

Comparison of the PF07598-Encoded Virulence-Modifying Proteins of L. interrogans and L. borgpetersenii

Abstract
1. Introduction
2. Leptospirosis and Vaccination
3. Virulence Factors in Leptospira and PF07598 Paralogous Gene Family
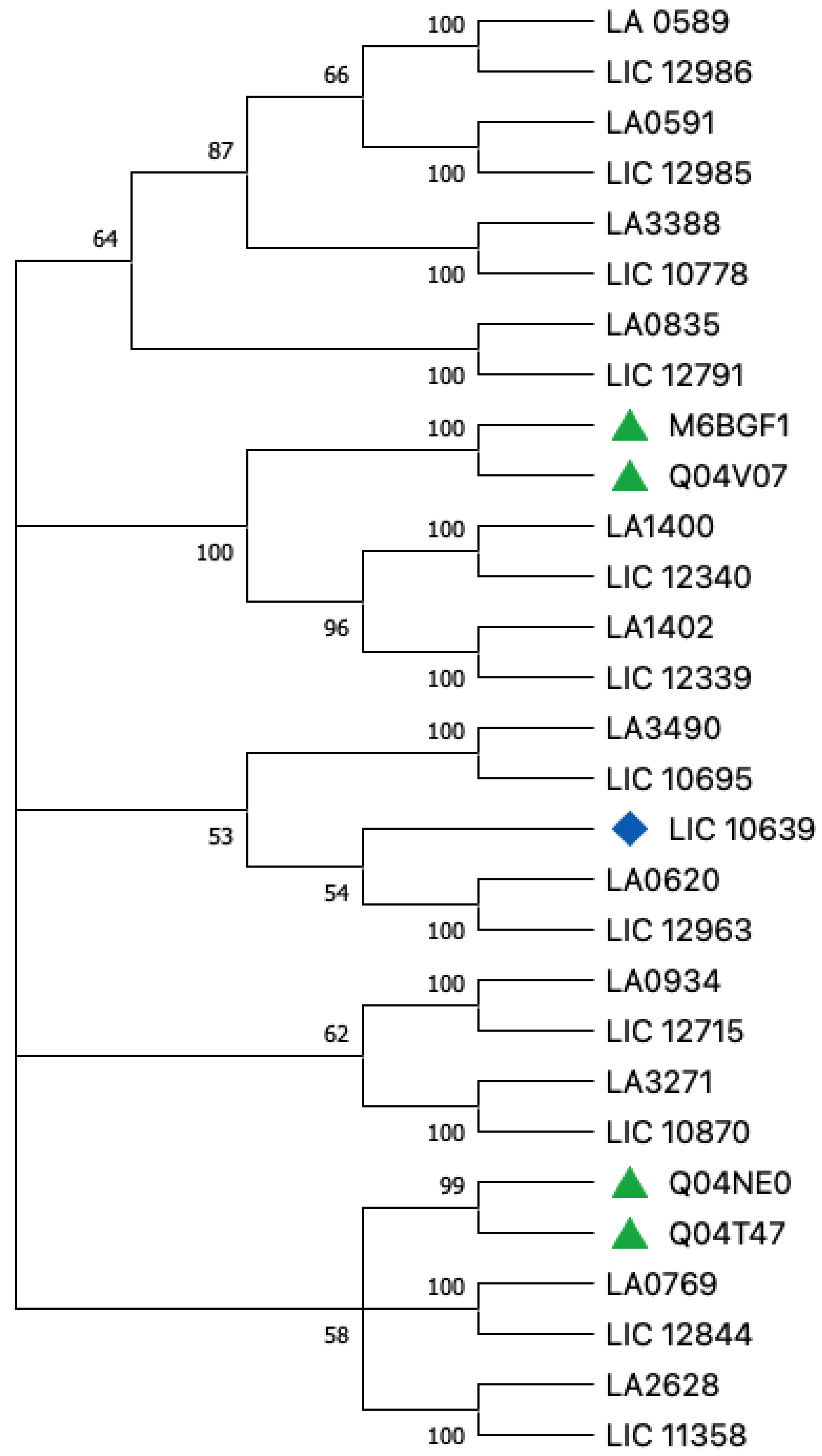
4. Proteins of PF07598 Paralogous Gene Family: General Comparison between Leptospirosis Agents
Host Tropism
5. General Hypothesis
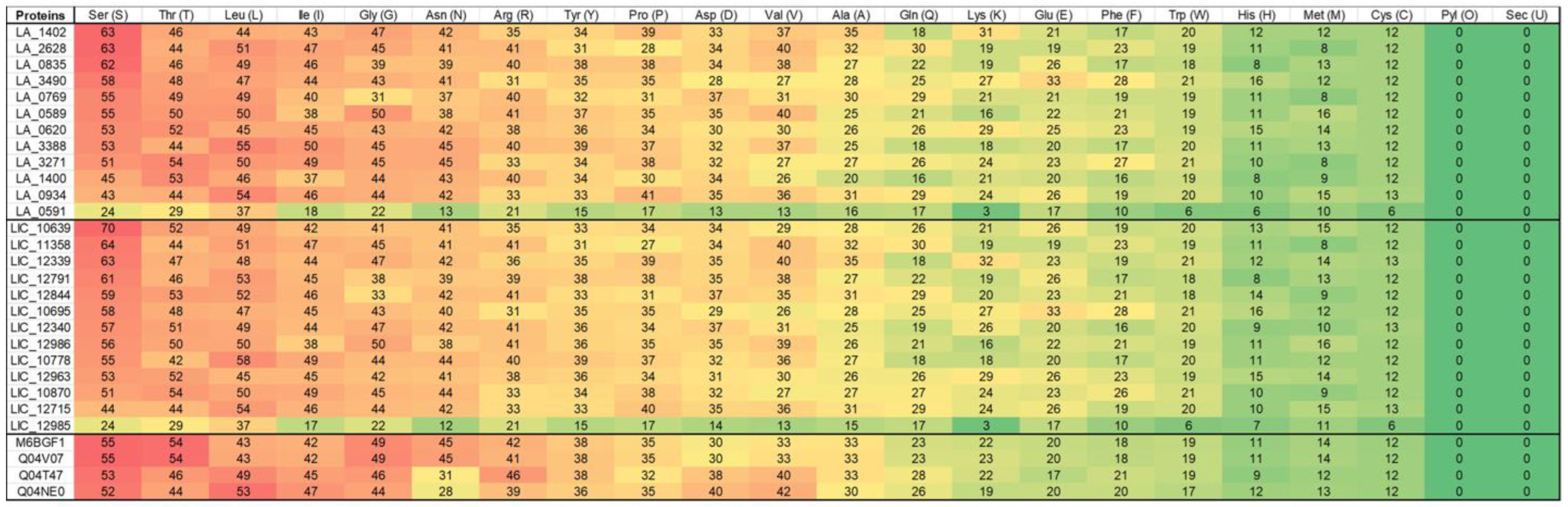
6. Amino Acids and Basic Evolutionary Differences
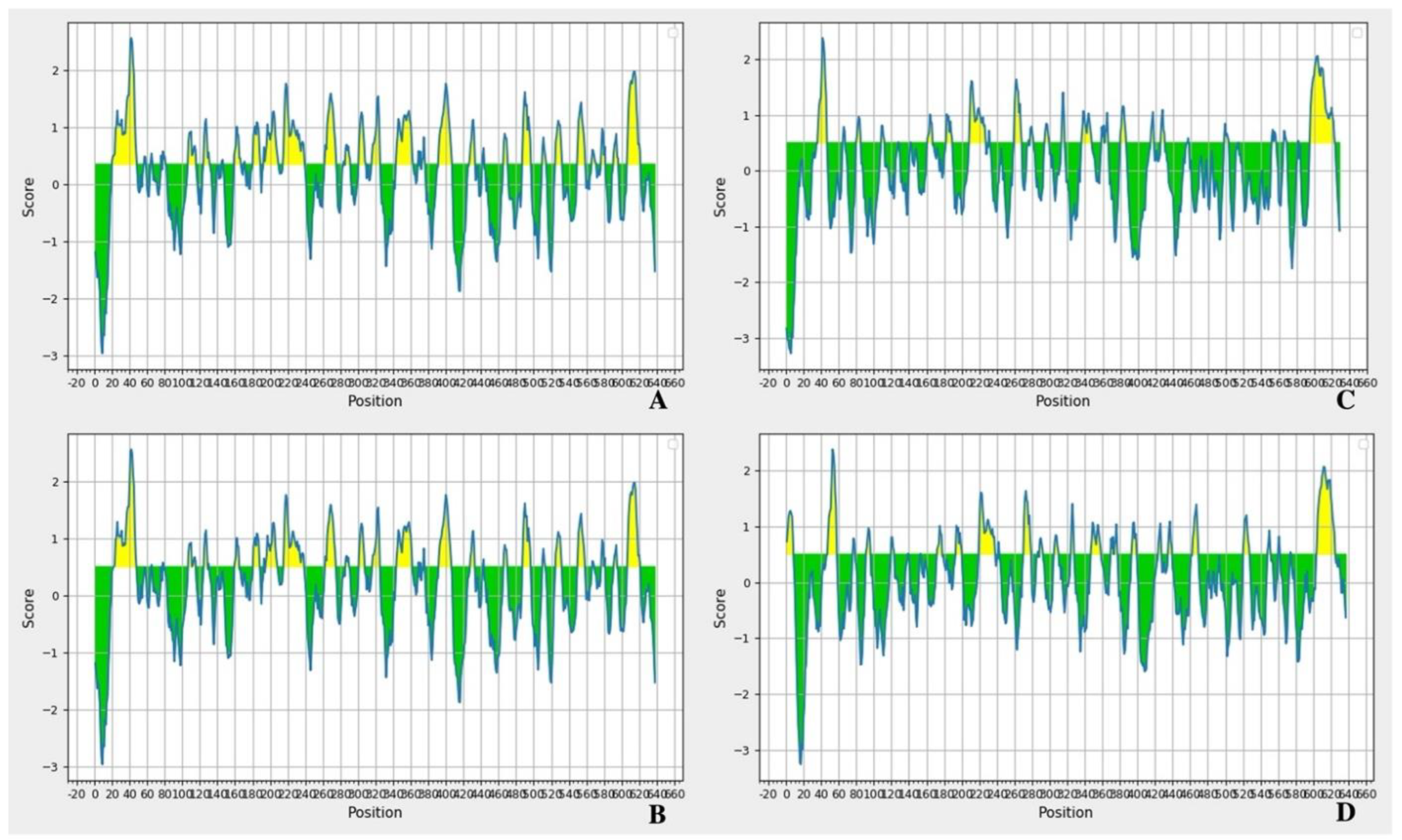
7. Antigenicity and Epitopes
8. Clinical Relevance and Challenges
9. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lopez, A.D.; Mathers, C.D. Measuring the global burden of disease and epidemiological transitions: 2002–2030. Ann. Trop. Med. Parasitol. 2006, 100, 481–499. [Google Scholar] [CrossRef] [PubMed]
- Garba, B.; Bahaman, A.R.; Bejo, S.K.; Zakaria, Z.; Mutalib, A.R.; Bande, F. Major epidemiological factors associated with leptospirosis in Malaysia. Acta Trop. 2018, 178, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.; Hagan, J.E.; Calcagno, J.; Kane, M.; Torgerson, P.; Martinez-Silveira, M.S.; Stein, C.; Abela-Ridder, B.; Ko, A.I. Global Morbidity and Mortality of Leptospirosis: A Systematic Review. PLoS Negl. Trop. Dis. 2015, 9, e0003898. [Google Scholar] [CrossRef] [PubMed]
- Martins, G.; Lilenbaum, W. Control of bovine leptospirosis: Aspects for consideration in a tropical environment. Res. Vet. Sci. 2017, 112, 156–160. [Google Scholar] [CrossRef]
- Picardeau, M. Virulence of the zoonotic agent of leptospirosis: Still terra incognita? Nat. Rev. Microbiol. 2017, 15, 297–307. [Google Scholar] [CrossRef]
- Agampodi, S.B.; Karunarathna, D.; Jayathilala, N.; Rathnayaka, H.; Agampodi, T.C.; Karunanayaka, L. Outbreak of leptospirosis after white-water rafting: Sign of a shift from rural to recreational leptospirosis in Sri Lanka? Epidemiol. Infect. 2014, 142, 843–846. [Google Scholar] [CrossRef]
- Agampodi, S.; Peacock, S.J.; Thevanesam, V. The potential emergence of leptospirosis in Sri Lanka. Lancet Infect. Dis. 2009, 9, 524–526. [Google Scholar] [CrossRef]
- Agampodi, S.B. Spatial epidemiology of leptospirosis in Sri Lanka. Epidemiol. Infect. 2012, 140, 1530–1531. [Google Scholar] [CrossRef][Green Version]
- Agampodi, S.B.; Dahanayaka, N.J.; Bandaranayaka, A.K.; Perera, M.; Priyankara, S.; Weerawansa, P.; Matthias, M.A.; Vinetz, J.M. Regional differences of leptospirosis in Sri Lanka: Observations from a flood-associated outbreak in 2011. PLoS Negl. Trop. Dis. 2014, 8, e2626. [Google Scholar] [CrossRef]
- Agampodi, S.B.; Nugegoda, D.B.; Thevanesam, V. Determinants of leptospirosis in Sri Lanka: Study protocol. BMC Infect. Dis. 2010, 10, 332. [Google Scholar] [CrossRef]
- Agampodi, S.B.; Peacock, S.J.; Thevanesam, V.; Nugegoda, D.B.; Smythe, L.; Thaipadungpanit, J.; Craig, S.B.; Burns, M.A.; Dohnt, M.; Boonsilp, S.; et al. Leptospirosis outbreak in Sri Lanka in 2008: Lessons for assessing the global burden of disease. Am. J. Trop. Med. Hyg. 2011, 85, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Ellis, W.A. Animal leptospirosis. Curr. Top. Microbiol. Immunol. 2015, 387, 99–137. [Google Scholar] [CrossRef] [PubMed]
- Nally, J.E.; Hornsby, R.L.; Alt, D.P.; Bayles, D.; Wilson-Welder, J.H.; Palmquist, D.E.; Bauer, N.E. Isolation and characterization of pathogenic leptospires associated with cattle. Vet. Microbiol. 2018, 218, 25–30. [Google Scholar] [CrossRef]
- Stringfellow, D.A.; Brown, R.R.; Hanson, L.E.; Schnurrenberger, P.R.; Johnson, J. Can antibody responses in cattle vaccinated with a multivalent leptospiral bacterin interfere with serologic diagnosis of disease? J. Am. Vet. Med. Assoc. 1983, 182, 165–167. [Google Scholar] [PubMed]
- Draghi, M.G.; Brihuega, B.; Benitez, D.; Sala, J.M.; Biotti, G.M.; Pereyra, M.; Homse, A.; Guariniello, L. Leptospirosis outbreak in calves from Corrientes Province, Argentina. Rev. Argent. Microbiol. 2011, 43, 42–44. [Google Scholar] [CrossRef]
- Fouts, D.E.; Matthias, M.A.; Adhikarla, H.; Adler, B.; Amorim-Santos, L.; Berg, D.E.; Bulach, D.; Buschiazzo, A.; Chang, Y.F.; Galloway, R.L.; et al. What Makes a Bacterial Species Pathogenic?:Comparative Genomic Analysis of the Genus Leptospira. PLoS Negl. Trop. Dis. 2016, 10, e0004403. [Google Scholar] [CrossRef]
- Lehmann, J.S.; Matthias, M.A.; Vinetz, J.M.; Fouts, D.E. Leptospiral pathogenomics. Pathogens 2014, 3, 280–308. [Google Scholar] [CrossRef]
- Lehmann, J.S.; Fouts, D.E.; Haft, D.H.; Cannella, A.P.; Ricaldi, J.N.; Brinkac, L.; Harkins, D.; Durkin, S.; Sanka, R.; Sutton, G.; et al. Pathogenomic inference of virulence-associated genes in Leptospira interrogans. PLoS Negl. Trop. Dis. 2013, 7, e2468. [Google Scholar] [CrossRef]
- Grassmann, A.A.; Souza, J.D.; McBride, A.J. A Universal Vaccine against Leptospirosis: Are We Going in the Right Direction? Front. Immunol. 2017, 8, 256. [Google Scholar] [CrossRef]
- Chen, X.; Xie, X.; Wu, D.; Zhang, S.; Zhang, W.; Cao, Y. The pre-activated immune response induced by LPS protects host from leptospirosis. PLoS ONE 2020, 15, e0242742. [Google Scholar] [CrossRef]
- Bonhomme, D.; Santecchia, I.; Vernel-Pauillac, F.; Caroff, M.; Germon, P.; Murray, G.; Adler, B.; Boneca, I.G.; Werts, C. Correction: Leptospiral LPS escapes mouse TLR4 internalization and TRIF-associated antimicrobial responses through O antigen and associated lipoproteins. PLoS Pathog. 2020, 16, e1009173. [Google Scholar] [CrossRef] [PubMed]
- Werts, C.; Tapping, R.I.; Mathison, J.C.; Chuang, T.H.; Kravchenko, V.; Saint Girons, I.; Haake, D.A.; Godowski, P.J.; Hayashi, F.; Ozinsky, A.; et al. Leptospiral lipopolysaccharide activates cells through a TLR2-dependent mechanism. Nat. Immunol. 2001, 2, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Bergmann Esteves, S.; Moreira Santos, C.; Ferreira Salgado, F.; Paldes Goncales, A.; Gil Alves Guilloux, A.; Marinelli Martins, C.; Kuribaiashi Hagiwara, M.; Alonso Miotto, B. Efficacy of commercially available vaccines against canine leptospirosis: A systematic review and meta-analysis. Vaccine 2022, 40, 1722–1740. [Google Scholar] [CrossRef] [PubMed]
- Dellagostin, O.A.; Grassmann, A.A.; Hartwig, D.D.; Felix, S.R.; da Silva, E.F.; McBride, A.J. Recombinant vaccines against leptospirosis. Hum. Vaccin. 2011, 7, 1215–1224. [Google Scholar] [CrossRef] [PubMed]
- Murray, G.L.; Lo, M.; Bulach, D.M.; Srikram, A.; Seemann, T.; Quinsey, N.S.; Sermswan, R.W.; Allen, A.; Adler, B. Evaluation of 238 antigens of Leptospira borgpetersenii serovar Hardjo for protection against kidney colonisation. Vaccine 2013, 31, 495–499. [Google Scholar] [CrossRef]
- Sant’Anna da Costa, R.; Di Azevedo, M.I.N.; Dos Santos Baptista Borges, A.L.; Aymee, L.; Martins, G.; Lilenbaum, W. Effect of Vaccination against Leptospira on Shelter Asymptomatic Dogs Following a Long-Term Study. Animals 2022, 12, 1788. [Google Scholar] [CrossRef]
- Smith, A.M.; Stull, J.W.; Moore, G.E. Potential Drivers for the Re-Emergence of Canine Leptospirosis in the United States and Canada. Trop. Med. Infect. Dis. 2022, 7, 377. [Google Scholar] [CrossRef]
- Eric Klaasen, H.L.; Adler, B. Recent advances in canine leptospirosis: Focus on vaccine development. Vet. Med. 2015, 6, 245–260. [Google Scholar] [CrossRef]
- Wilson-Welder, J.H.; Alt, D.P.; Nally, J.E.; Olsen, S.C. Bovine Immune Response to Vaccination and Infection with Leptospira borgpetersenii Serovar Hardjo. mSphere 2021, 6, e00988-20. [Google Scholar] [CrossRef]
- Dreyfus, A.; Wilson, P.; Collins-Emerson, J.; Benschop, J.; Moore, S.; Heuer, C. Risk factors for new infection with Leptospira in meat workers in New Zealand. Occup. Environ. Med. 2015, 72, 219–225. [Google Scholar] [CrossRef]
- Bolin, C.A.; Cassells, J.A.; Zuerner, R.L.; Trueba, G. Effect of vaccination with a monovalent Leptospira interrogans serovar hardjo type hardjo-bovis vaccine on type hardjo-bovis infection of cattle. Am. J. Vet. Res. 1991, 52, 1639–1643. [Google Scholar] [PubMed]
- Chaurasia, R.; Salovey, A.; Guo, X.; Desir, G.; Vinetz, J.M. Vaccination with Leptospira interrogans PF07598 gene family-encoded Virulence Modifying Proteins protects mice from severe leptospirosis and reduces bacterial load in liver and kidney. Front. Cell. Infect. Microbiol. 2022, 28, 822. [Google Scholar] [CrossRef] [PubMed]
- Putz, E.J.; Fernandes, L.G.V.; Sivasankaran, S.K.; Bayles, D.O.; Alt, D.P.; Lippolis, J.D.; Nally, J.E. Some like it hot, some like it cold; proteome comparison of Leptospira borgpetersenii serovar Hardjo strains propagated at different temperatures. J. Proteom. 2022, 262, 104602. [Google Scholar] [CrossRef] [PubMed]
- Maia, M.A.C.; Bettin, E.B.; Barbosa, L.N.; de Oliveira, N.R.; Bunde, T.T.; Pedra, A.C.K.; Rosa, G.A.; da Rosa, E.E.B.; Seixas Neto, A.C.P.; Grassmann, A.A.; et al. Challenges for the development of a universal vaccine against leptospirosis revealed by the evaluation of 22 vaccine candidates. Front. Cell. Infect. Microbiol. 2022, 12, 940966. [Google Scholar] [CrossRef] [PubMed]
- Wunder, E.A.; Adhikarla, H.; Hamond, C.; Owers Bonner, K.A.; Liang, L.; Rodrigues, C.B.; Bisht, V.; Nally, J.E.; Alt, D.P.; Reis, M.G.; et al. A live attenuated-vaccine model confers cross-protective immunity against different species of the Leptospira genus. eLife 2021, 10, e64166. [Google Scholar] [CrossRef]
- Chaurasia, R.; Marroquin, A.S.; Vinetz, J.M.; Matthias, M.A. Pathogenic Leptospira Evolved a Unique Gene Family Comprised of Ricin B-Like Lectin Domain-Containing Cytotoxins. Front. Microbiol. 2022, 13, 859680. [Google Scholar] [CrossRef]
- Brenner, D.J.; Kaufmann, A.F.; Sulzer, K.R.; Steigerwalt, A.G.; Rogers, F.C.; Weyant, R.S. Further determination of DNA relatedness between serogroups and serovars in the family Leptospiraceae with a proposal for Leptospira alexanderi sp. nov. and four new Leptospira genomospecies. Int. J. Syst. Bacteriol. 1999, 49 Pt 2, 839–858. [Google Scholar] [CrossRef]
- Bulach, D.M.; Zuerner, R.L.; Wilson, P.; Seemann, T.; McGrath, A.; Cullen, P.A.; Davis, J.; Johnson, M.; Kuczek, E.; Alt, D.P.; et al. Genome reduction in Leptospira borgpetersenii reflects limited transmission potential. Proc. Natl. Acad. Sci. USA 2006, 103, 14560–14565. [Google Scholar] [CrossRef]
- Vincent, A.T.; Schiettekatte, O.; Goarant, C.; Neela, V.K.; Bernet, E.; Thibeaux, R.; Ismail, N.; Mohd Khalid, M.K.N.; Amran, F.; Masuzawa, T.; et al. Revisiting the taxonomy and evolution of pathogenicity of the genus Leptospira through the prism of genomics. PLoS Negl. Trop. Dis. 2019, 13, e0007270. [Google Scholar] [CrossRef]
- Baumler, A.; Fang, F.C. Host specificity of bacterial pathogens. Cold Spring Harb. Perspect. Med. 2013, 3, a010041. [Google Scholar] [CrossRef]
- Ricaldi, J.N.; Fouts, D.E.; Selengut, J.D.; Harkins, D.M.; Patra, K.P.; Moreno, A.; Lehmann, J.S.; Purushe, J.; Sanka, R.; Torres, M.; et al. Whole Genome Analysis of Leptospira licerasiae Provides Insight into Leptospiral Evolution and Pathogenicity. PLoS Negl. Trop. Dis. 2012, 6, e1853. [Google Scholar] [CrossRef] [PubMed]
- Gunnell, M.K.; Robison, R.A.; Adams, B.J. Natural Selection in Virulence Genes of Francisella tularensis. J. Mol. Evol. 2016, 82, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Putz, E.J.; Sivasankaran, S.K.; Fernandes, L.G.V.; Brunelle, B.; Lippolis, J.D.; Alt, D.P.; Bayles, D.O.; Hornsby, R.L.; Nally, J.E. Distinct transcriptional profiles of Leptospira borgpetersenii serovar Hardjo strains JB197 and HB203 cultured at different temperatures. PLoS Negl. Trop. Dis. 2021, 15, e0009320. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Sruthi, C.K.; Prakash, M. Amino acid impact factor. PLoS ONE 2018, 13, e0198645. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Yin, R.; Kwoh, C.K.; Zheng, J. A context-free encoding scheme of protein sequences for predicting antigenicity of diverse influenza A viruses. BMC Genom. 2018, 19, 936. [Google Scholar] [CrossRef]
- Anfinsen, C.B. Principles that govern the folding of protein chains. Science 1973, 181, 223–230. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. In 2-D Proteome Analysis Protocols; Link, A.J., Ed.; Methods in Molecular Biology; Humana Press: Clifton, NJ, USA, 1999; Volume 112, pp. 531–552. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Grassmann, A.A.; Kremer, F.S.; Dos Santos, J.C.; Souza, J.D.; Pinto, L.D.S.; McBride, A.J.A. Discovery of Novel Leptospirosis Vaccine Candidates Using Reverse and Structural Vaccinology. Front. Immunol. 2017, 8, 463. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Wang, D.; Hu, N.; Zhu, Q.; Chen, K.; Dong, K.; Zhang, Y.; Yao, Y.; Guo, X.; Chang, Y.F.; et al. A Novel Pan-Genome Reverse Vaccinology Approach Employing a Negative-Selection Strategy for Screening Surface-Exposed Antigens against leptospirosis. Front. Microbiol. 2017, 8, 396. [Google Scholar] [CrossRef] [PubMed]
- Dellagostin, O.A.; Grassmann, A.A.; Rizzi, C.; Schuch, R.A.; Jorge, S.; Oliveira, T.L.; McBride, A.J.; Hartwig, D.D. Reverse Vaccinology: An Approach for Identifying Leptospiral Vaccine Candidates. Int. J. Mol. Sci. 2017, 18, 158. [Google Scholar] [CrossRef] [PubMed]
- Doytchinova, I.A.; Flower, D.R. Identifying candidate subunit vaccines using an alignment-independent method based on principal amino acid properties. Vaccine 2007, 25, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Trincado, J.L.; Gomez-Perosanz, M.; Reche, P.A. Fundamentals and Methods for T- and B-Cell Epitope Prediction. J. Immunol. Res. 2017, 2017, 2680160. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lepstospira Strain | Gene | Protein ID | Amino Acids | Cleavage Site |
|---|---|---|---|---|
| L. interrogans Lai | LA_3490 | Q8F0K3 | 639 | 18aa–19aa |
| LA_0620 | Q8F8D7 | 637 | 29aa–30aa | |
| LA_1402 | Q8F6A7 | 641 | 27aa–28aa | |
| LA_1400 | Q8F6A9 | 573 | NA | |
| LA_0591 | Q8F8G6 | 313 | 22aa–23aa | |
| LA_3388 | Q8F0V3 | 631 | 22aa–23aa | |
| LA_0835 | Q8F7V7 | 631 | 22aa–23aa | |
| LA_0589 | Q8F8G8 | 632 | 22aa–23aa | |
| LA_3271 | Q8F166 | 636 | 31aa–32aa | |
| LA_0934 | Q8F7L0 | 638 | 31aa–32aa, | |
| LA_0769 | Q8F820 | 602 | NA | |
| LA_2628 | Q8F2Y3 | 638 | 22aa–23aa | |
| L. borgpeterseni | LBJ_1339 | Q04T47 | 637 | 30aa–31aa |
| LBJ_0577 | Q04V07 | 638 | 22aa–23aa | |
| LBJ_4195 | Q04NE0 | 629 | 19aa–20aa | |
| LEP1GSC016_2732 | M6BGF1 | 638 | 22aa–23aa | |
| L. interrogans Copenhageni | LIC_12986 | Q72N52 | 632 | 22aa–23aa |
| LIC_12339 | Q72PX8 | 663 | NA | |
| LIC_12340 | Q72PX7 | 627 | 30aa–31aa | |
| LIC_10695 | Q72UG2 | 639 | 30aa–31aa | |
| LIC_10639 | Q72UL8 | 640 | 33aa–34aa | |
| LIC_12963 | Q72N74 | 637 | 31aa–32aa | |
| LIC_12844 | Q72NJ0 | 639 | 24aa–25aa | |
| LIC_10870 | Q72TZ4 | 636 | 31aa–32aa | |
| LIC_12715 | Q72NW3 | 638 | 31aa–32aa | |
| LIC_11358 | 72SM1 | 638 | 22aa–23aa | |
| LIC_12791 | Q72NP1 | 631 | 22aa–23aa | |
| LIC_10778 | Q72U83 | 631 | 22aa–23aa | |
| LIC_12985 | Q72U83 | 631 | 22aa–23aa |
| Protein | Antigenicity | Prediction for the Protective Antigen |
|---|---|---|
| LA1400 | + | 0.4561 |
| LA1402 | + | 0.3814 |
| M6BGE1 | + | 0.3925 |
| Q04V07 | + | 0.3998 |
| Q04NE0 | + | 0.3928 |
| Q04T47 | + | 0.4243 |
| Taxon Id | Scientific Name |
|---|---|
| 1303729 * | Leptospira borgpetersenii serovar Hardjo-bovis str. Sponselee |
| 280504 | Leptospira borgpetersenii serovar Javanica |
| 280505 | Leptospira borgpetersenii serovar Ballum |
| 280506 | Leptospira borgpetersenii serovar Tarassovi |
| 280507 | Leptospira borgpetersenii serovar Mini |
| 328971 | Leptospira borgpetersenii serovar Hardjo |
| 338217 | Leptospira borgpetersenii serovar Hardjo-bovis |
| 338220 | Leptospira borgpetersenii serovar Sejroe |
| 355277 * | Leptospira borgpetersenii serovar Hardjo-bovis str. JB197 |
| 376921 | Leptospira borgpetersenii serovar Castellonis |
| 400680 | Leptospira borgpetersenii serovar Saxkoebing |
| 400681 | Leptospira borgpetersenii serovar Wolfii |
| 508535 | Leptospira borgpetersenii serovar Balcanica |
| 508536 | Leptospira borgpetersenii serovar Ceylonica |
| 508537 | Leptospira borgpetersenii serovar Dikkeni |
| 508538 | Leptospira borgpetersenii serovar Jules |
| 508539 | Leptospira borgpetersenii serovar Kisuba |
| 508540 | Leptospira borgpetersenii serovar Kwale |
| 508541 | Leptospira borgpetersenii serovar Moldaviae |
| 508542 | Leptospira borgpetersenii serovar Nero |
| 508543 | Leptospira borgpetersenii serovar Nyanza |
| 508544 | Leptospira borgpetersenii serovar Pina |
| 508545 | Leptospira borgpetersenii serovar Poi |
| 508546 | Leptospira borgpetersenii serovar Polonica |
| 508547 | Leptospira borgpetersenii serovar Sorexjalna |
| 508548 | Leptospira borgpetersenii serovar Tunis |
| 508549 | Leptospira borgpetersenii serovar Worsfoldi |
| 508550 | Leptospira borgpetersenii serovar Srebarna |
| 561995 | Leptospira borgpetersenii serovar Whitticombi |
| 577404 | Leptospira borgpetersenii serovar Istrica |
| 652580 | Leptospira borgpetersenii serovar Arborea |
| 1049771 | Leptospira borgpetersenii serovar Kenya |
| 1049780 | Leptospira borgpetersenii str. Brem 328 |
| 1049781 | Leptospira borgpetersenii str. Noumea 25 |
| 1049782 | Leptospira borgpetersenii str. UI 09149 |
| 1049786 | Leptospira borgpetersenii str. Brem 307 |
| 1141102 | Leptospira borgpetersenii str. 4E |
| 1192865 | Leptospira borgpetersenii serovar Pomona |
| 1193007 | Leptospira borgpetersenii str. 200701203 |
| 1193008 | Leptospira borgpetersenii str. 200801773 |
| 1193009 | Leptospira borgpetersenii str. 200801926 |
| 2034701 | Leptospira borgpetersenii serovar Piyasena |
| 2814707 | Leptospira borgpetersenii serovar Guangdong |
| 2814708 | Leptospira borgpetersenii serovar Hamptoni |
| 2814709 | Leptospira borgpetersenii serovar Nigeria |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieira, D.S.; Chaurasia, R.; Vinetz, J.M. Comparison of the PF07598-Encoded Virulence-Modifying Proteins of L. interrogans and L. borgpetersenii. Trop. Med. Infect. Dis. 2023, 8, 14. https://doi.org/10.3390/tropicalmed8010014
Vieira DS, Chaurasia R, Vinetz JM. Comparison of the PF07598-Encoded Virulence-Modifying Proteins of L. interrogans and L. borgpetersenii. Tropical Medicine and Infectious Disease. 2023; 8(1):14. https://doi.org/10.3390/tropicalmed8010014
Chicago/Turabian StyleVieira, Dielson S., Reetika Chaurasia, and Joseph M. Vinetz. 2023. "Comparison of the PF07598-Encoded Virulence-Modifying Proteins of L. interrogans and L. borgpetersenii" Tropical Medicine and Infectious Disease 8, no. 1: 14. https://doi.org/10.3390/tropicalmed8010014
APA StyleVieira, D. S., Chaurasia, R., & Vinetz, J. M. (2023). Comparison of the PF07598-Encoded Virulence-Modifying Proteins of L. interrogans and L. borgpetersenii. Tropical Medicine and Infectious Disease, 8(1), 14. https://doi.org/10.3390/tropicalmed8010014