


Evaluation of Methods for Collecting Mosquitoes (Culicidae: Diptera) in Canopy and Ground Strata in the Brazilian Savanna

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Research Area
2.2. Capture of Specimens
2.2.1. Capture by Hand-Net
2.2.2. Capture with Automatic Traps
2.3. Identification of Material
2.4. Data Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Consoli, R.A.G.B.; de Oliveira, R.L. Principais Mosquitos de Importância Sanitária no Brasil; Editora FIOCRUZ: Rio de Janeiro, Brazil, 1994; ISBN 978-85-85676-03-2. Available online: https://static.scielo.org/scielobooks/th/pdf/consoli-9788575412909.pdf (accessed on 1 July 2022).
- Forattini, O.P. Culicidologia Médica: Identificação, Biologia, Epidemiologia; Edusp: São Paulo, Brasil, 2002; ISBN 13: 9788531406997. [Google Scholar]
- Pinheiro, F.P.; da Rosa, A.P.A.T.; Moraes, M.A.P. An epidemic of yellow fever in central Brazil, 1972–1973: II. Ecological studies. Am. J. Trop. Med. Hyg. 1981, 30, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, P.F.C.; da Rosa, A.P.A.T.; Dégalier, N.; da Rosa, J.F.S.T.; Pinheiro, F.P. Clinical and Ecoepidemiological Situation of Human Arboviruses in Brazilian Amazonia. Ciência e Cultura 1992, 44, 117–124. Available online: http://memoria.bn.br/docreader/DocReader.aspx?bib=003069&pagfis=59549 (accessed on 1 July 2022).
- Iversson, L.B. Current status of the eco-epidemiological knowledge on arboviruses pathogenic to humans in the Atlantic Forest region of the State of São Paulo. Rev. Inst. Med. Trop. Sao Paulo 1994, 36, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, P.F.; Rodrigues, S.G.; Degallier, N.; Moraes, M.A.; da Rosa, J.F.; da Rosa, E.S.; Mondet, B.; Barros, V.L.; da Rosa, A.P. An epidemic of sylvatic yellow fever in the southeast region of Maranhao state, Brazil, 1993–1994: Epidemiologic and entomologic findings. Am. J. Trop. Med. Hyg. 1997, 57, 132–137. [Google Scholar] [CrossRef]
- Rosa, A.P.D.A.T.D. A História a Arbovirologia No Instituto Evandro Chagas, Belém, Pará, Brasil, de 1954 a 1998. Rev. Pan Amaz. Saúde 2016, 7, 61–70. [Google Scholar] [CrossRef]
- Dégallier, N.; Rosa, A.P.D.A.T.D.; Vasconcelos, P.F.D.C.; Rosa, E.S.T.D.; Rodrigues, S.G.; Sá Filho, G.C.; Rosa, J.F.S.T.D. New entomological and virological data on the vectors of sylvatic yellow fever in Brazil. Arthropod-Borne Virus Inf Exch 1993, 21–22. Available online: https://patuaback.iec.gov.br/server/api/core/bitstreams/63d9d538-fdf9-4b50-949a-c3cff5c8a0e1/content. (accessed on 1 July 2022).
- Vasconcelos, P.F.; Rosa, A.P.; Rodrigues, S.G.; Rosa, E.S.; Monteiro, H.A.; Cruz, A.C.; Barros, V.L.; Souza, M.R.; Rosa, J.F. Yellow fever in Pará state, Amazon region of Brazil, 1998–1999: Entomologic and epidemiologic findings. Emerg. Infect. Dis. 2001, 7, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Brasil, M.d.S. Guia de Vigilância de Epizootias em Primatas Não Humanos e Entomologia Aplicada à Vigilância Da Febre Amarela, 2nd ed.; Ministério da Saúde, Secretaria de Vigilância em Saúde, Departamento de Vigilância das Doenças Transmissíveis: Brasília (DF), Brasil, 2014; ISBN 978-85-334-2102-8. Available online: https://bvsms.saude.gov.br/bvs/publicacoes/guia_vigilancia_epizootias_primatas_entomologia.pdf (accessed on 1 July 2022).
- Reiter, P.; Gubler, D. Surveillance and Control of Urban Dengue Vectors. In Dengue and Dengue Hemorrhagic Fever; Gubler, D.J., Kuno, G., Eds.; CAB International: London, UK, 1997; pp. 425–462. [Google Scholar]
- Lima, J.B.P.; Rosa-Freitas, M.G.; Rodovalho, C.M.; Santos, F.; Lourenço-de-Oliveira, R. Is there an efficient trap or collection method for sampling Anopheles darlingi and other malaria vectors that can describe the essential parameters affecting transmission dynamics as effectively as human landing catches?—A review. Mem. Inst. Oswaldo Cruz 2014, 109, 685–705. [Google Scholar] [CrossRef]
- Achee, N.L.; Youngblood, L.; Bangs, M.J.; Lavery, J.V.; James, S. Considerations for the use of human participants in vector biology research: A tool for investigators and regulators. Vector Borne Zoonotic Dis. 2015, 15, 89–102. [Google Scholar] [CrossRef]
- Service, M.W. Mosquito Ecology: Field Sampling Methods; Springer Science & Business Media: Berlin, Germany, 1993; ISBN 978-94-011-1868-2. [Google Scholar]
- Santos, J.C.; de Almeida, W.R.; Fernandes, G.W. Arthropods: Why It Is So Crucial To Know Their Biodiversity? In Measuring Arthropod Biodiversity: A Handbook of Sampling Methods; Santos, J.C., Fernandes, G.W., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 3–11. ISBN 978-3-030-53226-0. [Google Scholar]
- Service, M.W. A critical review of procedures for sampling populations of adult mosquitoes. Bull. Entomol. Res. 1977, 67, 343–382. [Google Scholar] [CrossRef]
- Laporta, G.Z.; Sallum, M.A.M. Effect of CO2 and 1-Octen-3-Ol Attractants for estimating species richness and the abundance of diurnal mosquitoes in the southeastern atlantic forest, Brazil. Mem. Inst. Oswaldo Cruz 2011, 106, 279–284. [Google Scholar] [CrossRef]
- Barrio-Nuevo, K.M.; Cunha, M.S.; Luchs, A.; Fernandes, A.; Rocco, I.M.; Mucci, L.F.; de Souza, R.P.; Medeiros-Sousa, A.R.; Ceretti-Junior, W.; Marrelli, M.T. Detection of zika and dengue viruses in wild-caught mosquitoes collected during field surveillance in an environmental protection area in São Paulo, Brazil. PLoS ONE 2020, 15, e0227239. [Google Scholar] [CrossRef] [PubMed]
- Hendy, A.; Hernandez-Acosta, E.; Valério, D.; Mendonça, C.; Costa, E.R.; Júnior, J.T.A.; Assunção, F.P.; Scarpassa, V.M.; Gordo, M.; Fé, N.F.; et al. The vertical stratification of potential bridge vectors of mosquito-borne viruses in a central amazonian forest bordering Manaus, Brazil. Sci. Rep. 2020, 10, 18254. [Google Scholar] [CrossRef] [PubMed]
- Alencar, J.; Melandri, V.; Silva, J.; Albuquerque, H.G.; Guimarães, A.E. Ecological characterization of mosquitoes (Diptera: Culicidae) in areas of the mato grosso pantanal, Mato Grosso State, Brazil. Trop. Zool. 2021, 34, 1–15. [Google Scholar] [CrossRef]
- de Deus, J.T.; Mucci, L.F.; Reginatto, S.L.; Pereira, M.; Bergo, E.S.; de Camargo-Neves, V.L.F. Evaluation of methods to collect diurnal culicidae (diptera) at canopy and ground strata, in the atlantic forest biome. Insects 2022, 13, 202. [Google Scholar] [CrossRef]
- Gama, R.A.; Da Silva, I.M.; Monteiro, H.A.D.O.; Eiras, A.E. Fauna of culicidae in rural areas of porto velho and the first record of Mansonia (Mansonia) flaveola (Coquillet, 1906), for the state of Rondônia, Brazil. Rev. Soc. Bras. Med. Trop. 2012, 45, 125–127. [Google Scholar] [CrossRef]
- de Sá, I.L.R.; Sallum, M.A.M. Comparison of automatic traps to capture mosquitoes (Diptera: Culicidae) in rural areas in the tropical Atlantic Rainforest. Mem. Inst. Oswaldo Cruz 2013, 108, 1014–1020. [Google Scholar] [CrossRef]
- Chaves, L.S.M.; Laporta, G.Z.; Sallum, M.A.M. Effectiveness of mosquito magnet in preserved area on the coastal atlantic rainforest: Implication for entomological surveillance. J. Med. Entomol. 2014, 51, 915–924. [Google Scholar] [CrossRef]
- Sant’Ana, D.C.; de Sa, I.L.R.; Sallum, M.A.M. Effectiveness of mosquito magnet® trap in rural areas in the southeastern tropical Atlantic forest. Mem. Inst. Oswaldo Cruz 2014, 109, 1021–1029. [Google Scholar] [CrossRef][Green Version]
- Pinheiro, G.G.; Rocha, M.N.; de Oliveira, M.A.; Moreira, L.A.; Filho, J.D.A. Detection of yellow fever virus in sylvatic mosquitoes during disease outbreaks of 2017–2018 in Minas Gerais state, Brazil. Insects 2019, 10, 136. [Google Scholar] [CrossRef]
- Kröckel, U.; Rose, A.; Eiras, A.E.; Geier, M. New Tools for surveillance of adult yellow fever mosquitoes: Comparison of trap catches with human landing rates in an urban environment. J. Am. Mosq. Control. Assoc. 2006, 22, 229–238. [Google Scholar] [CrossRef]
- de Ázara, T.M.F.; Degener, C.M.; Roque, R.A.; Ohly, J.J.; Geier, M.; Eiras, Á.E. The impact of CO2 on collection of Aedes aegypti (Linnaeus) and Culex quinquefasciatus Say by BG-Sentinel(r) traps in Manaus, Brazil. Mem. Inst. Oswaldo Cruz 2013, 108, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Degener, C.M.; de Ázara, T.M.F.; Roque, R.A.; Codeço, C.T.; Nobre, A.A.; Ohly, J.J.; Geier, M.; Eiras, Á.E. Temporal Abundance of Aedes aegypti in Manaus, Brazil, Measured by Two Trap Types for Adult Mosquitoes. Mem. Inst. Oswaldo Cruz 2014, 109, 1030–1040. Available online: http://www.bioline.org.br/pdf?oc14155 (accessed on 1 July 2022). [CrossRef] [PubMed]
- São Paulo, S. Secretaria do Meio Ambiente do Estado de São Paulo Plano de Manejo pe Vassununga 2009. Secr. Meio. Ambiente. Estado. São Paulo. 2009, 309. Available online: http://arquivos.ambiente.sp.gov.br/consema/2011/11/oficio_consema_2011_092/Plano_de_Manejo.pdf (accessed on 1 July 2022).
- Pivello, V.R.; Bitencourt, M.D.; Mantovani, W.; Junior, N.D.M.; Batalha, M.A.; Shida, C.N. Proposta de Zoneamento Ecológico para a Reserva de Cerrado Pé-de-Gigante (Santa Rita do Passa Quatro, SP). Braz. J. Ecol. 1998, 18. Available online: http://ecologia.ib.usp.br/lepac/conservacao/Artigos/83_zoneamento_eco.pdf (accessed on 1 July 2022).
- Veloso, H.P.; Rangel Filho, A.L.R.; Lima, J.C.A. Classificação da Vegetação Brasileira, Adaptada a um Sistema Universal; Ministério da Economia, Fazenda e Planejamento, Fundação Instituto Brasileiro de Geografia e Estatística, Diretoria de Geociências, Departamento de Recursos Naturais e Estudos Ambientais: Rio de Janeiro, Brazil, 1991; ISBN 978-85-240-0384-4. Available online: http://www.botanicaamazonica.wiki.br/labotam/lib/exe/fetch.php?media=projetos:campinas:biblio:veloso_etal_1991.pdf (accessed on 1 July 2022).
- Sudia, W.D.; Chamberlain, R.W. Battery-operated light trap, an improved model. Mosq. News 1962, 22, 126–129. Available online: https://www.biodiversitylibrary.org/content/part/JAMCA/JAMCA_V04_N4_P536-538.pdf (accessed on 1 July 2022).
- Reinert, J.E. Revised list of abbreviations for genera and subgenera of culicidae (diptera) and notes on generic and subgeneric changes. J. Am. Mosq. Control. Assoc. 2001, 17, 51–55. Available online: https://www.biodiversitylibrary.org/content/part/JAMCA/JAMCA_V17_N1_P051-055.pdf (accessed on 1 July 2022).
- Lane, J. Neotropical Culicidae; EDUSP: São Paulo, Brazil, 1953; Volume 2. [Google Scholar]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples 2013. Available online: http://purl.oclc.org/estimates (accessed on 1 July 2022).
- Hammer, O.; Harper, D.; Ryan, P. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. Available online: https://palaeo-electronica.org/2001_1/past/past.pdf (accessed on 1 July 2022).
- Guimarães, A.É.; Arlé, M. Mosquitos no parque nacional da serra dos órgãos, estado do rio de janeiro, Brasil: I- distribuição estacional. Mem. Inst. Oswaldo Cruz 1984, 79, 309–323. [Google Scholar] [CrossRef]
- Forattini, O.P.; Gomes, A.D.C. Biting activity of Aedes scapularis (Rondani) and Haemagogus mosquitoes in southern Brazil (Diptera: Culicidae). Rev. Saúde. Pública. 1988, 22, 84–93. [Google Scholar] [CrossRef]
- Gomes, A.D.C.; Torres, M.A.N.; de Paula, M.B.; Fernandes, A.; Marassá, A.M.; Consales, C.A.; Fonseca, D.F. Ecologia de Haemagogus e Sabethes (Diptera: Culicidae) em áreas epizoóticas do vírus da febre amarela, rio grande do sul, Brasil. Epidemiol. E Serviços. Saúde. 2010, 19, 101–113. [Google Scholar] [CrossRef]
- Silva, J.D.S.; Pacheco, J.B.; Alencar, J.; Guimarães, A.E. Biodiversity and influence of climatic factors on mosquitoes (Diptera: Culicidae) around the Peixe Angical hydroelectric scheme in the state of Tocantins, Brazil. Mem. Inst. Oswaldo Cruz 2010, 105, 155–162. [Google Scholar] [CrossRef]
- Alencar, J.; Pacheco, J.B.; Silva, J.D.S.; Silva, S.O.F.; Guimarães, A.E. Influence of climatic factors on Psorophora (Janthinosoma) albigenu in Pantanal Landscape, Mato Grosso State, Brazil. J. Am. Mosq. Control. Assoc. 2018, 34, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Couto-Lima, D.; Andreazzi, C.S.; Leite, P.J.; Bersot, M.I.L.; Alencar, J.; Lourenço-de-Oliveira, R. Seasonal population dynamics of the primary yellow fever vector Haemagogus leucocelaenus (Dyar & Shannon) (Diptera: Culicidae) is mainly influenced by temperature in the atlantic forest, Southeast Brazil. Mem. Inst. Oswaldo Cruz. 2020, 115. [Google Scholar] [CrossRef]
- Carvalho, M.E.S.D.; Naves, H.A.M.; Carneiro, E.; de Miranda, M.F. Distribuição vertical de mosquitos dos gêneros Haemagogus e Sabehtes, em zona urbana de Goiânia-Goiás-Brasil. Rev. Patol. Trop. J. Trop. Pathol. 1997, 26. [Google Scholar] [CrossRef]
- Lira-Vieira, A.R.; Gurgel-Gonçalves, R.; Moreira, I.M.; Yoshizawa, M.A.C.; Coutinho, M.L.; Prado, P.S.; de Souza, J.L.; Chaib, A.J.D.M.; Moreira, J.S.; de Castro, C.N. Ecological aspects of mosquitoes (Diptera: Culicidae) in the gallery forest of Brasilia National Park, Brazil, with an emphasis on potential vectors of yellow fever. Rev. Soc. Bras. Med. Trop. 2013, 46, 566–574. [Google Scholar] [CrossRef]
- Werneck, F.P.; Nogueira, C.; Colli, G.R.; Sites, J.W., Jr.; Costa, G.C. Climatic stability in the Brazilian Cerrado: Implications for biogeographical connections of south american savannas, species richness and conservation in a biodiversity hotspot. J. Biogeogr. 2012, 39, 1695–1706. [Google Scholar] [CrossRef]
- Camargo-Neves, V.L.F.; Poletto, D.W.; Rodas, L.A.C.; Pacchioli, M.; Cardoso, R.P.; Scandar, S.A.S.; Sampaio, S.M.P.; Koyanagui, P.H.; Botti, M.V.; Mucci, L.F.; et al. Entomological investigation of a sylvatic yellow fever area in São Paulo State, Brazil. Cad Saude Públ. 2005, 21, 1278–1286. [Google Scholar] [CrossRef]
- Moreno, E.S.; Rocco, I.M.; Bergo, E.; Brasil, R.A.; Siciliano, M.M.; Suzuki, A.; Silveira, V.R.; Bisordi, I.; de Souza, R.P. Reemergence of yellow fever: Detection of transmission in the State of São Paulo, Brazil, 2008. Rev. Soc. Bras. Med. Trop. 2011, 44, 290–296. [Google Scholar] [CrossRef]
- Cunha, M.S.; da Costa, A.C.; de Azevedo Fernandes, N.C.C.; Guerra, J.M.; dos Santos, F.C.P.; Nogueira, J.S.; D’Agostino, L.G.; Komninakis, S.V.; Witkin, S.S.; Ressio, R.A.; et al. Epizootics Due to Yellow Fever Virus in São Paulo State, Brazil: Viral dissemination to new areas (2016–2017). Sci. Rep. 2019, 9, 5474. [Google Scholar] [CrossRef]
- Cunha, M.S.; Tubaki, R.M.; de Menezes, R.M.T.; Pereira, M.; Caleiro, G.S.; Coelho, E.; del Castillo Saad, L.; de Azevedo Fernandes, N.C.C.; Guerra, J.M.; Nogueira, J.S. Possible Non-sylvatic transmission of yellow fever between non-human primates in São Paulo City, Brazil, 2017–2018. Sci. Rep. 2020, 10, 15751. [Google Scholar] [CrossRef]
- Fidelis, A.T.; de Godoy, S.A.P. Estrutura de um cerrado strico sensu na Gleba Cerrado Pé-de-Gigante, Santa Rita do Passa Quatro, SP. Acta Bot. Bras. 2003, 17, 531–539. [Google Scholar] [CrossRef]
- Marcondes, C.B.; Alencar, J. Revisão de mosquitos Haemagogus Williston (Diptera: Culicidae) do Brasil. Rev. Biomed. 2010, 21, 18. Available online: http://www.revbiomed.uady.mx/pdf/rb102139.pdf (accessed on 1 July 2022).
- Alencar, J.; Dégallier, N.; Hannart, A.; Silva, J.D.S.; Pacheco, J.B.; Guimarães, A.E. Circadian and Seasonal Preferences for Hematophagy among Haemagogus capricornii, Hg. janthinomys, and Hg. leucocelaenus (Diptera: Culicidae) in Different Regions of Brazil. J. Vector Ecol. 2008, 33, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Hendy, A.; Valério, D.; Fé, N.F.; Hernandez-Acosta, E.; Mendonça, C.; Andrade, E.; Pedrosa, I.; Costa, E.R.; Júnior, J.T.A.; Assunção, F.P.; et al. Microclimate and the vertical stratification of potential bridge vectors of mosquito-borne viruses captured by nets and ovitraps in a central amazonian forest bordering Manaus, Brazil. Sci. Rep. 2021, 11, 21129. [Google Scholar] [CrossRef] [PubMed]
- Câmara, D.C.P.; Codeço, C.T.; Ayllón, T.; Nobre, A.A.; Azevedo, R.C.; Ferreira, D.F.; da Silva Pinel, C.; Rocha, G.P.; Honório, N.A. Entomological surveillance of Aedes mosquitoes: Comparison of different collection methods in an endemic area in RIO de Janeiro, Brazil. Trop. Med. Infect. Dis. 2022, 7, 114. [Google Scholar] [CrossRef]
- Cano, M.E.; Marti, G.A.; Alencar, J.; Silva, S.O.F.; Micieli, M.V. Categorization by score of mosquito species (Diptera: Culicidae) related to yellow fever epizootics in Argentina. J. Med. Entomol. 2022, 59, 1766–1777. [Google Scholar] [CrossRef]
- Monath, T.P.; Vasconcelos, P.F.C. Yellow fever. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2015, 64, 160–173. [Google Scholar] [CrossRef]
- de Abreu, F.V.S.; Ribeiro, I.P.; Ferreira-de-Brito, A.; dos Santos, A.A.C.; de Miranda, R.M.; Bonelly, I.D.S.; Neves, M.S.A.S.; Bersot, M.I.; dos Santos, T.P.; Gomes, M.Q.; et al. Haemagogus leucocelaenus and Haemagogus janthinomys are the primary vectors in the major yellow fever outbreak in Brazil, 2016–2018. Emerg. Microbes Infect. 2019, 8, 218–231. [Google Scholar] [CrossRef]
- Pinto, C.S.; Confalonieri, U.E.; Mascarenhas, B.M. Ecology of Haemagogus sp. and Sabethes sp.(Diptera: Culicidae) in relation to the microclimates of the Caxiuanã national forest, Pará, Brazil. Mem. Inst. Oswaldo Cruz 2009, 104, 592–598. [Google Scholar] [CrossRef]
- Confalonieri, U.E.C.; Neto, C.C. Diversity of mosquito vectors (Diptera: Culicidae) in Caxiuanã, Pará, Brazil. Interdiscip. Perspect. Infect. Dis. 2012, 2012, 741273. [Google Scholar] [CrossRef][Green Version]
- Cardoso, J.D.C.; de Almeida, M.A.; dos Santos, E.; da Fonseca, D.F.; Sallum, M.A.; Noll, C.A.; Monteiro, H.A.D.O.; Cruz, A.C.; Carvalho, V.L.; Pinto, E.V.; et al. Yellow fever virus in Haemagogus leucocelaenus and Aedes serratus Mosquitoes, southern Brazil, 2008. Emerg. Infect. Dis. 2010, 16, 1918. [Google Scholar] [CrossRef]
- De Souza, R.P.; Petrella, S.; Coimbra, T.L.M.; Maeda, A.Y.; Rocco, I.M.; Bisordi, I.; Silveira, V.R.; Pereira, L.E.; Suzuki, A.; Silva, S.J.D.S.; et al. Isolation of yellow fever virus (YFV) from naturally infectied Haemagogus (Conopostegus) leucocelaenus (diptera, cukicudae) in São Paulo State, Brazil, 2009. Rev. Inst. Med. Trop. São Paulo 2011, 53, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Obara, M.T.; Monteiro, H.; de Paula, M.B.; Gomes, A.D.C.; Yoshizawa, M.A.C.; Lira, A.R.; Boffil, M.I.R.; de Carvalho, M.D.S.L. Infecção Natural de Haemagogus janthinomys e Haemagogus leucocelaenus pelo vírus da febre amarela no distrito federal, Brasil, 2007-2008. Epidemiol. E Serviços Saúde 2012, 21, 457–463. [Google Scholar] [CrossRef]
- Brasil, M.D.S. Guia de Vigilância do Culex Quinquefasciatus; Normas e Manuais Técnicos, 3rd ed.; Ministério da Saúde: Brasília, Brazil, 2011. Available online: https://bvsms.saude.gov.br/bvs/publicacoes/guia_vigilancia_culex_quinquefasciatus.pdf (accessed on 1 July 2022).
- Bowman, L.R.; Runge-Ranzinger, S.; McCall, P.J. Assessing the relationship between vector indices and dengue transmission: A systematic review of the evidence. PLoS Negl. Trop. Dis. 2014, 8, e2848. [Google Scholar] [CrossRef] [PubMed]
- Tabbabi, A.; Daaboub, J. First investigation of deltamethrin pyrethroid susceptibility and resistance status of Anopheles labranchiae (Falleroni, 1926), potential malaria vector in Tunisia. Asian Pac. J. Trop. Biomed. 2017, 7, 1067–1070. [Google Scholar] [CrossRef]




{kind=link}
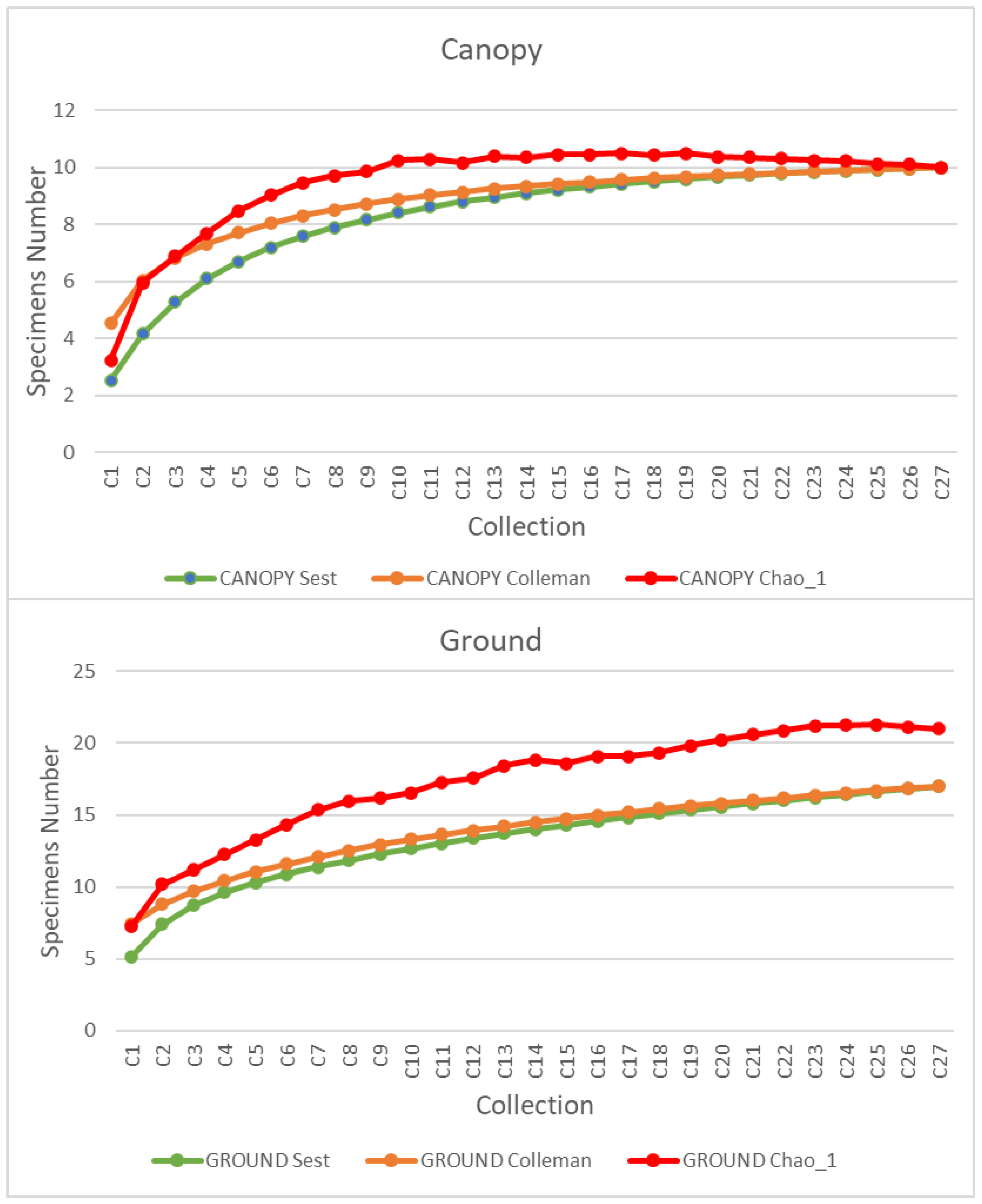{kind=link}
{kind=link}
| Taxa | Canopy | Ground | Total | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BGL | CO2 | CO2 + BGL1 | CO2 + BGL2 | NET | TOTAL | % | BGL | CO2 | CO2 + BGL1 | CO2 + BGL2 | NET | TOTAL | % | N | % | |
| Aedes albopictus | 4 | 4 | 1.7 | 2 | 9 | 6 | 17 | 115 | 149 | 11.4 | 153 | 10.0 | ||||
| Aedes scapularis | 3 | 12 | 1 | 18 | 34 | 14.8 | 14 | 65 | 78 | 34 | 408 | 599 | 46.0 | 633 | 41.3 | |
| Aedes serratus | 2 | 2 | 0.2 | 2 | 0.1 | |||||||||||
| Coquillettidia juxtamansonia | 1 | 1 | 0.1 | 1 | 0.1 | |||||||||||
| Culex sp. | 1 | 7 | 8 | 3.5 | 1 | 1 | 4 | 22 | 28 | 2.2 | 36 | 2.4 | ||||
| Haemagogus janthinomys/capricornii | 1 | 2 | 1 | 69 | 73 | 31.9 | 1 | 6 | 5 | 134 | 146 | 11.2 | 219 | 14.3 | ||
| Haemagogus leucocelaenus | 2 | 2 | 0.2 | 2 | 0.1 | |||||||||||
| Limatus durhamii | 1 | 1 | 0.4 | 1 | 0.1 | |||||||||||
| Psorophora albigenu | 1 | 11 | 1 | 16 | 29 | 12.7 | 8 | 10 | 18 | 9 | 123 | 168 | 12.9 | 197 | 12.9 | |
| Psorophora discrucians | 1 | 1 | 0.1 | 1 | 0.1 | |||||||||||
| Psorophora ferox | 1 | 3 | 4 | 0.3 | 4 | 0.3 | ||||||||||
| Psorophora varipes | 4 | 3 | 7 | 0.5 | 7 | 0.5 | ||||||||||
| Psorophora sp. | 1 | 3 | 4 | 0.3 | 4 | 0.3 | ||||||||||
| Sabethes albiprivus | 3 | 5 | 35 | 43 | 18.8 | 1 | 26 | 7 | 18 | 92 | 144 | 11.1 | 187 | 12.2 | ||
| Sabethes belisarioi | 1 | 5 | 6 | 2.6 | 1 | 3 | 4 | 0.3 | 10 | 0.7 | ||||||
| Sabethes glaucodaemon | 1 | 2 | 3 | 22 | 28 | 12.2 | 2 | 3 | 5 | 22 | 32 | 2.5 | 60 | 3.9 | ||
| Sabethes purpureus | 3 | 3 | 1.3 | 2 | 1 | 4 | 7 | 0.5 | 10 | 0.7 | ||||||
| Sabethes tarsopus | 1 | 1 | 0.1 | 1 | 0.1 | |||||||||||
| Sabethes sp. | 1 | 1 | 2 | 0.2 | 2 | 0.1 | ||||||||||
| Wyeomyia confusa | 1 | 1 | 0.1 | 1 | 0.1 | |||||||||||
| Total | 1 | 8 | 27 | 13 | 180 | 229 | 100.0 | 27 | 119 | 128 | 89 | 939 | 1302 | 100.0 | 1531 | 100.0 |
| % | 0.4 | 3.5 | 11.8 | 5.7 | 78.6 | 100.0 | 2.1 | 9.1 | 9.8 | 6.8 | 72.1 | 100.0 | 100.0 | |||
| 15.0 | 85.0 | |||||||||||||||
| Parameter | Canopy | Ground | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| BGL | CO2 | CO2 + BGL1 | CO2 + BGL2 | NET | BGL | CO2 | CO2 + BGL1 | CO2 + BGL2 | NET | |
| S (Taxa) | 1 | 4 | 4 | 7 | 10 | 5 | 10 | 9 | 7 | 16 |
| N (Abundance) | 1 | 8 | 27 | 13 | 180 | 26 | 118 | 127 | 89 | 936 |
| D (Dominance) | 1 | 0.31 | 0.37 | 0.23 | 0.22 | 0.39 | 0.37 | 0.41 | 0.24 | 0.25 |
| 1-D (Geni-Simpson Index) | 0 | 0.69 | 0.63 | 0.77 | 0.78 | 0.61 | 0.63 | 0.59 | 0.76 | 0.75 |
| H (Shannon Index) | 0 | 1.26 | 1.11 | 1.69 | 1.80 | 1.14 | 1.37 | 1.37 | 1.61 | 1.70 |
| Strata | Canopy | Ground | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Methods | BGL | CO2 | CO2 + BGL1 | CO2 + BGL2 | NET | BGL | CO2 | CO2 + BGL1 | CO2 + BGL2 | NET | |
| p valor | |||||||||||
| Canopy | BGL | 0.622 | 0.096 | 0.101 | 0.184 | 0.571 | 0.520 | 0.853 | 0.492 | 0.536 | |
| CO2 | −0.12 | 0.001 | 0.011 | 0.001 | 0.004 | 0.004 | 0.001 | 0.001 | 0.001 | ||
| CO2 + BG1 | 0.39 | 0.68 | 0.024 | 0.002 | 0.064 | 0.022 | 0.008 | 0.008 | 0.002 | ||
| CO2 + BG2 | 0.39 | 0.57 | 0.52 | 0.000 | 0.096 | 0.006 | 0.000 | 0.004 | 0.040 | ||
| NET | 0.32 | 0.69 | 0.66 | 0.73 | 0.011 | 0.005 | 0.003 | 0.002 | 0.000 | ||
| Ground | BGL | −0.14 | 0.63 | 0.43 | 0.39 | 0.57 | 0.001 | 0.001 | 0.002 | 0.001 | |
| CO2 | 0.16 | 0.62 | 0.52 | 0.60 | 0.62 | 0.71 | 0.000 | 0.000 | 0.004 | ||
| CO2 + BG1 | 0.05 | 0.69 | 0.59 | 0.75 | 0.65 | 0.71 | 0.74 | 0.000 | 0.002 | ||
| CO2 + BG2 | 0.17 | 0.70 | 0.59 | 0.63 | 0.67 | 0.66 | 0.91 | 0.81 | 0.002 | ||
| NET | 0.15 | 0.68 | 0.66 | 0.47 | 0.81 | 0.69 | 0.63 | 0.67 | 0.65 | ||
| ρ valor | |||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mucci, L.F.; Bergo, E.S.; Deus, J.T.d.; Reginato, S.L.; Pereira, M.; Camargo-Neves, V.L.F.d. Evaluation of Methods for Collecting Mosquitoes (Culicidae: Diptera) in Canopy and Ground Strata in the Brazilian Savanna. Trop. Med. Infect. Dis. 2022, 7, 446. https://doi.org/10.3390/tropicalmed7120446
Mucci LF, Bergo ES, Deus JTd, Reginato SL, Pereira M, Camargo-Neves VLFd. Evaluation of Methods for Collecting Mosquitoes (Culicidae: Diptera) in Canopy and Ground Strata in the Brazilian Savanna. Tropical Medicine and Infectious Disease. 2022; 7(12):446. https://doi.org/10.3390/tropicalmed7120446
Chicago/Turabian StyleMucci, Luis Filipe, Eduardo Sterllino Bergo, Juliana Telles de Deus, Simone Luchetta Reginato, Mariza Pereira, and Vera Lucia Fonseca de Camargo-Neves. 2022. "Evaluation of Methods for Collecting Mosquitoes (Culicidae: Diptera) in Canopy and Ground Strata in the Brazilian Savanna" Tropical Medicine and Infectious Disease 7, no. 12: 446. https://doi.org/10.3390/tropicalmed7120446
APA StyleMucci, L. F., Bergo, E. S., Deus, J. T. d., Reginato, S. L., Pereira, M., & Camargo-Neves, V. L. F. d. (2022). Evaluation of Methods for Collecting Mosquitoes (Culicidae: Diptera) in Canopy and Ground Strata in the Brazilian Savanna. Tropical Medicine and Infectious Disease, 7(12), 446. https://doi.org/10.3390/tropicalmed7120446