M.; Horta, P.A.; Simonassi, J.C.; Fonseca, A.L.; Pagliosa, P.R. Global controls on carbon storage in mangrove soils. Nat. Clim. Chan. 2018, 3, 534–538. [Google Scholar] [CrossRef]
- Bala Krishna Prasad, M.; Ramanathan, A.L. Sedimentary nutrient dynamics in a tropical estuarine mangrove ecosystem. Estuar. Coast. Shelf Sci. 2008, 80, 60–66. [Google Scholar] [CrossRef]
- Alongi, D.M.; Tirendi, F.; Clough, B.F. Below-ground decomposition of organic matter in forests of the mangroves Rhizophora stylosa and Avicennia marina along the arid coast of Western Australia. Aquat. Bot. 2000, 68, 97–122. [Google Scholar] [CrossRef]
- Alongi, D.M.; Christoffersen, P.; Tirendi, F. The influence of microbial-nutrient relationships in tropical mangrove sediments. J. Exp. Mar. Biol. Ecol. 1993, 171, 201–223. [Google Scholar] [CrossRef]
- Alongi, D.M.; de Carvalho, N.A. The effect of small-scale logging on stand characteristics and soil biogeochemistry in mangrove forests of Timor Leste. For. Ecol. Manag. 2008, 255, 1359–1366. [Google Scholar] [CrossRef]
- Alongi, D.M.; Tirendi, F.; Dixon, P.; Trott, L.A.; Brunskill, G.J. Mineralization of organic matter in intertidal sediments of a tropical semi-enclosed delta. Estuar. Coast. Shelf Sci. 1999, 48, 451–467. [Google Scholar] [CrossRef]
- Alongi, D.M.; Ramanathan, A.L.; Kannan, L.; Tirendi, F.; Trott, L.A.; Bala Krishna Prasad, M. Influence of human-induced disturbance on benthic microbial metabolism in the Pichavaram mangroves, Vellar-Coleroon estuarine complex, India. Mar. Biol. 2005, 147, 1033–1044. [Google Scholar] [CrossRef]
- Alongi, D.M. Bacterial productivity and microbial biomass in tropical mangrove sediments. Microb. Ecol. 1988, 15, 59–79. [Google Scholar] [CrossRef]
- Alongi, D.M.; de Carvalho, N.A.; Amaral, A.L.; da Costa, A.; Trott, L.A.; Tirendi, F. Uncoupled surface and below-ground soil respiration in mangroves: Implications for estimates of dissolved inorganic carbon export. Biogeochemistry 2012, 109, 151–162. [Google Scholar] [CrossRef]
- Alongi, D.M.; Trott, L.A.; Tirendi, F.; McKinnon, A.D.; Undu, M.C. Growth and development of mangrove forests overlying smothered coral reefs, Sulawesi and Sumatra, Indonesia. Mar. Ecol. Prog. Ser. 2008, 370, 97–109. [Google Scholar] [CrossRef][Green Version]
- Alongi, D.M.; Tirendi, F.; Trott, L.A.; Xuan, T.T. Benthic decomposition rates and pathways in plantations of the mangrove Rhizophora apiculata in the Mekong delta, Vietnam. Mar. Ecol. Prog. Ser. 2000, 194, 87–101. [Google Scholar] [CrossRef]
- Alongi, D.M.; Trott, L.A.; Wattayakorn, G.; Clough, B.F. Below-ground nitrogen cycling in relation to net canopy production in mangrove forests of southern Thailand. Mar. Biol. 2002, 140, 855–864. [Google Scholar]
- Kristensen, E.; Holmer, M.; Banta, G.T.; Jensen, M.H.; Hansen, K. Carbon, nitrogen and sulfur cycling in sediments of the Ao Nam Bor mangrove forest, Phuket, Thailand: A review. Phuket Mar. Biol. Cent. Res. Bull. 1995, 60, 37–64. [Google Scholar]
- Nguyen, H.T.; Yoneda, R.; Ninomiya, I.; Harada, K.; Van Dao, T.; Sy, T.M.; Phan, H.N. The effects of stand-age and inundation on carbon accumulation in mangrove plantation soil in Namdinh, northern Vietnam. Tropics 2004, 14, 24–34. [Google Scholar] [CrossRef][Green Version]
- Castañeda-Moya, E.; Twilley, R.R.; Rivera-Monroy, V.H.; Zhang, K.; Davis, S.E., III; Ross, M. Sediment and nutrient deposition associated with Hurricane Wilma in mangroves of the Florida Coastal Everglades. Estuar. Coast. 2010, 33, 45–58. [Google Scholar] [CrossRef]
- Garcias-Bonet, N.; Delgado-Huertas, A.; Carrillo-de-Albornoz, P.; Anton, A.; Almahasheer, H.; Marbá, N.; Hendricks, I.E.; Krause-Jensen, D.; Duarte, C.M. Carbon and nitrogen concentrations, stocks, and isotopic compositions in Red Sea seagrass and mangrove sediments. Front. Mar. Sci. 2019, 6, 267. [Google Scholar] [CrossRef]
- Rivera-Monroy, V.H.; Twilley, R.R.; Medina, E.; Moser, E.B.; Botero, L.; Francisco, A.M.; Bullard, E. Spatial variability of soil nutrients in disturbed riverine mangrove forests at different stages of regeneration in the San Juan River estuary, Venezuela. Estuaries 2004, 27, 44–57. [Google Scholar] [CrossRef]
- Gandaseca, S.; Pazi, A.M.M.; Zulkipli, M.N.S.; Hamzah, A.H.; Zaki, P.H.; Abdu, A. Assessment of nitrogen and phosphorus in mangrove forest soil at Awat-Awat Lawas Sarawak. Am. J. Agricult. For. 2016, 4, 136–139. [Google Scholar] [CrossRef]
- Sarker, S.; Masud-Ul-Alam, M.; Hossain, M.S.; Chowdhury, S.R.; Sharifuzzaman, S.M. A review of bioturbation and sediment organic geochemistry in mangroves. Geol. J. 2020. [Google Scholar] [CrossRef]
- Sofawi, A.B.; Nazri, M.N.; Rozainah, M.Z. Nutrient availability in mangrove soil: Anthropogenic, seasonal and depth variation factors. Appl. Ecol. Environ. Res. 2017, 15, 1983–1998. [Google Scholar] [CrossRef]
- Shilla, D.J.; Shilla, D.A. Assessment of the geochemical characteristics of water and surface sediments of Rufiji mangrove forest, Tanzania. Tanzan. J. Sci. 2020, 46, 482–497. [Google Scholar]
- Gutiérrez, J.C.S.; Ponce-Palafox, J.T.; Pineda-Jaimes, N.B.; Arenas-Fuentes, V.; Arredondo-Figueroa, J.L.; Cifuentes-Lemus, J.L. Comparison of the mangrove soil with different levels of disturbance in tropical Agua Brava Lagoon, Mexican Pacific. Appl. Ecol. Environ. Res. 2016, 14, 45–57. [Google Scholar] [CrossRef]
- Ramanathan, A.L.; Datta, D.K.; Ghosh, P.; Kaushal, S.; Murtudde, R. Tracing Nitrogen and Carbon Biogeochemical Processes in the Intertidal Mangrove Ecosystem (Sundarbans) of India and Bangladesh: Implications for the Global Environmental Change; Final Report for APN Project: ARCP2012-07CMY-Ramanathan; Asia-Pacific Network for Global Change Research: Kobe, Japan, 2012. [Google Scholar]
- Scharler, U.M.; Ulanowicz, R.E.; Fogel, M.L.; Wooller, M.J.; Jacobson-Meyers, M.E.; Lovelock, C.E.; Feller, I.C.; Frischer, M.; Lee, R.; McKee, K.; et al. Variable nutrient stoichiometry (carbon:nitrogen:phosphorus) across trophic levels determines community and ecosystem properties in an oligotrophic mangrove system. Oecologia 2015, 179, 863–876. [Google Scholar] [CrossRef]
- Nirmal Kumar, I.J.; Sajish, P.R.; Nirmal Kumar, R.; Bash, G.; Shailendra, V. Nutrient dynamics in an Avicennia marina (Forsk.) Vierh., mangrove forest in Vamleshwar, Gujarat, India. Not. Sci. Biol. 2011, 3, 51–56. [Google Scholar] [CrossRef]
- Nordhaus, I.; Salewski, T.; Jennerjahn, T.C. Interspecific variations in mangrove leaf litter decomposition are related to labile nitrogenous compounds. Estuar. Coast. Shelf Sci. 2017, 192, 137–143. [Google Scholar] [CrossRef]
- Bulmer, R.H.; Schwendenmann, L.; Lundquist, C.J. Carbon and nitrogen stocks and below-ground allometry in temperate mangroves. Front. Mar. Sci. 2016, 3, 150. [Google Scholar] [CrossRef]
- Alongi, D.M.; Clough, B.F.; Dixon, P.; Tirendi, F. Nutrient partitioning and storage in arid-zone forests of the mangroves Rhizophora stylosa and Avicennia marina. Trees 2003, 17, 51–60. [Google Scholar] [CrossRef]
- Marchand, C.; Lallier-Vergès, E.; Baltzer, F. The composition of sedimentary organic matter in relation to the dynamic features of the mangrove-fringed coast of French Guiana. Estuar. Coast. Shelf Sci. 2003, 56, 119–130. [Google Scholar] [CrossRef]
- Enamul Hoq, M.; Islam, M.L.; Paul, H.K.; Ahmed, S.U.; Islam, M.N. Decomposition and seasonal changes in nutrient constituents in mangrove litter of Sundarbans mangrove, Bangladesh. Ind. J. Mar. Sci. 2002, 31, 130–135. [Google Scholar]
- Haryadi, J.; Basukriandi, A. The study on mangrove litter as a source of nutrients for Blanakan mangrove pond, Subang, West Java. Indo. Aquacult. J. 2013, 8, 55–64. [Google Scholar]
- Nordhaus, I.; Salewski, T.; Jennerjahn, T.C. Food preferences on mangrove crabs related to leaf nitrogen compounds in the Segara Anakan Lagoon, Java, Indonesia. J. Sea Res. 2011, 65, 414–426. [Google Scholar] [CrossRef]
- Twilley, R.R.; Lugo, A.E.; Patterson-Zucca, C. Litter production and turnover in basin mangrove forests in southwest Florida. Ecology 1986, 67, 670–683. [Google Scholar] [CrossRef]
- Wafar, S.; Untawale, A.G.; Wafar, M. Litter fall and energy flux in a mangrove ecosystem. Estuar. Coast. Shelf Sci. 1997, 44, 111–124. [Google Scholar] [CrossRef]
- Hemati, Z.; Hossian, M.; Rozainah, M.Z. Determination of carbon and nitrogen in litter fall of mangrove ecosystem in Peninsular Malaysia. Pak. J. Bot. 2017, 49, 1381–1386. [Google Scholar]
- Ye, Y.; Chen, Y.P.; Chen, G.C. Litter production and litter elemental composition in two rehabilitated Kandelia obovata mangrove forests in Jiulongjiang estuary, China. Mar. Environ. Res. 2013, 83, 63–72. [Google Scholar] [CrossRef]
- Srisunont, C.; Jaiyen, T.; Tenrung, M.; Likitchaikul, M.; Srisunont, T. Nutrient accumulation by litterfall in mangrove forest at Klong Khone, Thailand. Sci. Technol. Asia 2017, 9–18. [Google Scholar] [CrossRef]
- Adame, M.F.; Zacaria, R.M.; Fry, B.; Chong, V.C.; Then, Y.H.A.; Brown, C.J.; Lee, S.Y. Loss and recovery of carbon and nitrogen after mangrove clearing. Ocean. Coast. Manag. 2018, 161, 117–126. [Google Scholar] [CrossRef]
- Lugo, A.E.; Medina, E.; Cuevas, E.; Laboy Nieves, E.N.; Schäeffer Novelli, Y. Ecophysiology of a mangrove forest in Jobos Bay, Puerto Rico. Carrib. J. Sci. 2007, 43, 200–219. [Google Scholar] [CrossRef]
- Pinto, L. Litterfall and its element content in the Pagbilao Mangrove Forest Reserve, Philippines. Mahasagar 1992, 25, 97–104. [Google Scholar]
- Pedrosa Fragoso, C.; Bernini, E.; Ferreira Araújo, B.; Gomes de Almeida, M.; Eduardo de Rezende, C. Mercury in litterfall and sediment using elemental and isotopic composition of carbon and nitrogen in the mangrove of southeastern Brazil. Estuar. Coast. Shelf Sci. 2018, 202, 30–39. [Google Scholar] [CrossRef]
- Bunt, J.S. Studies of Mangrove Litter Fall in Tropical Australia. In Mangrove Ecosystems in Australia: Structure, Function and Management; Clough, B.F., Ed.; Australian National University Press: Canberra, Australia, 1982; pp. 221–237. [Google Scholar]
- Woodroffe, C.D.; Bardsley, K.N.; Ward, P.J.; Hanley, J.R. Production of mangrove litter in a macrotidal embayment, Darwin Harbour, N.T., Australia. Estuar. Coast. Shelf Sci. 1988, 26, 581–598. [Google Scholar] [CrossRef]
- Thongtham, N.; Kristensen, E. Carbon and nitrogen balance of leaf-eating sesarmid crabs (Neoepisesarma versicolor) offered different food sources. Estuar. Coast. Shelf Sci. 2005, 65, 213–222. [Google Scholar] [CrossRef]
- Thongtham, N.; Kristensen, E.; Puangprasan, S.-Y. Leaf removal by sesarmid crabs in Bangrong mangrove forests, Phuket, Thailand; with emphasis on the feeding ecology of Neoepisesarma versicolor. Estuar. Coast. Shelf Sci. 2008, 80, 583–590. [Google Scholar] [CrossRef]
- Feller, I.C.; McKee, K.L.; Whigham, D.F.; O’Neill, J.P. Nitrogen vs. phosphorus limitation across an ecotonal gradient in a mangrove forest. Biogeochemistry 2002, 62, 145–175. [Google Scholar] [CrossRef]
- Saravanakumar, A.; Rajkumar, M.; Sesh Serebiah, J.; Thivakaran, G.A. Seasonal variations in physico-chemical characteristics of water, sediment and soil texture in arid zone mangroves of Kachchh-Gujarat. J. Environ. Biol. 2008, 29, 725–732. [Google Scholar]
- Nabiul Islam Khan, M.; Suwa, R.; Hagihara, A. Carbon and nitrogen pools in a mangrove stand of Kandelia obovata (S.L.) Yong: Vertical distribution in the soil-vegetation system. Wetlands Ecol. Manag. 2007, 15, 141–153. [Google Scholar] [CrossRef]
- Mantiquilla, J.A.; Salmasan, S.F.D.; Obelidhon, M.K.A.; Abad, R.G. Nutrient status of Nipa (Nypa fruticans Wurmb.) in selected areas of Mindanao, the Philippines. Banwa B 2019, 14, art012. [Google Scholar]
- Medina, E.; Francisco, M. Osmolality and δ13C of leaf tissues of mangrove species from environments of contrasting rainfall and salinity. Estuar. Coast. Shelf Sci. 1997, 45, 337–344. [Google Scholar] [CrossRef]
- Alongi, D.M.; Wattayakorn, G.; Tirendi, F.; Dixon, P. Nutrient capital in different aged forests of the mangrove Rhizophora apiculata. Bot. Mar. 2004, 47, 116–124. [Google Scholar] [CrossRef]
- Tognella, M.M.P.; Soares, M.L.G.; Cuevas, E.; Medina, E. Heterogeneity of elemental composition and natural abundance of stable isotopes of C and N in soils and leaves of mangroves at their southernmost West Atlantic range. Braz. J. Biol. 2016, 76, 994–1003. [Google Scholar] [CrossRef]
- Jayasekera, R. Chemical composition of the mangrove, Rhizophora mangle L. J. Plant Physiol. 1991, 138, 119–121. [Google Scholar] [CrossRef]
- Feller, I.C.; Lovelock, C.E.; McKee, K.L. Nutrient addition differentially affects ecological processes of Avicennia germinans in nitrogen versus phosphorus limited mangrove ecosystems. Ecosystems 2007, 10, 347–359. [Google Scholar] [CrossRef]
- Telave, A.B. Ecophysiological studies on Sonneratia L. from the coast of Maharashtra, India. Ind. J. Geo-Mar. Sci. 2015, 44, 1239–1244. [Google Scholar]
- Muzuka, A.N.N.; Shunula, J.P. Stable isotope compositions of organic carbon and nitrogen of two mangrove stands along the Tanzanian coastal zone. Estuar. Coast. Shelf Sci. 2006, 66, 447–458. [Google Scholar] [CrossRef]
- Bernini, E.; Silva, M.A.B.D.; Carmo, T.M.S.D.; Cuzzuol, G.R.F. Composição química do sediment e de folhas das espécies do manguezal do estuário do Rio São Mateus, Espírito Santo, Brasil. Rev. Brasil Bot. 2006, 29, 689–699. [Google Scholar] [CrossRef]
- Medina, E.; Giarrizzo, T.; Menezes, M.; Carvalho Lira, M.; Carvalho, E.A.; Peres, A.; Silva, B.; Vilhena, R.; Reise, A.A.; Braga, F.C. Mangal communities of the “Salgado Paraense”: Ecological heterogeneity along the Bragança peninsula assessed through soil and leaf analysis. Amazonia 2001, 16, 397–416. [Google Scholar]
- Lovelock, C.E.; Feller, I.C.; McKee, K.L.; Engelbrecht, B.M.J.; Ball, M.C. The effect of nutrient enrichment on growth, photosynthesis and hydraulic conductance of dwarf mangroves in Panama. Funct. Ecol. 2004, 18, 25–33. [Google Scholar] [CrossRef]
- Lin, Y.-M.; Liu, X.-W.; Zhang, H.; Fan, H.-Q.; Lin, G.-H. Nutrient conservation strategies of a mangrove species Rhizophora stylosa under nutrient limitation. Plant Soil 2010, 326, 469–479. [Google Scholar] [CrossRef]
- Gritcan, I.; Duxbury, M.; Leuzinger, S.; Alfaro, A.C. Leaf stable isotope and nutrient status of temperate mangroves as ecological indicators to assess anthropogenic activity and recovery from eutrophication. Front. Plant Sci. 2016, 7, 1922. [Google Scholar] [CrossRef]
- Simpson, L.T.; Lovelock, C.E.; Cherry, J.A.; Feller, I.C. Short-lived effects of nutrient enrichment on Avicennia germinans decomposition in a saltmarsh-mangrove ecotone. Estuar. Coast. Shelf Sci. 2020, 235, 106598. [Google Scholar] [CrossRef]
- Ariyanto, D.; Gunawan, H.; Puspitasari, D.; Susanti Ningsih, S.; Jayanegara, A.; Hamim, H. The differences of the element content in Rhizophora mucronata leaves from Asahan Regency, North Sumatra, Indonesia. Pol. J. Natur. Sci. 2019, 34, 481–491. [Google Scholar]
- Li, M.S. Nutrient dynamics of a Futian mangrove forest in Shenzhen, South China. Estuar. Coast. Shelf Sci. 1997, 45, 463–472. [Google Scholar] [CrossRef]
- Bernini, E.; da Silva, M.A.B.; do Carmo, T.M.S.; Cuzzuol, G.R.F. Spatial and temporal variation of the nutrients in the sediment and leaves of two Brazilian mangrove species and their role in the retention of environmental heavy metals. Braz. J. Plant Physiol. 2010, 22, 177–187. [Google Scholar] [CrossRef]
- Ahmed, A.; Ohlson, M.; Hoque, S.; Moula, M.G. Chemical composition of leaves of a mangrove tree (Sonneratia apetala Buch.Ham.) and their correlation with some soil variables. Bangladesh J. Bot. 2010, 39, 61–69. [Google Scholar] [CrossRef]
- Mfilinge, P.L.; Atta, N.; Tsuchiya, M. Nutrient dynamics and leaf litter decomposition in a subtropical mangrove forest at Oura Bay, Okinawa, Japan. Trees 2002, 16, 172–180. [Google Scholar] [CrossRef]
- Saatchi, S.S.; Harris, N.L.; Brown, S.; Lefsky, M.; Mitchard, E.T.A.; Salas, W.; Zutta, B.R.; Buermann, W.; Lewis, S.L.; Hagen, S.; et al. Benchmark map of forest carbon stocks in tropical regions across three continents. Proc. Natl. Acad. Sci. USA 2011, 108, 9899–9904. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.J.P.; Talbot, J.; Lewis, S.L.; Phillips, O.L.; Qie, L.; Begne, S.K.; Chave, J.; Cuni-Sanchez, A.; Hubau, W.; Lopez-Gonzalez, G.; et al. Diversity and carbon storage across the tropical forest biome. Sci. Rep. 2017, 7, 39102. [Google Scholar] [CrossRef]
- Wantzen, K.M.; Couto, E.G.; Mund, E.E.; Amorim, R.S.S.; Siqueira, A.; Tielbörger, K.; Seifan, M. Soil carbon stocks in stream-valley-ecosystems in the Brazilian Cerrado agroscape. Agric. Ecosyst. Environ. 2012, 151, 70–79. [Google Scholar] [CrossRef]
- Descloux, S.; Chanudet, V.; Poilvé, H.; Grégoire, A. Co-assessment of biomass and soil organic carbon stocks in a future reservoir area located in Southeast Asia. Environ. Monit. Assess. 2011, 173, 723–741. [Google Scholar] [CrossRef]
- Laumonier, Y.; Edin, A.; Kanninen, M.; Munandar, A.W. Landscape-scale variation in the structure and biomass of the hill dipterocarp forest of Sumatra: Implications for carbon stock assessments. For. Ecol. Manag. 2010, 259, 505–513. [Google Scholar] [CrossRef]
- Yuen, J.Q.; Ziegler, A.D.; Webb, E.L.; Ryan, C.M. Uncertainty in below-ground carbon biomass for major land covers in Southeast Asia. For. Ecol. Manag. 2013, 310, 915–926. [Google Scholar] [CrossRef]
- Powers, J.S.; Treseder, K.K.; Lerdau, M.T. Fine roots, arbuscular mycorrhizal hyphae, and soil nutrients in four neotropical rain forests: Patterns across large geographic distances. New Phytol. 2005, 165, 913–921. [Google Scholar] [CrossRef]
- Cusack, D.F.; Markesteijn, L.; Condit, R.; Lewis, O.T.; Turner, B.L. Soil carbon stocks across tropical forests of Panama regulated by base cation effects on fine roots. Biogeochemistry 2018, 137, 253–266. [Google Scholar] [CrossRef]
- Thamdrup, B. New pathways and processes in the global nitrogen cycle. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 407–428. [Google Scholar] [CrossRef]
- Mukherji, S.; Haldar, S.; Ghosh, A. Investigation of the Structural and Functional Microbial Diversity in Indian Mangroves. In Microorganisms in Saline Environments: Strategies and Functions; Giri, B., Varma, A., Eds.; Springer Nature: Gland, Switzerland, 2019; pp. 93–130. [Google Scholar]
- Cheung, M.K.; Wong, C.K.; Chu, K.H.; Kwan, H.S. Community structure, dynamics and interactions of bacteria, archaea and fungi in subtropical coastal wetland sediments. Sci. Rep. 2018, 8, 14397. [Google Scholar] [CrossRef] [PubMed]
- Loganathachetti, D.S.; Poosakkannu, A.; Muthuraman, S. Fungal community assemblage of different soil compartments in mangrove ecosystem. Sci. Rep. 2017, 7, 8560. [Google Scholar] [CrossRef] [PubMed]
- Marcos, M.S.; Barboza, A.D.; Keijzer, R.M.; Laanbroek, H.J. Tide as steering factor in structuring archaeal and bacterial ammonia-oxidizing communities in mangrove forest soils dominated by Avicennia germinans and Rhizophora mangle. Microb. Ecol. 2018, 75, 997–1008. [Google Scholar] [CrossRef]
- Li, R.; Tong, T.; Wu, S.; Chai, M.; Xie, S. Multiple factors govern the biogeographic distribution of archaeal community in mangrove across China. Estuar. Coast. Shelf Sci. 2019, 231, 106414. [Google Scholar] [CrossRef]
- Bai, S.; Li, J.; He, Z.; Van Nostrand, J.D.; Tian, Y.; Lin, G.; Zhou, J.; Zheng, T. GeoChip-based analysis of the functional gene diversity and metabolic potential of soil microbial communities in mangroves. Appl. Microbiol. Biotechnol. 2013, 97, 7035–7048. [Google Scholar] [CrossRef]
- Zhang, M.; Luo, Y.; Lin, L.; Lin, X.; Hetharua, B.; Zhao, W.; Zhou, M.; Zhan, Q.; Xu, H.; Zheng, T.; et al. Molecular and stable isotope evidence for the occurrence of nitrite-dependent anaerobic methane-oxidizing bacteria in the mangrove sediment of Zhangjiang estuary, China. Appl. Microbiol. Biotechnol. 2018, 102, 2441–2454. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, E.; Jensen, M.H.; Banta, G.T.; Hansen, K.; Holmer, M.; King, G.M. Transformation and transport of inorganic nitrogen in sediments of a southeast Asian mangrove forest. Aquat. Microb. Ecol. 1998, 15, 165–175. [Google Scholar] [CrossRef]
- Viner, A.B. The Status and Transport of Nutrients through the Purari River (Papua New Guinea). In Purari River Hydroelectric Scheme Environmental Studies; Publication of the Environment and Conservation: Waigani, Papua New Guinea, 1979; Volume 9, pp. 1–28. [Google Scholar]
- Kyaruzi, J.J.; Kyewalyanga, M.S.; Muruke, M.H.S. Cyanobacteria composition and impact of seasonality on their in situ nitrogen fixation rate in a mangrove ecosystem adjacent to Zanzibar town. West. Indian Ocean J. Mar. Sci. 2003, 2, 35–44. [Google Scholar] [CrossRef]
- van der Valk, A.G.; Attiwill, P.M. Acetylene reduction in an Avicennia marina community in southern Australia. Aust. J. Bot. 1984, 32, 157–164. [Google Scholar]
- Lee, R.Y.; Joye, S.B. Seasonal patterns of nitrogen fixation and denitrification in oceanic mangrove habitats. Mar. Ecol. Prog. Ser. 2006, 307, 127–141. [Google Scholar] [CrossRef]
- Joye, S.B.; Lee, R.Y. Benthic microbial mats: Important sources of fixed nitrogen and carbon to the Twin Cays, Belize ecosystem. Atoll Res. Bull. 2004, 528, 1–26. [Google Scholar] [CrossRef]
- Whigham, D.F.; Verhoeven, J.T.A.; Samarkin, V.; Megonigal, P.J. Responses of Avicennia germinans (Black Mangrove) and the soil microbial community to nitrogen addition in a hypersaline wetland. Estuar. Coast. 2009, 32, 926–936. [Google Scholar] [CrossRef]
- Alongi, D.M.; Sasekumar, A.; Chong, V.C.; Pfitzner, J.; Trott, L.A.; Tirendi, F.; Dixon, P.; Brunskill, G.J. Sediment accumulation and organic material flux in a managed mangrove ecosystem: Estimates of land-ocean-atmosphere exchange in peninsular Malaysia. Mar. Geol. 2004, 208, 383–402. [Google Scholar] [CrossRef]
- Twilley, R.R.; Rivera-Monroy, V.H.; Rovai, A.S.; Castañeda-Moya, E.; Davis, S. Mangrove Biogeochemistry at Local and Global Scales Using Ecogeomorphic Approaches. In Coastal Wetlands: An Integrated Ecosystem Approach; Perillo, G.M.E., Wolanski, E., Cahoon, D.R., Hopkinson, C.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 717–785. [Google Scholar]
- Morell, J.M.; Corredor, J.E. Sediment trapping in a mangrove lagoon. Estuar. Coast. Shelf Sci. 1993, 37, 203–212. [Google Scholar] [CrossRef]
- Das, S.; Ganguly, D.; Mukherjee, A.; Chakraborty, S.; De, T.K. Exploration of N2 fixation and denitrification processes in the Sundarbans mangrove ecosystem, India. Ind. J. Geo-Mar. Sci. 2020, 49, 740–747. [Google Scholar]
- Pelegri, S.P.; Rivera-Monroy, V.H.; Twilley, R.R. A comparison of nitrogen fixation (acetylene reduction) among three species of mangrove litter, sediments, and pneumatophores in south Florida, USA. Hydrobiologia 1997, 356, 73–79. [Google Scholar] [CrossRef]
- Shiau, Y.-J.; Lin, M.-F.; Tan, C.-C.; Tian, G.; Chiu, C.-Y. Assessing N2 fixation in estuarine mangrove soils. Estuar. Coast. Shelf Sci. 2017, 189, 84–89. [Google Scholar] [CrossRef]
- Romero, I.C.; Jacobson, M.; Fuhrman, J.A.; Fogel, M.; Capone, D.G. Long-term nitrogen and phosphorus fertilization effects on N2 fixation rates and nifH gene community patterns in mangrove sediments. Mar. Ecol. 2012, 33, 117–127. [Google Scholar] [CrossRef]
- Vovides, A.G.; Lόpez-Portillo, J.; Bashan, Y. N2-fixation along a gradient of long-term disturbance in tropical mangroves bordering the Gulf of Mexico. Biol. Fertil. Soils 2011, 47, 567–576. [Google Scholar] [CrossRef]
- Zuberer, D.A.; Silver, W.S. Biological nitrogen fixation (acetylene reduction) associated with Florida mangroves. Appl. Environ. Microbiol. 1978, 35, 567–575. [Google Scholar] [CrossRef]
- Sheridan, R.P. Epicaulous, nitrogen-fixing microepiphytes in a tropical mangal community, Guadeloupe, French West Indies. Biotropica 1991, 23, 530–541. [Google Scholar] [CrossRef]
- Mann, F.D.; Steinke, T.D. Biological nitrogen fixation (acetylene reduction) associated with blue-green algal (cyanobacterial) communities in the Beachwood Mangrove Nature Reserve. I. The effect of environmental factors on acetylene reduction activity. S. Afr. Tydskr. Plantk. 1989, 55, 438–446. [Google Scholar] [CrossRef]
- Mann, F.D.; Steinke, T.D. Biological nitrogen fixation (acetylene reduction) associated with blue-green algal (cyanobacterial) communities in the Beachwood Mangrove Nature Reserve. II. Seasonal variation in acetylene reduction activity. S. Afr. J. Bot. 1993, 59, 1–8. [Google Scholar]
- Pelagri, S.P.; Twilley, R.R. Heterotrophic nitrogen fixation (acetylene reduction) during leaf-litter decomposition of two mangrove species from south Florida, USA. Mar. Biol. 1998, 131, 53–61. [Google Scholar]
- Sengupta, A.; Chaudhuri, S. Ecology of heterotrophic dinitrogen fixation in the rhizosphere of mangrove plant community at the Ganges River estuary in India. Oecologia 1991, 87, 560–564. [Google Scholar] [CrossRef]
- Vovides, A.G.; Bashan, Y.; Lόpez-Portillo, J.; Guevara, R. Nitrogen fixation in preserved, reforested, naturally regenerated and impaired mangroves as an indicator of functional restoration in mangroves in an arid region of Mexico. Restor. Ecol. 2010, 19, 236–244. [Google Scholar] [CrossRef]
- Sheridan, R.P. Nitrogen fixation by epicaulous cyanobacteria in the Pointe de la Saline mangrove community, Guadeloupe, French West Indies. Biotropica 1992, 24, 571–574. [Google Scholar] [CrossRef]
- Lugomela, C.; Bergman, B. Biological N2-fixation on mangrove pneumatophores: Preliminary observations and perspectives. Ambio 2002, 31, 612–623. [Google Scholar] [CrossRef]
- Potts, M.; Whitton, B.A. Nitrogen fixation by blue-green algal communities in the intertidal zone of the lagoon of Aldabra Atoll. Oecologia 1977, 27, 275–283. [Google Scholar] [CrossRef]
- Potts, M. Nitrogen fixation (acetylene reduction) associated with communities of heterocystous and non-heterocystous blue-green algae in mangrove forests of Sinai. Oecologia 1979, 39, 359–373. [Google Scholar] [CrossRef] [PubMed]
- Boto, K.G.; Robertson, A.I. The relationship between nitrogen fixation and tidal exports of nitrogen in a tropical mangrove system. Estuar. Coast. Shelf Sci. 1990, 31, 531–540. [Google Scholar] [CrossRef]
- Woitchik, A.F.; Ohowa, B.; Kazungu, J.M.; Rao, R.G.; Goeyens, L.; Dehairs, F. Nitrogen enrichment during decomposition of mangrove leaf litter in an east African coastal lagoon (Kenya): Relative importance of biological nitrogen fixation. Biogeochemistry 1997, 39, 15–35. [Google Scholar] [CrossRef]
- Uchino, F.; Hambali, G.G.; Yatazawa, M. Nitrogen-fixing bacteria from warty lenticellate bark of a mangrove tree, Bruguiera gymnorrhiza (L.) Lamk. Appl. Environ. Microbiol. 1984, 47, 44–48. [Google Scholar] [CrossRef]
- Zuberer, D.A.; Silver, W.S. N2-fixation (acetylene reduction) and the microbial colonization of mangrove roots. New Phytol. 1979, 82, 467–471. [Google Scholar] [CrossRef]
- Toledo, G.; Bashan, Y.; Soeldner, A. Cyanobacteria and black mangroves in northwestern Mexico: Colonization, and diurnal and seasonal nitrogen fixation on aerial roots. Can. J. Microbiol. 1995, 41, 999–1011. [Google Scholar] [CrossRef]
- Gotto, J.W.; Taylor, B.F. N2 fixation associated with decaying leaves of the red mangrove (Rhizophora mangle). Appl. Environ. Microbiol. 1976, 31, 781–783. [Google Scholar] [CrossRef] [PubMed]
- Qashqari, M.S.; Garcias-Bonet, N.; Fusi, M.; Booth, J.M.; Daffonchio, D.; Duarte, C.M. High temperature and crab density reduce atmospheric nitrogen fixation in Red Sea mangrove sediments. Estuar. Coast. Shelf Sci. 2020, 232, 106487. [Google Scholar] [CrossRef]
- Capone, D.G. Determination of Nitrogenase Activity in Aquatic Samples Using the Acetylene Reduction Procedure. In Aquatic Microbial Ecology; Kemp, P.F., Sherr, B.F., Sherr, E.B., Cole, J.J., Eds.; Lewis Publishers: Boca Raton, FL, USA, 1993; pp. 621–632. [Google Scholar]
- Wang, H.-T.; Su, J.-Q.; Zheng, T.-L.; Yang, X.-R. Impacts of vegetation, tidal process, and depth on the activities, abundances, and community compositions of denitrifiers in mangrove sediment. Appl. Microbiol. Biotechnol. 2014, 98, 9375–9387. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.O.; Loka Bharathi, P.A. Denitrification: An important pathway for nitrous oxide production in tropical mangrove sediments (Goa, India). J. Environ. Qual. 2010, 39, 1507–1516. [Google Scholar] [CrossRef]
- Fernandes, S.O.; Loka Bharathi, P.A. Nitrate levels modulate denitrification activity in tropical mangrove sediments (Goa, India). Environ. Monit. Assess. 2011, 173, 117–125. [Google Scholar] [CrossRef]
- Kristensen, E.; Mangion, P.; Tang, M.; Flindt, M.R.; Holmer, M.; Ulomi, S. Microbial carbon oxidation rates and pathways in sediments of two Tanzanian mangrove forests. Biogeochemistry 2011, 103, 143–158. [Google Scholar] [CrossRef]
- Gao, G.-F.; Li, P.-F.; Zhong, J.-X.; Shen, Z.-J.; Chen, J.; Li, Y.-T.; Isabew, A.; Zhu, X.-Y.; Ding, Q.-S.; Zhang, S.; et al. Spartina alterniflora invasion alters soil bacterial communities and enhances soil N2O emissions by stimulating soil denitrification in mangrove wetland. Sci. Total Environ. 2019, 653, 231–240. [Google Scholar] [CrossRef]
- Pérez-Villalona, H.; Cornwell, J.C.; Ortiz-Zayas, J.R.; Cuevas, E. Sediment denitrification and nutrient fluxes in the San José Lagoon, a tropical lagoon in the highly urbanized San Juan Bay estuary, Puerto Rico. Estuar. Coast. 2015, 38, 2259–2278. [Google Scholar] [CrossRef]
- Senthilkumar, B. Biogeochemical and Biophysical Aspects of Pichavarum Mangrove Ecosystem, South India. Ph.D. Thesis, Anna University, Chennai, India, 2007. [Google Scholar]
- Rivera-Monroy, V.H.; Twilley, R.R.; Boustany, R.G.; Day, J.W.; Vera-Herrera, F.; del Carmen Ramirez, M. Direct denitrification in mangrove sediments in Terminos Lagoon, Mexico. Mar. Ecol. Prog. Ser. 1995, 126, 97–109. [Google Scholar] [CrossRef]
- Guan, Q.S.; Cao, W.Z.; Wang, M.; Wu, G.J.; Wang, F.F.; Jiang, C.; Tao, Y.R.; Gao, Y. Nitrogen loss through anaerobic ammonium oxidation coupled with iron reduction in a mangrove wetland. Eur. J. Soil Sci. 2018, 69, 732–741. [Google Scholar] [CrossRef]
- Zhang, M.; Dai, P.; Lin, X.; Lin, L.; Hetharua, B.; Zhang, Y.; Tian, Y. Nitrogen loss by anaerobic ammonium oxidation in a mangrove wetland of the Zhangjiang estuary, China. Sci. Total Environ. 2020, 698, 134291. [Google Scholar] [CrossRef]
- Reis, C.R.G.; Nardoto, G.B.; Rochelle, A.L.C.; Vieira, S.A.; Oliveira, R.S. Nitrogen dynamics in subtropical fringe and basin mangrove forests inferred from stable isotopes. Oecologia 2017, 183, 841–848. [Google Scholar] [CrossRef]
- Kristensen, E.; Andersen, F.Ø.; Holmboe, N.; Holmer, M.; Thongtham, N. Carbon and nitrogen mineralization in sediments of the Bangrong mangrove area, Phuket, Thailand. Aquat. Microb. Ecol. 2000, 22, 199–213. [Google Scholar] [CrossRef]
- Nedwell, D.B.; Blackburn, T.H.; Wiebe, W.J. Dynamic nature of the turnover of organic carbon, nitrogen and sulphur in the sediments of a Jamaican mangrove forest. Mar. Ecol. Prog. Ser. 1994, 110, 223–231. [Google Scholar] [CrossRef]
- Cao, W.; Guan, Q.; Li, Y.; Wang, M.; Liu, B. The contribution of denitrification and anaerobic ammonium oxidation to N2 production in mangrove sediments of southeast China. J. Soils Sediments 2017, 17, 1767–1776. [Google Scholar] [CrossRef]
- Krishnan, K.P.; Loka Bharathi, P.A. Organic carbon and iron modulate nitrification rates in mangrove swamps of Goa, south west coast of India. Estuar. Coast. Shelf Sci. 2009, 84, 419–426. [Google Scholar] [CrossRef]
- Dunn, R.J.K.; Welsh, D.T.; Jordan, M.A.; Waltham, N.J.; Lemckert, C.J.; Teasdale, P.R. Benthic metabolism and nitrogen dynamics in a sub-tropical coastal lagoon: Microphytobenthos stimulate nitrification and nitrate reduction through photosynthetic oxygen evolution. Estuar. Coast. Shelf Sci. 2012, 113, 272–282. [Google Scholar] [CrossRef]
- Xiao, K.; Wu, J.; Li, H.; Hong, Y.; Wilson, A.M.; Jiao, J.J.; Shananan, M. Nitrogen fate in a subtropical mangrove swamp: Potential association with seawater-groundwater exchange. Sci. Total Environ. 2018, 635, 586–597. [Google Scholar] [CrossRef]
- Amano, T.; Yoshinaga, I.; Yamagishi, T.; Van Thuoc, C.; The Thu, P.; Ueda, S.; Kato, K.; Sako, Y.; Suwa, Y. Contribution of anammox bacteria to benthic nitrogen cycling in a mangrove forest and shrimp ponds, Haiphong, Vietnam. Microb. Environ. 2011, 26, 1011040240. [Google Scholar] [CrossRef]
- Purvaja, R.; Ramesh, R.; Ray, A.K.; Rixen, T. Nitrogen cycling: A review of the processes, transformations and fluxes in coastal ecosystems. Curr. Sci. 2008, 94, 1419–1438. [Google Scholar]
- Luvizotto, D.M.; Araujo, J.E.; de Cássia, P.; Silva, M.; Dias, A.C.F.; Kraft, B.; Tegetmeye, H.; Strous, M.; Andreote, F.D. The rates and players of denitrification, dissimilatory nitrate reduction to ammonia (DNRA) and anaerobic ammonia oxidation (anammox) in mangrove soils. An. Acad. Bras. Cienc. 2019, 91, e20180373. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.F.; Sobey, M.N.; Smith, C.J.; Rusmana, I.; Phillips, W.; Stott, A.; Osborn, A.M.; Nedwell, D.B. Dissimilatory reduction of nitrate to ammonium, not denitrification or anammox, dominates benthic nitrate reduction in tropical estuaries. Limnol. Oceangr. 2011, 56, 279–291. [Google Scholar] [CrossRef]
- Fernandes, S.O.; Michotey, V.D.; Guasco, S.; Bonin, P.C.; Loka Bharathi, P.A. Denitrification prevails over anammox in tropical mangrove sediments. Mar. Environ. Res. 2012, 74, 9–19. [Google Scholar] [CrossRef]
- Zheng, Y.; Hou, L.; Zhang, Z.; Chen, F.; Gao, D.; Yin, G.; Han, P.; Dong, H.; Liang, X.; Yang, Y.; et al. Anaerobic ammonium oxidation (anammox) bacterial diversity, abundance, and activity in sediments of the Indus estuary. Estuar. Coast. Shelf Sci. 2020, 243, 106925. [Google Scholar] [CrossRef]
- Meyer, R.L.; Risgaard-Petersen, N.; Allen, D.E. Correlation between anammox activity and microscale distribution of nitrite in a subtropical mangrove sediment. Appl. Environ. Microbiol. 2005, 71, 6142–6149. [Google Scholar] [CrossRef]
- Cao, W.; Yang, J.; Li, Y.; Liu, B.; Wang, F.; Chang, C. Dissimilatory nitrate reduction to ammonium conserves nitrogen in anthropogenically affected subtropical mangrove sediments in southeast China. Mar. Pollut. Bull. 2016, 110, 155–161. [Google Scholar] [CrossRef]
- Molnar, N.; Welsh, D.T.; Marchand, C.; Deborde, J.; Meziane, T. Impacts of shrimp farm effluent on water quality, benthic metabolism and N-dynamics in a mangrove forest (New Caledonia). Estuar. Coast. Shelf Sci. 2013, 117, 12–21. [Google Scholar] [CrossRef]
- Dunn, R.J.K.; Welsh, D.T.; Jordan, M.A.; Teasdale, P.R.; Lemckert, C.J. Influence of natural amphipod (Victoriopisa australiensis) (Chilton, 1923) population densities on benthic metabolism, nutrient fluxes, denitrification and DNRA in sub-tropical estuarine sediment. Hydrobiologia 2009, 628, 95–109. [Google Scholar] [CrossRef]
- Fernandes, S.O.; Bonin, P.C.; Michotey, V.D.; Garcia, N.; Loka Bharathi, P.A. Nitrogen-limited mangrove ecosystems conserve N through dissimilatory nitrate reduction to ammonium. Sci. Rep. 2012, 2, 419. [Google Scholar] [CrossRef]
- Iizumi, H. Soil nutrient dynamics. In Workshop on Mangrove Ecosystem Dynamics; Cragg, S., Polunin, N., Eds.; UNESCO: New Delhi, India, 1986; pp. 171–180. [Google Scholar]
- Chen, R.; Twilley, R.R. Patterns of mangrove forest structure and soil nutrient dynamics along the Shark River estuary, Florida. Estuaries 1999, 22, 955–970. [Google Scholar] [CrossRef]
- Chen, J.; Wu, F.-H.; Xiao, Q.; Yang, Z.-H.; Huang, S.-K.; Wang, J.; Wu, Y.-G.; Dong, X.-J.; Pei, Z.-P.; Zheng, H.-L. Diurnal variation in nitric oxide emission flux from a mangrove wetland in Zhangjiang River estuary, China. Estuar. Coast. Shelf Sci. 2010, 90, 212–220. [Google Scholar] [CrossRef]
- Castillo, J.A.A.; Apan, A.A.; Maraseni, T.N.; Salmo, S.G., III. Soil greenhouse gas fluxes in tropical mangrove forests and in land uses on deforested mangrove lands. Catena 2017, 159, 60–69. [Google Scholar] [CrossRef]
- Allen, D.; Dalal, R.C.; Rennenberg, H.; Schmidt, S. Seasonal variation in nitrous oxide and methane emissions from subtropical estuary and coastal mangrove sediments, Australia. Plant Biol. 2011, 13, 126–133. [Google Scholar] [CrossRef]
- Padhy, S.R.; Bhattacharya, P.; Dash, P.K.; Reddy, C.S.; Chakraborty, A.; Pathak, H. Seasonal fluctuations in three mode of greenhouse gas emission in relation to soil labile carbon pools in degraded mangrove, Sundarban, India. Sci. Total Environ. 2020, 705, 135909. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Chen, B.; Yu, D.; Ye, Y.; Tam, N.F.Y.; Chen, S. Soil greenhouse gas emissions reduce the contribution of mangrove plants to the atmospheric cooling effect. Environ. Res. Lett. 2016, 11, 124019. [Google Scholar] [CrossRef]
- Chauhan, R.; Datta, A.; Ramanathan, A.; Adhya, T.K. Factors influencing spatio-temporal variation of methane and nitrous oxide emission from a tropical mangrove of eastern coast of India. Atmos. Environ. 2015, 107, 95–106. [Google Scholar] [CrossRef]
- Konnerup, D.; Betancourt-Portela, J.M.; Villamil, C.; Parra, J.P. Nitrous oxide and methane emissions from the restored mangrove ecosystem of the Ciénaga de Santa Marta, Colombia. Estuar. Coast. Shelf Sci. 2014, 140, 43–51. [Google Scholar] [CrossRef]
- Chen, G.C.; Tam, N.F.Y.; Ye, Y. Summer fluxes of atmospheric greenhouse gases N2O, CH4 and CO2 from mangrove soil in South China. Sci. Total Environ. 2010, 408, 2761–2767. [Google Scholar] [CrossRef]
- Queiroz, H.M.; Artur, A.G.; Taniguchi, C.A.K.; da Silva, M.R.S.; do Nascimento, J.C.; Nόbrega, G.N.; Otero, X.L.; Ferreira, T.O. Hidden contribution of shrimp farming effluents to greenhouse gas emissions from mangrove soils. Estuar. Coast. Shelf Sci. 2019, 221, 8–14. [Google Scholar] [CrossRef]
- Chen, G.C.; Tam, N.F.Y.; Ye, Y. Spatial and seasonal variations of atmospheric N2O and CO2 fluxes from a subtropical mangrove swamp and their relationships with soil characteristics. Soil Biol. Biochem. 2012, 48, 175–181. [Google Scholar] [CrossRef]
- Wang, H.; Liao, G.; D’Souza, M.; Yu, X.; Yang, J.; Yang, X.; Zheng, T. Temporal and spatial variations of greenhouse gas fluxes from a tidal mangrove wetland in southeast China. Environ. Sci. Pollut. Res. 2016, 23, 1873–1885. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.C.; Ulumuddin, Y.I.; Pramudji, S.; Chen, S.Y.; Chen, B.; Ye, Y.; Ou, D.Y.; Ma, Z.Y.; Huang, H.; Wang, J.K. Rich carbon and nitrogen but low atmospheric greenhouse gas fluxes from North Sulawesi mangrove swamps in Indonesia. Sci. Total Environ. 2014, 487, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, R.; Ramanathan, A.L.; Adhya, T.K. Assessment of methane and nitrous oxide flux from mangroves along the east coast of India. Geofluids 2008, 8, 321–332. [Google Scholar] [CrossRef]
- Bauza, J.F.; Morell, J.M.; Corredor, J.E. Biogeochemistry of nitrous oxide production in the red mangrove (Rhizophora mangle) forest sediments. Estuar. Coast. Shelf Sci. 2002, 55, 697–704. [Google Scholar] [CrossRef]
- Allen, D.E.; Dalal, R.C.; Rennenberg, H.; Meyer, R.L.; Reeves, S.; Schmidt, S. Spatial and temporal variation of nitrous oxide and methane flux between subtropical mangrove sediments and the atmosphere. Soil Biol. Biochem. 2007, 39, 622–631. [Google Scholar] [CrossRef]
- Muñoz-Hincapié, M.; Morell, J.M.; Corredor, J.E. Increase of nitrous oxide flux to the atmosphere upon nitrogen addition to red mangroves sediments. Mar. Pollut. Bull. 2002, 44, 992–996. [Google Scholar] [CrossRef]
- Cameron, C.; Hutley, L.B.; Friess, D.A.; Brown, B. High greenhouse gas emissions mitigation benefits from mangrove rehabilitation in Sulawesi, Indonesia. Ecosyst. Serv. 2019, 40, 101035. [Google Scholar] [CrossRef]
- Chen, G.; Chen, J.; Ou, D.; Tam, N.F.T.Y.; Chen, S.; Zhang, Q.; Chen, B.; Ye, Y. Increased nitrous oxide emissions from intertidal soil receiving wastewater from dredging shrimp pond sediments. Environ. Res. Lett. 2020, 15, 094015. [Google Scholar] [CrossRef]
- Krithika, K.; Purjava, R.; Ramesh, R. Fluxes of methane and nitrous oxide from an Indian mangrove. Curr. Sci. 2008, 94, 218–224. [Google Scholar]
- Müller, D.; Bange, H.W.; Warneke, T.; Rixen, T.; Müller, M.; Mujahid, A.; Notholt, J. Nitrous oxide and methane in two tropical estuaries in a peat-dominated region of northwestern Borneo. Biogeosciences 2016, 13, 2415–2428. [Google Scholar] [CrossRef]
- Bange, H.W.; Sim, C.H.; Bastian, D.; Kallert, J.; Kock, A.; Mujahid, A.; Müller, M. Nitrous oxide (N2O) and methane (CH4) in rivers and estuaries of northwestern Borneo. Biogeosciences 2019, 16, 4321–4335. [Google Scholar] [CrossRef]
- Barnes, J.; Ramesh, R.; Purvaja, R.; Rajkumar, A.N.; Sentil Kumar, B.; Krithika, K.; Ravichandran, K.; Uher, G.; Upstill-Goddard, R. Tidal dynamics and rainfall control N2O and CH4 emissions from a pristine mangrove creek. Geophys. Res. Lett. 2006, 33, L15405. [Google Scholar] [CrossRef]
- Maher, D.T.; Sippo, J.Z.; Tait, D.R.; Holloway, C.; Santos, I.R. Pristine mangrove creek waters are a sink of nitrous oxide. Sci. Rep. 2016, 6, 25701. [Google Scholar] [CrossRef] [PubMed]
- Divia, J.I.; Neetha, V.; Hariharan, G.; Kakolee, B.; Purvaja, V.; Ramesh, R. N2O flux from South Andaman mangroves and surrounding creek waters. Int. J. Ocean. Oceanogr. 2013, 7, 73–82. [Google Scholar]
- Murray, R.; Erler, D.; Rosentreter, J.; Maher, D.; Eyre, B. A seasonal source and sink of nitrous oxide in mangroves: Insights from concentration, isotope, and isotopomer measurements. Geochim. Cosmochim. Acta 2018, 238, 169–192. [Google Scholar] [CrossRef]
- Reading, M.J.; Santos, I.R.; Maher, D.T.; Jeffrey, L.C.; Tait, D.R. Shifting nitrous oxide source/sink behaviour in a subtropical estuary revealed by automated time series observations. Estuar. Coast. Shelf Sci. 2017, 194, 66–76. [Google Scholar] [CrossRef]
- Sturm, K.; werner, U.; Grinham, A.; Yuan, Z. tidal variability in methane and nitrous oxide emissions along a subtropical estuarine gradient. Estuar. Coast. Shelf Sci. 2017, 192, 159–169. [Google Scholar] [CrossRef]
- Rao, G.D.; Sarma, V.V.S.S. Contribution of N2O emissions to the atmosphere from Indian monsoonal estuaries. Tellus B 2013, 65, 19660. [Google Scholar] [CrossRef]
- Murray, R.; Erler, D.; Rosentreter, J.; Wells, N.; Eyre, B. Seasonal and spatial controls on N2O concentrations and emissions in low-nitrogen estuaries: Evidence from three tropical systems. Mar. Chem. 2020, 221, 103779. [Google Scholar] [CrossRef]
- Chen, N.; Wu, J.; Zhou, X.; Chen, Z.; Lu, T. Riverine N2O production, emissions and export from a region dominated by agriculture in Southeast Asia (Jiulong River). Agric. Ecosyst. Environ. 2015, 208, 37–47. [Google Scholar] [CrossRef]
- Lin, H.; Dai, M.; Kao, S.J.; Wang, L.; Roberts, E.; Yang, J.Y.T.; Huang, T.; He, B. Spatiotemporal variability of nitrous oxide in a large eutrophic estuarine system: The Pearl River estuary, China. Mar. Chem. 2016, 182, 14–24. [Google Scholar] [CrossRef]
- Liu, X.L.; Bai, L.; Wang, Z.L.; Li, J.; Yue, F.J.; Li, S.L. Nitrous oxide emissions from river network with variable nitrogen loading in Tianjin, China. J. Geochem. Expl. 2015, 157, 153–161. [Google Scholar] [CrossRef]
- Wells, N.S.; Maher, D.T.; Erlet, D.V.; Hipsey, M.; Rosentreter, J.A.; Eyre, B.D. Estuaries as sources and sinks of N2O across a land-use gradient in subtropical Australia. Glob. Biogeochem. Cycle. 2018, 32, 877–894. [Google Scholar] [CrossRef]
- Manjrekar, S.; Uskaikar, H.; Morajkar, S. Seasonal production of nitrous oxide in a tropical estuary off western India. Reg. Stud. Mar. Sci. 2020, 39, 101418. [Google Scholar] [CrossRef]
- Mohammed, S.M.; Johnstone, R.W. Porewater profiles and nutrient sediment-water exchange in a tropical mangrove waterway, Mapopwe Creek, Chwaka Bay, Zanzibar. Afr. J. Ecol. 2002, 40, 172–178. [Google Scholar] [CrossRef]
- Jiang, S.; Müller, M.; Jin, J.; Wy, Y.; Zhu, K.; Zhang, G.; Mujahid, A.; Rixen, T.; Fakharuddin Muhamad, M.; Sien Aun Sia, E.; et al. Dissolved inorganic nitrogen in a tropical estuary in Malaysia: Transport and transformation. Biogeosciences 2019, 16, 2821–2836. [Google Scholar] [CrossRef]
- Terada, K.; Koibuchi, Y.; Isobe, M. Rainfall effect on sediment and nutrient fluxes in a small mangrove river, Okinawa, Japan. J. Mar. Res. 2017, 75, 1–17. [Google Scholar] [CrossRef]
- Davis, S.E., III; Lirman, D.; Wozniak, J.R. Nitrogen and Phosphorus Exchange among Tropical Coastal Ecosystems. In Ecological Connectivity among Tropical Coastal Ecosystems; Nagelkerken, I., Ed.; Springer: Amsterdam, The Netherlands, 2009; pp. 9–43. [Google Scholar]
- Davis, S.E., III; Childers, D.L.; Day, J.W., Jr.; Rudnick, D.T.; Sklar, F.H. Wetland-water column exchanges of carbon, nitrogen, and phosphorus in a Southern Everglades dwarf mangrove. Estuaries 2001, 24, 610–622. [Google Scholar] [CrossRef]
- Boto, K.G.; Bunt, J.S. Tidal export of particulate organic matter from a northern Australian mangrove system. Estuar. Coast. Shelf Sci. 1981, 13, 247–255. [Google Scholar] [CrossRef]
- Rivera-Monroy, V.H.; Day, J.W.; Twilley, R.R.; Vera-Herrera, F.; Coronado-Molina, C. Flux of nitrogen and sediment in a fringe mangrove forest in Terminos Lagoon, Mexico. Estuar. Coast. Shelf Sci. 1995, 40, 139–160. [Google Scholar] [CrossRef]
- Davis, S.E., III; Childers, D.L.; Day, J.W., Jr.; Rudnick, D.T.; Sklar, F.H. Nutrient dynamics in vegetated and unvegetated areas of a Southern Everglades mangrove creek. Estuar. Coast. Shelf Sci. 2001, 52, 753–768. [Google Scholar] [CrossRef]
- Barboza, C.D.N.; Paes, E.T.; de Andrade Jandre, K.; Marques, A.N., Jr. Concentrations and fluxes of nutrients and suspended organic matter in a tropical estuarine system: The Tinharé-Boipeba Islands Archipelago (Baixo Sul Baiano, Brazil). J. Coast. Res. 2014, 30, 1197–1209. [Google Scholar] [CrossRef]
- Tay, H.W.; Bryan, K.R.; Pilditch, C.A.; Park, S.; Hamilton, D.P. Variations in nutrient concentrations at different time scales in two shallow tidally dominated estuaries. Mar. Freshw. Res. 2012, 63, 95–109. [Google Scholar] [CrossRef]
- Valiela, I.; Elmstrom, E.; Lloret, J.; Stone, T.; Camilli, L. Tropical land-sea couplings: Role of watershed deforestation, mangrove estuary processing, and marine inputs on N fluxes in coastal Pacific Panama. Sci. Total Environ. 2018, 630, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Monroy, V.H.; Twilley, R.R.; Davis, S.E., III; Childers, D.L.; Simard, M.; Chambers, R.; Jaffe, R.; Boyer, J.N.; Rudnick, D.T.; Zhang, K.; et al. The role of the Everglades mangrove ecotone region (EMER) in regulating nutrient cycling and wetland productivity in South Florida. Crit. Rev. Environ. Sci. Technol. 2011, 41 (Suppl. S1), 633–669. [Google Scholar] [CrossRef]
- Ray, R.; Majumder, N.; Das, S.; Chowdhury, C.; Kumar Jana, T. Biogeochemical cycle of nitrogen in a tropical mangrove ecosystem, east coast of India. Mar. Chem. 2014, 167, 33–43. [Google Scholar] [CrossRef]
- Adame, M.F.; Virdis, B.; Lovelock, C.E. Effect of geomorphological setting and rainfall on nutrient exchange in mangroves during tidal inundation. Mar. Freshw. Res. 2010, 61, 1197–1206. [Google Scholar] [CrossRef]
- Adame, M.F.; Lovelock, C.E. Carbon and nutrient exchange of mangrove forests with the coastal ocean. Hydrobiologia 2011, 663, 23–50. [Google Scholar] [CrossRef]
- Hamilton, S.E.; Casey, D. Creation of a high spatio-temporal resolution global database of continuous mangrove forest cover for the 21st century. Glob. Ecol. Biogeogr. 2016, 25, 729–738. [Google Scholar] [CrossRef]
- Alongi, D.M. Carbon cycling in the world’s mangrove ecosystems revisited: Significance of non-steady state diagenesis and subsurface linkages between the forest floor and the coastal ocean. Forests 2020, 11, 977. [Google Scholar] [CrossRef]
- Hillebrand, H.; Sommer, U. The nutrient stoichiometry of benthic microalgal growth: Redfield proportions are optimal. Limnol. Oceangr. 1999, 44, 440–446. [Google Scholar] [CrossRef]
- Cerόn, R.M.; Cerόn, J.G.; Muriel, M.; Anguebes, F.; Ramirez, M.; Zavala, J.; Carballo, C.; Escoffie, R.C. Spatial and Temporal Distribution of Throughfall Deposition of Nitrogen and Sulfur in the Mangrove Forests Associated with Terminos Lagoon. In Current Air Quality Issues; Nejadkoorki, F., Ed.; Intech Open Ltd: London, UK, 2015; pp. 147–164. [Google Scholar]
- Seitzinger, S.; Harrison, J.A.; Böhlke, J.K.; Bouwman, A.F.; Lowrance, R.; Peterson, B.; Tobias, C.; Van Drecht, G. Denitrification across landscapes and waterscapes: A synthesis. Ecol. Appl. 2006, 16, 2064–2090. [Google Scholar] [CrossRef]
- Reis, C.R.; Nardoto, G.B.; Oliveira, R.S. Global overview on nitrogen dynamics in mangroves and consequences of increasing nitrogen availability for these systems. Plant Soil 2017, 410, 1–9. [Google Scholar] [CrossRef]
- Alvarez-Cobelas, M.; Angeler, D.G.; Sánchez-Carrillo, S. Export of nitrogen from catchments: A worldwide analysis. Environ. Pollut. 2008, 156, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Beusen, A.H.W.; Bouwman, A.F.; Van Beck, L.P.H.; Mogollόn, J.M.; Middelburg, J.J. Global riverine N and P transport to ocean increased during the 20th century despite increased retention along the aquatic continuum. Biogeosciences 2016, 13, 2441–2451. [Google Scholar] [CrossRef]
- Fowler, D.; Coyle, M.; Skiba, U.; Sutton, M.A.; Cape, J.N.; Reis, S.; Sheppard, L.J.; Jenkins, A.; Grizzetti, B.; Galloway, J.N.; et al. The global nitrogen cycle in the twenty-first century. Philos. Trans. R. Soc. B 2013, 368, 20130164. [Google Scholar] [CrossRef]
- Alongi, D.M. Carbon balance in salt marsh and mangrove ecosystems: A global synthesis. J. Mar. Sci. Eng. 2020, 8, 767. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Number of Observations | Mean ± 1 SE | Median | Range |
|---|---|---|---|---|
| Green leaves (%N) | 109 | 1.61 ± 0.05 | 1.54 | 2.55 |
| Stems (%N) | 58 | 0.37 ± 0.03 | 0.35 | 1.22 |
| Branches (%N) | 55 | 0.31 ± 0.02 | 0.29 | 0.65 |
| Roots (%N) | 63 | 0.49 ± 0.03 | 0.45 | 1.69 |
| Litter (%N) | 153 | 0.69 ± 0.03 | 0.64 | 2.04 |
| Component | Number of Observations | Mean ± 1 SE | Median | Range |
|---|---|---|---|---|
| Soil surface | 57 | 8.22 ± 1.69 | 3.22 | 0–58.92 |
| Cyanobacteria mats | 28 | 9.69 ± 2.62 | 3.43 | 0–60.42 |
| Aboveground roots | 18 | 31.78 ± 5.63 | 26.25 | 2.77–73.4 |
| Belowground roots + rhizomes | 15 | 6.21 ± 1.65 | 4.50 | 1.04–26.20 |
| Litter | 9 | 1.16 ± 0.40 | 0.45 | 0.21–3.30 |
| Senescent leaves | 7 | 0.75 ± 0.13 | 0.83 | 0.39–1.20 |
| Stem bark | 5 | 100.95 ± 36.42 | 100.95 | 17.40–201.20 |
| Transformation Process | Number of Observations | Mean ± 1 SE | Median | Range |
|---|---|---|---|---|
| Denitrification (complete) | 165 | 26.25 ± 3.34 | 3.90 | 0–443.52 |
| Gross ammonification | 17 | 301.63 ± 50.90 | 267.40 | 77.40–898.80 |
| Net Ammonification | 52 | 31.40 ± 6.19 | 15.40 | 0.17–200.00 |
| Gross nitrification | 25 | 15.08 ± 5.81 | 4.74 | 0–141.00 |
| Net nitrification | 37 | 5.80 ± 1.20 | 1.93 | 0–30.80 |
| Anammox | 35 | 22.11 ± 5.49 | 4.35 | 0–99.4 |
| DNRA | 21 | 18.19 ± 6.77 | 4.54 | 0.01–108.64 |
| Gas and Solute Soil Fluxes | Number of Observations | Mean ± 1 SE | Median | Range |
|---|---|---|---|---|
| N2O soil–air flux | 69 | 0.60 ± 0.17 | 0.22 | −2.03–9.01 |
| NO soil–air flux | 2 | 0.05 ± 0.009 | 0.05 | 0.04–0.06 |
| DON soil–water flux | 41 | −18.29 ± 18.14 | 0.00 | −743–19.6 |
| NO2− + NO3− soil–water flux | 66 | −4.46 ± 1.05 | −0.24 | −29.04–3.36 |
| NH4+ soil–water flux | 78 | −1.48 ± 2.20 | −0.16 | −88.7–55.19 |
| Component | Number of Observations | Mean ± 1 SE | Median | Range |
|---|---|---|---|---|
| NH4+ import | 26 | −9.81 ± 4.35 | −1.57 | −100.76–−0.006 |
| NH4+ export | 23 | 8.00 ± 3.63 | 0.39 | 0.03–62.40 |
| NH4+ net exchange | 49 | −1.45 ± 3.12 | −0.01 | −100.76–62.4 |
| DON import | 9 | −4.69 ± 2.17 | −2.87 | −21.84–−0.91 |
| DON export | 10 | 1.45 ± 0.33 | 1.37 | 0.08–3.27 |
| DON net exchange | 19 | −1.46 ± 1.24 | 0.08 | −21.84–3.27 |
| NO2− + NO3− import | 24 | −3.62 ± 1.52 | −0.73 | −28.80–−0.003 |
| NO2− + NO3− export | 27 | 1.84 ± 0.55 | 0.29 | 0.05–11.90 |
| NO2− + NO3− net exchange | 51 | −0.73 ± 0.85 | 0.08 | −28.8–11.90 |
| Inputs | Outputs | ||
|---|---|---|---|
| Component | Gg N a−1 | Component | Gg N a−1 |
| Net primary production | 1250 | Burial | 1239 |
| Wood | 245 | Soil NO release | 1.6 |
| Litter | 466 | Soil N2O release | 19 |
| Roots | 556 | Denitrification/Anammox | 1527 |
| Microalgae | 1250 | Water-air exchange | |
| N2-fixation | N2O | 3.5 | |
| Roots | 196 | Tidal exchange | |
| Soil | 260 | DON | 58 |
| Precipitation | 9 | NO2− + NO3− | 46 |
| Tidal exchange | NH4+ | 253 | |
| DON | 148 | PON | 214 |
| NO2− + NO3− | 114 | ||
| NH4+ | 310 | ||
| Soil-Water exchange | |||
| DON | 577 | ||
| NO2− + NO3− | 141 | ||
| NH4+ | 47 | ||
| Total | 4319 | TOTAL | 3361.1 |
| Inputs − Outputs = 957.9 (Net Gain) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alongi, D.M. Nitrogen Cycling and Mass Balance in the World’s Mangrove Forests. Nitrogen 2020, 1, 167-189. https://doi.org/10.3390/nitrogen1020014
Alongi DM. Nitrogen Cycling and Mass Balance in the World’s Mangrove Forests. Nitrogen. 2020; 1(2):167-189. https://doi.org/10.3390/nitrogen1020014
Chicago/Turabian StyleAlongi, Daniel M. 2020. "Nitrogen Cycling and Mass Balance in the World’s Mangrove Forests" Nitrogen 1, no. 2: 167-189. https://doi.org/10.3390/nitrogen1020014
APA StyleAlongi, D. M. (2020). Nitrogen Cycling and Mass Balance in the World’s Mangrove Forests. Nitrogen, 1(2), 167-189. https://doi.org/10.3390/nitrogen1020014