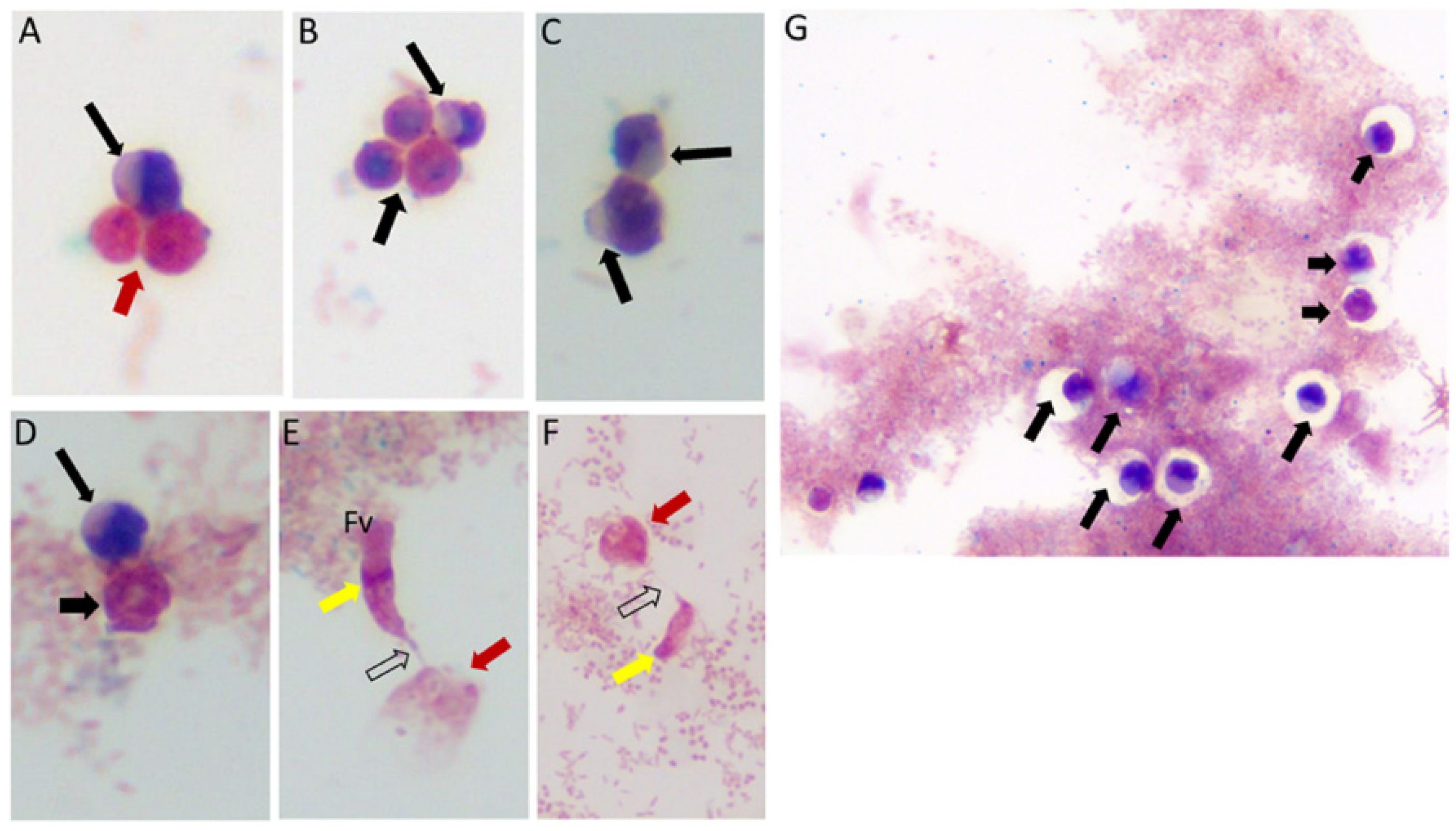
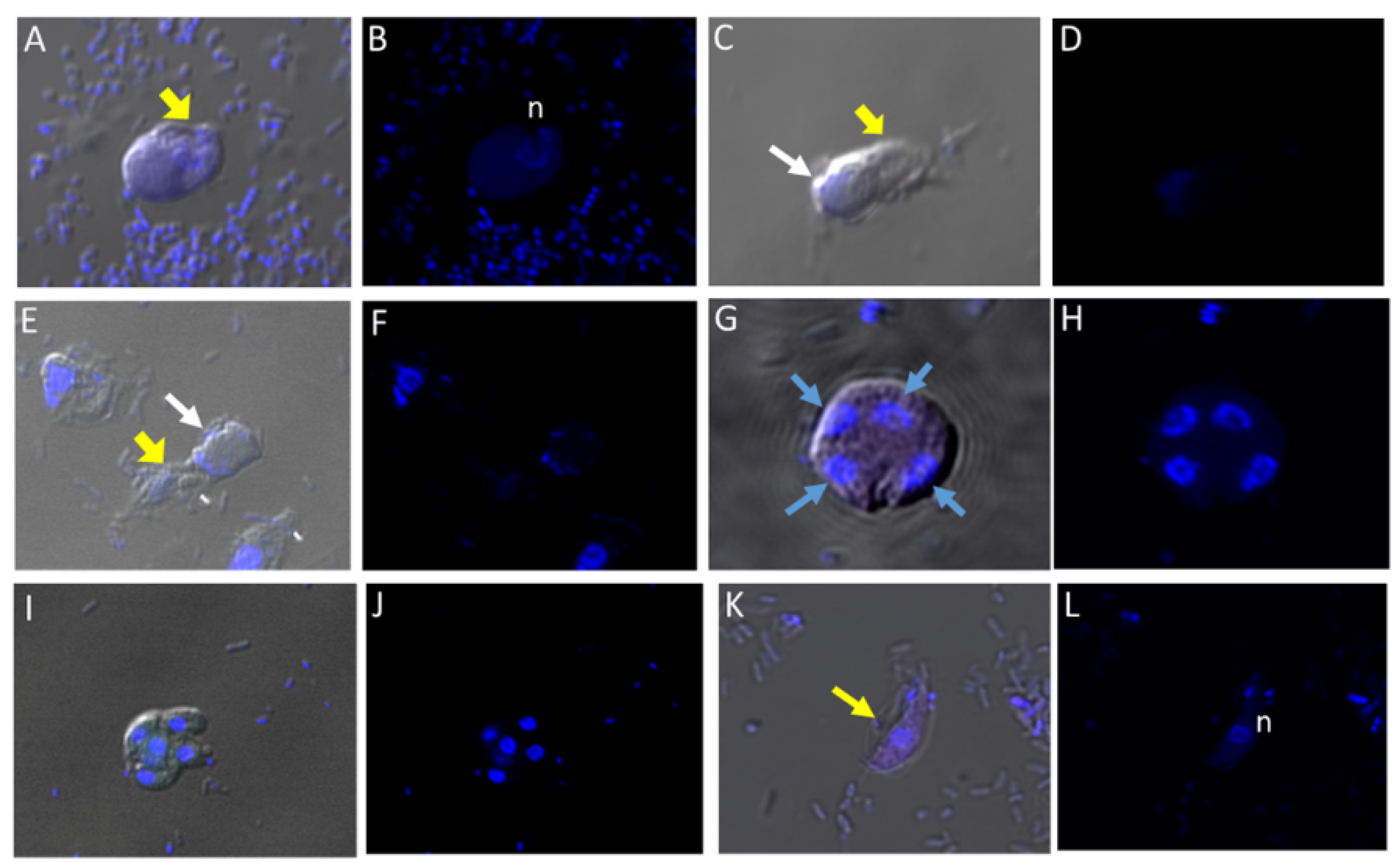
Figure 1.
Life cycle stages observed from di-protist cultures of Colpodella sp. (ATCC 50594) and Parabodo caudatus. Black arrows in panels (A–D,G) depict young demilune cysts of Colpodella sp. (ATCC 50594). Black arrowheads show mature cysts, red arrows show cysts of P. caudatus. In panels (E,F) yellow arrows show Colpodella sp. (ATCC 50594) trophozoites in myzo-cytosis with P. caudatus prey (red arrows). Open arrows identify tubular tether formed between predator and prey. Clear zones separating bacteria from cysts of Colpodella sp. (ATCC 50594) were observed, black arrows.
Figure 1.
Life cycle stages observed from di-protist cultures of Colpodella sp. (ATCC 50594) and Parabodo caudatus. Black arrows in panels (A–D,G) depict young demilune cysts of Colpodella sp. (ATCC 50594). Black arrowheads show mature cysts, red arrows show cysts of P. caudatus. In panels (E,F) yellow arrows show Colpodella sp. (ATCC 50594) trophozoites in myzo-cytosis with P. caudatus prey (red arrows). Open arrows identify tubular tether formed between predator and prey. Clear zones separating bacteria from cysts of Colpodella sp. (ATCC 50594) were observed, black arrows.
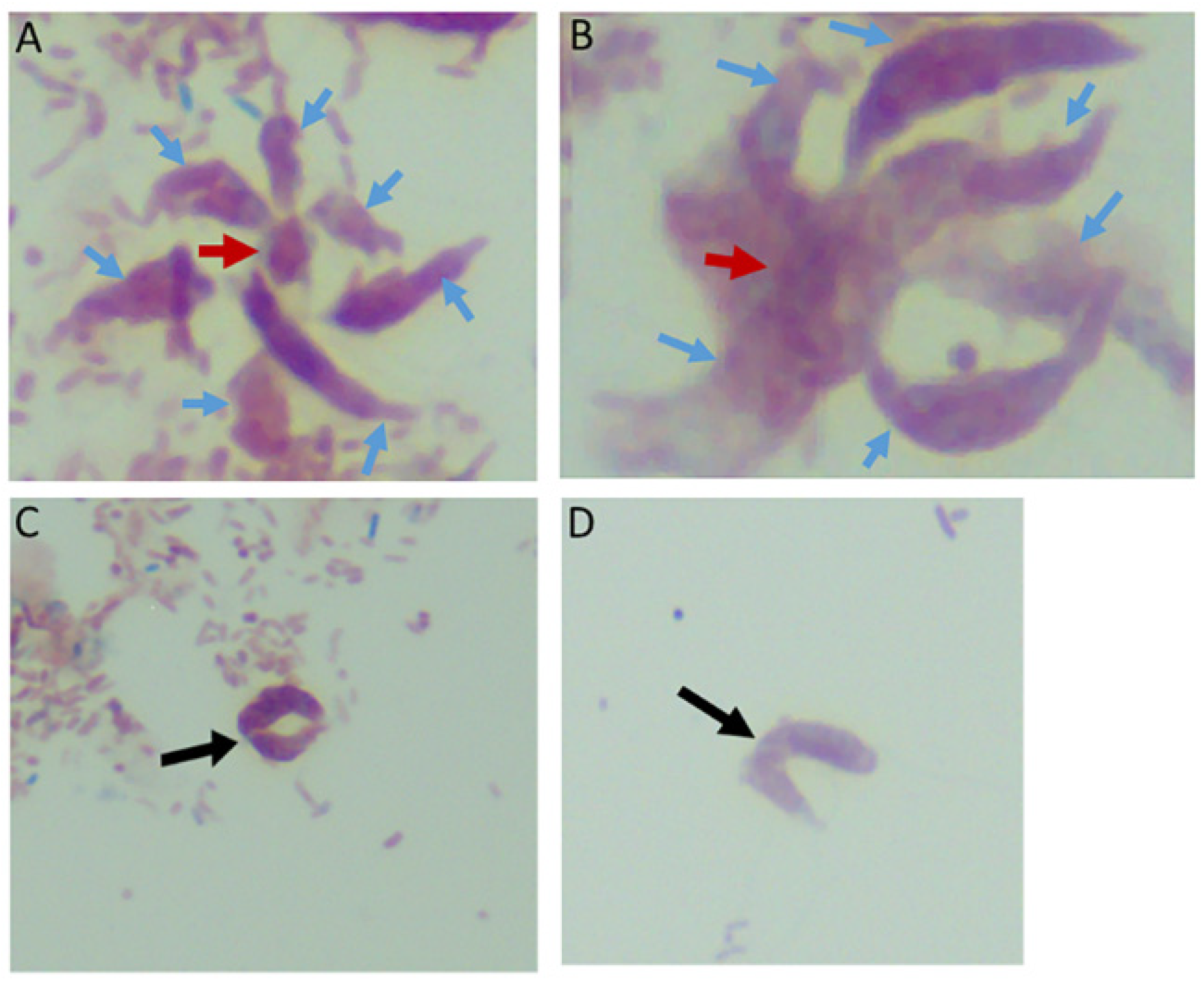
Figure 2.
Multiple Colpodella sp. (ATCC 50594) trophozoites (blue arrows) feeding on a single P. caudatus prey (red arrowhead) in panels (A,B). Paired juveniles still attached after egress from the cyst were observed with point of attachment shown by the black arrow in panels (C,D).
Figure 2.
Multiple Colpodella sp. (ATCC 50594) trophozoites (blue arrows) feeding on a single P. caudatus prey (red arrowhead) in panels (A,B). Paired juveniles still attached after egress from the cyst were observed with point of attachment shown by the black arrow in panels (C,D).
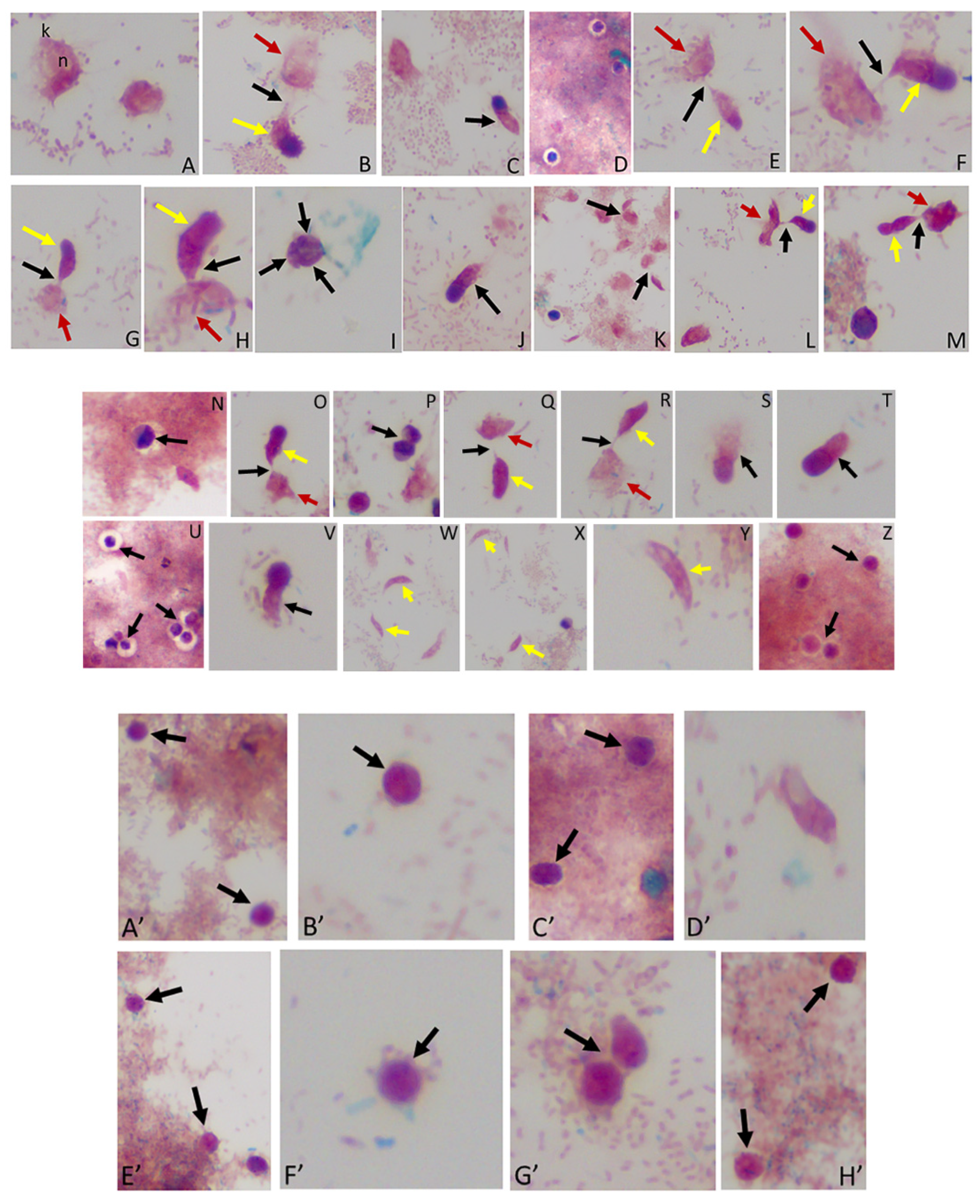
Figure 3.
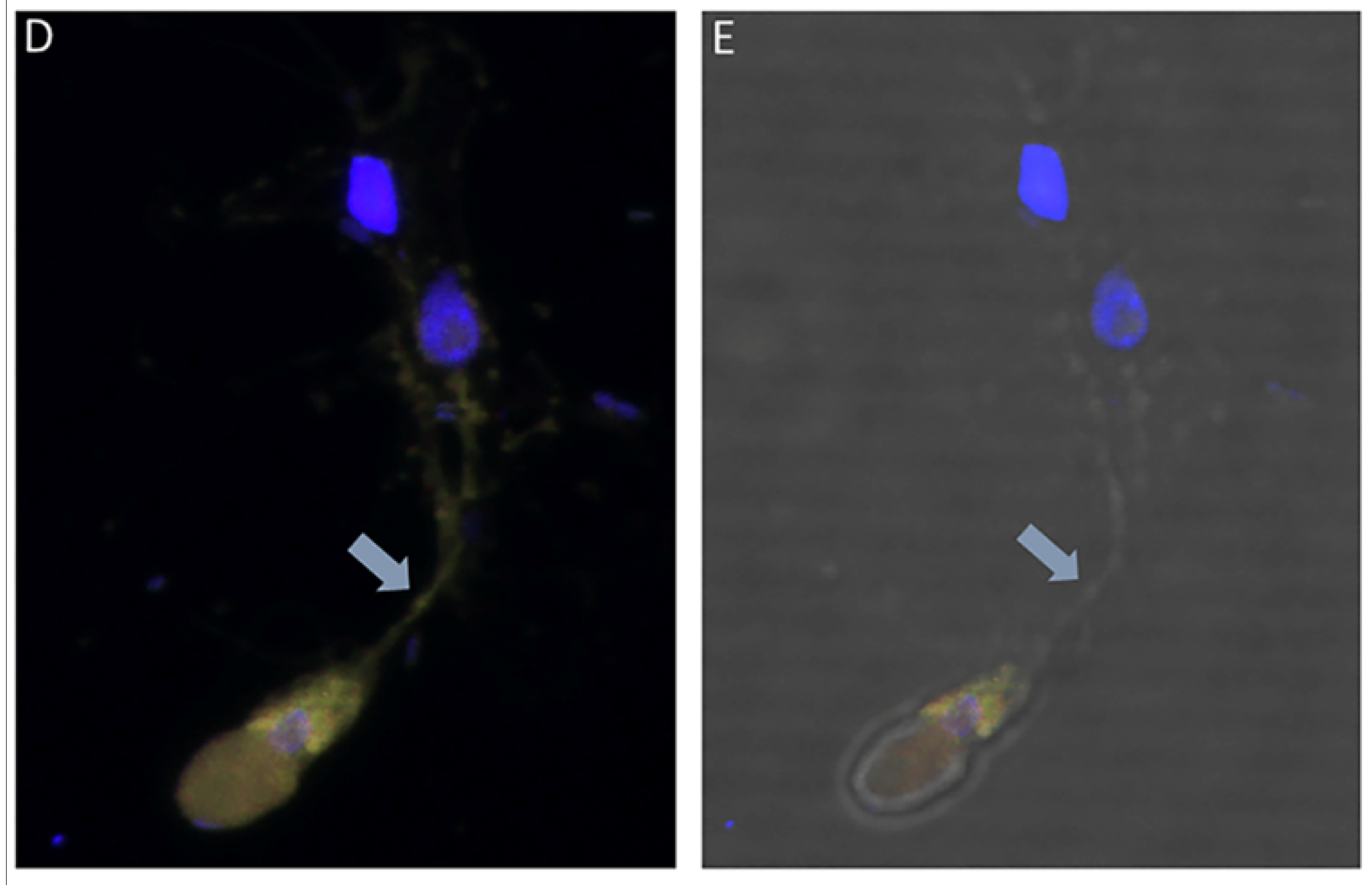
(A–M): Time course experiments showing development and timing of life cycle stages (A,B 20 h), (C,D 22h), (E,F 24 h), (G–I 25 h), (J,K 26 h), and (L,M 27 h). Kinetoplast (k) and nucleus (n) of Parabodo caudatus is shown in panel (A). Black arrows in panels (B,E–G,H,K–M) identify tubular tethers between Colpodella sp. (ATCC 50594) (yellow arrows) and P. caudatus (red arrows). Black arrowheads identify pre-cysts. A mature cyst of Colpodella sp. (ATCC 50594) is shown in panel I. Young demilune cysts of Colpodella sp. (ATCC 50594) shown in panel (D). (N–Z): Time course experiments showing development and timing of life cycle stage transitions in culture. (N,O 28 h), (P,Q 29 h), (R–T 30 h), (U,V 32 h), (W,X 34 h) and (Y,Z 36 h). Young demilune cysts are shown in panels (N,P,U). Yellow arrows identify Colpodella sp. (ATCC 50594) trophozoites in myzocytosis with P. caudatus prey (red arrows) in panels (O,Q,R). Black arrows identify tubular tethers. Black arrowheads in panels (S,T,V) identify pre-cysts of Colpodella sp. (ATCC 50594). Young trophozoites of Colpodella sp. (ATCC 50594) were identified in panels (W–Y) and mature cysts of Colpodella sp. (ATCC 50594) were seen in panel (Z). Zones of inhibition/clear zones were observed surrounding and separating cysts of Colpodella sp. (ATCC 50594) from bacteria (antibacterial zone). (A’–H’): Time course experiments showing development and timing of life cycle stage transitions in culture. (A’,B’ 38 h), (C’,D’ 40 h), (E’,F’ 5 days) and (G’,H’ 7 days). Mature cysts of Colpodella sp. (ATCC 50594) are shown in panels (A’–E’). Resting cysts of Colpodella sp. (ATCC 50594) are shown in panels (F’−H’).
Figure 3.
(A–M): Time course experiments showing development and timing of life cycle stages (A,B 20 h), (C,D 22h), (E,F 24 h), (G–I 25 h), (J,K 26 h), and (L,M 27 h). Kinetoplast (k) and nucleus (n) of Parabodo caudatus is shown in panel (A). Black arrows in panels (B,E–G,H,K–M) identify tubular tethers between Colpodella sp. (ATCC 50594) (yellow arrows) and P. caudatus (red arrows). Black arrowheads identify pre-cysts. A mature cyst of Colpodella sp. (ATCC 50594) is shown in panel I. Young demilune cysts of Colpodella sp. (ATCC 50594) shown in panel (D). (N–Z): Time course experiments showing development and timing of life cycle stage transitions in culture. (N,O 28 h), (P,Q 29 h), (R–T 30 h), (U,V 32 h), (W,X 34 h) and (Y,Z 36 h). Young demilune cysts are shown in panels (N,P,U). Yellow arrows identify Colpodella sp. (ATCC 50594) trophozoites in myzocytosis with P. caudatus prey (red arrows) in panels (O,Q,R). Black arrows identify tubular tethers. Black arrowheads in panels (S,T,V) identify pre-cysts of Colpodella sp. (ATCC 50594). Young trophozoites of Colpodella sp. (ATCC 50594) were identified in panels (W–Y) and mature cysts of Colpodella sp. (ATCC 50594) were seen in panel (Z). Zones of inhibition/clear zones were observed surrounding and separating cysts of Colpodella sp. (ATCC 50594) from bacteria (antibacterial zone). (A’–H’): Time course experiments showing development and timing of life cycle stage transitions in culture. (A’,B’ 38 h), (C’,D’ 40 h), (E’,F’ 5 days) and (G’,H’ 7 days). Mature cysts of Colpodella sp. (ATCC 50594) are shown in panels (A’–E’). Resting cysts of Colpodella sp. (ATCC 50594) are shown in panels (F’−H’).
![Tropicalmed 06 00127 g003]()
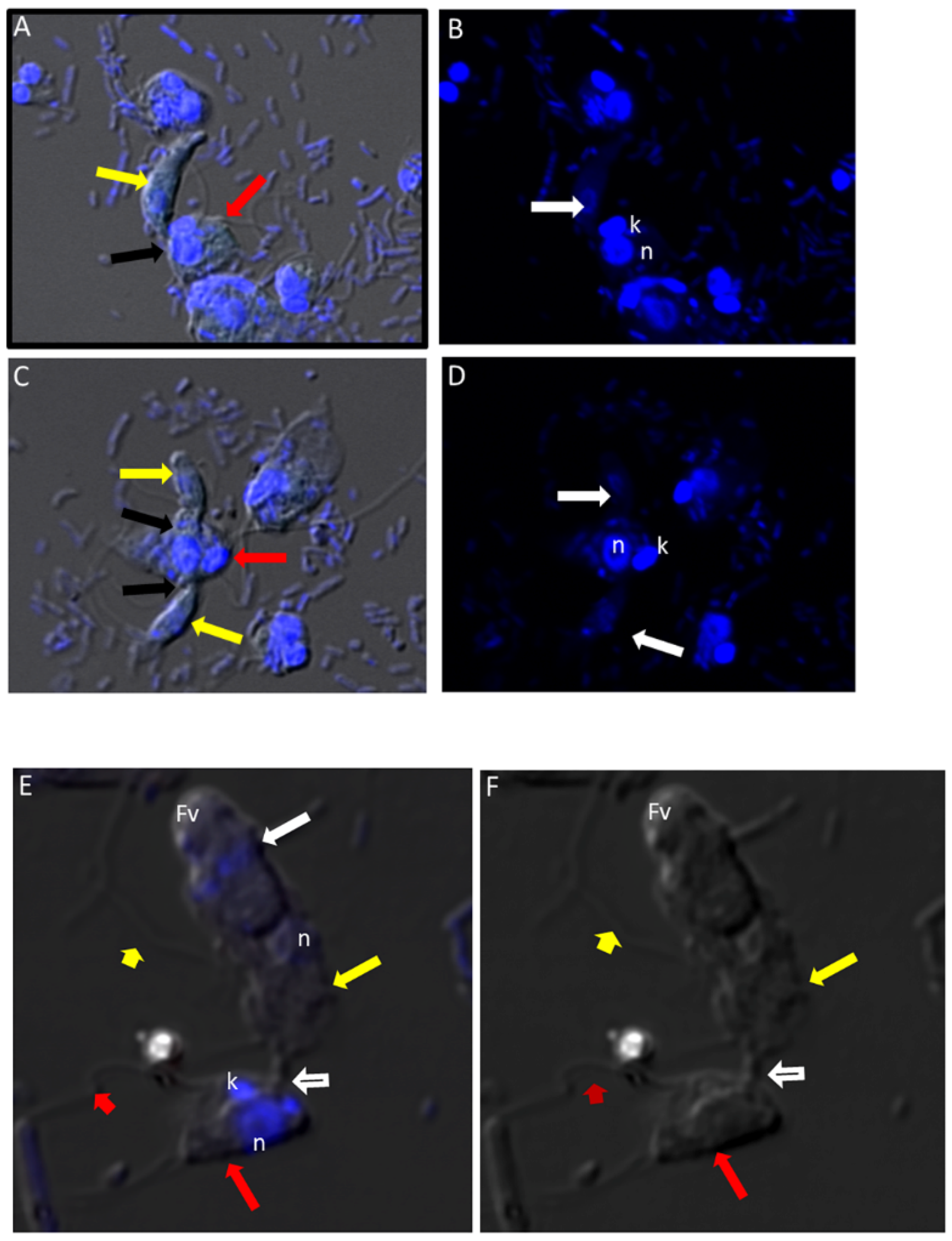
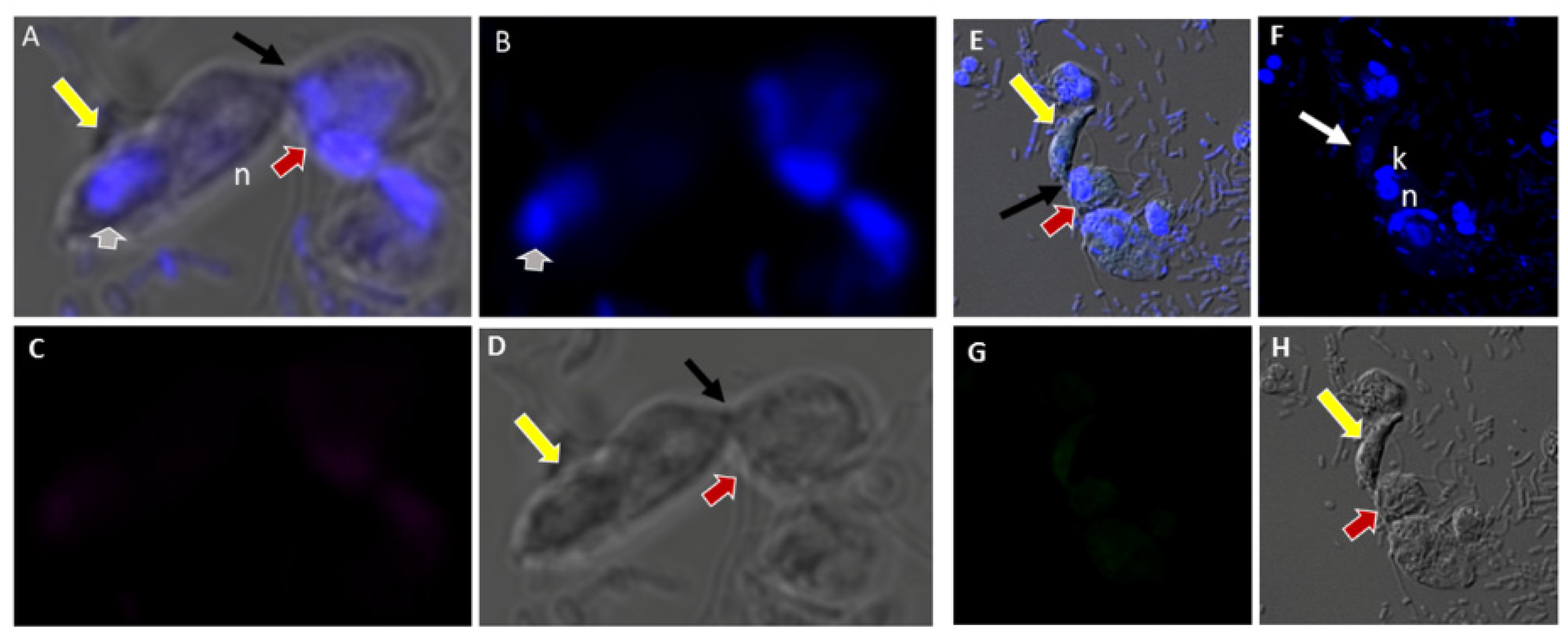
Figure 4.
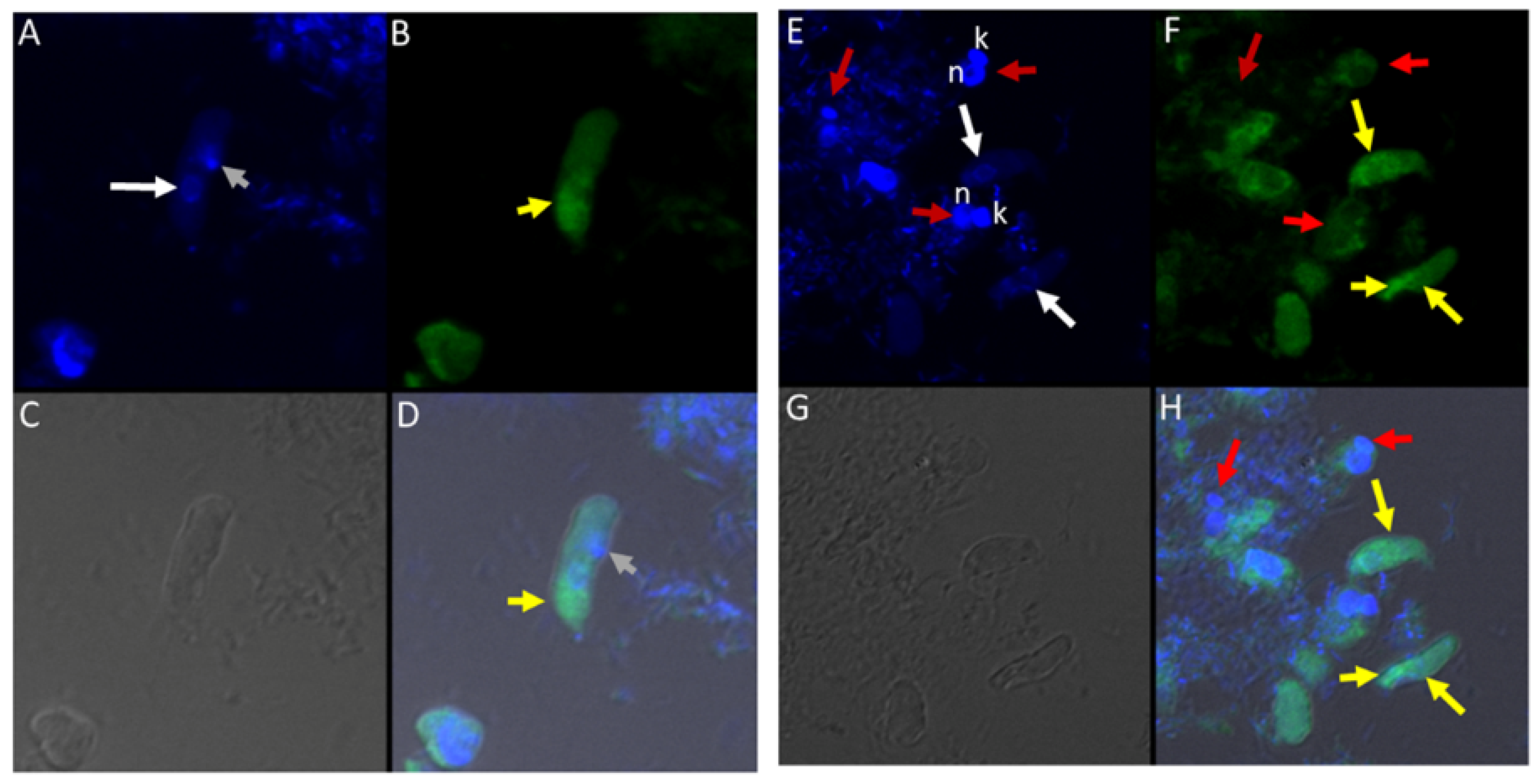
(A–D): Differential Interference contrast (DIC) microscopy of DAPI stained Colpodella sp. (ATCC 50594) life cycle stage transitions. (A,C) Merger of DIC and DAPI stained Colpodella sp. (ATCC 50594) trophozoites (yellow arrow) and P. caudatus prey (red arrow) in myzocytosis. Two predators feeding on one prey are shown in panel (C). The tubular tether joining predator and prey is indicated by the black arrow. (B,D) DAPI stained nuclei and kinetoplast. The kinetoplast (k) and nucleus (n) of P. caudatus is identified in panels (B,D). The central nucleus of Colpodella sp. (ATCC 50594) trophozoites is indicated by the white arrows. (E,F): Differential Interference contrast (DIC) microscopy of DAPI stained Colpodella sp. (ATCC 50594) life cycle stage transitions. (E) Merger of DIC and DAPI. (F) DIC alone. Colpodella sp. (ATCC 50594) trophozoite (yellow arrow) shown in myzocytosis with P. caudatis (red arrow). The tubular tether between predator and prey is indicated by the open arrow. DAPI stained aspirated contents can be seen at the posterior food vacuole of the Colpodella sp. (ATCC 50594). Yellow arrowhead identifies the flagella of Colpodella sp. (ATCC 50594) and the flagella of the prey are indicated by the red arrowhead. The kinetoplast (k) and nucleus of the prey and the nucleus of Colpodella sp. (ATCC 50594) were identified.
Figure 4.
(A–D): Differential Interference contrast (DIC) microscopy of DAPI stained Colpodella sp. (ATCC 50594) life cycle stage transitions. (A,C) Merger of DIC and DAPI stained Colpodella sp. (ATCC 50594) trophozoites (yellow arrow) and P. caudatus prey (red arrow) in myzocytosis. Two predators feeding on one prey are shown in panel (C). The tubular tether joining predator and prey is indicated by the black arrow. (B,D) DAPI stained nuclei and kinetoplast. The kinetoplast (k) and nucleus (n) of P. caudatus is identified in panels (B,D). The central nucleus of Colpodella sp. (ATCC 50594) trophozoites is indicated by the white arrows. (E,F): Differential Interference contrast (DIC) microscopy of DAPI stained Colpodella sp. (ATCC 50594) life cycle stage transitions. (E) Merger of DIC and DAPI. (F) DIC alone. Colpodella sp. (ATCC 50594) trophozoite (yellow arrow) shown in myzocytosis with P. caudatis (red arrow). The tubular tether between predator and prey is indicated by the open arrow. DAPI stained aspirated contents can be seen at the posterior food vacuole of the Colpodella sp. (ATCC 50594). Yellow arrowhead identifies the flagella of Colpodella sp. (ATCC 50594) and the flagella of the prey are indicated by the red arrowhead. The kinetoplast (k) and nucleus of the prey and the nucleus of Colpodella sp. (ATCC 50594) were identified.
![Tropicalmed 06 00127 g004]()
Figure 5.
(A–L): Differential Interference contrast (DIC) microscopy of DAPI stained Colpodella sp. (ATCC 50594) life cycle stage transitions. DIC and DAPI staining of pre-cyst and cyst stages of Colpodella sp. (ATCC 50594). Yellow arrowhead shows frayed anterior end of encysting trophozoites in panels (A,C,E). White arrows identify the DAPI stained aspirated contents from the prey in the posterior food vacuole. A four-nuclei cyst is identified in panels (G,H), a five-nuclei cyst in panels (I,J) and a young trophozoite in panels (K,L). The central nucleus (n) was identified.
Figure 5.
(A–L): Differential Interference contrast (DIC) microscopy of DAPI stained Colpodella sp. (ATCC 50594) life cycle stage transitions. DIC and DAPI staining of pre-cyst and cyst stages of Colpodella sp. (ATCC 50594). Yellow arrowhead shows frayed anterior end of encysting trophozoites in panels (A,C,E). White arrows identify the DAPI stained aspirated contents from the prey in the posterior food vacuole. A four-nuclei cyst is identified in panels (G,H), a five-nuclei cyst in panels (I,J) and a young trophozoite in panels (K,L). The central nucleus (n) was identified.
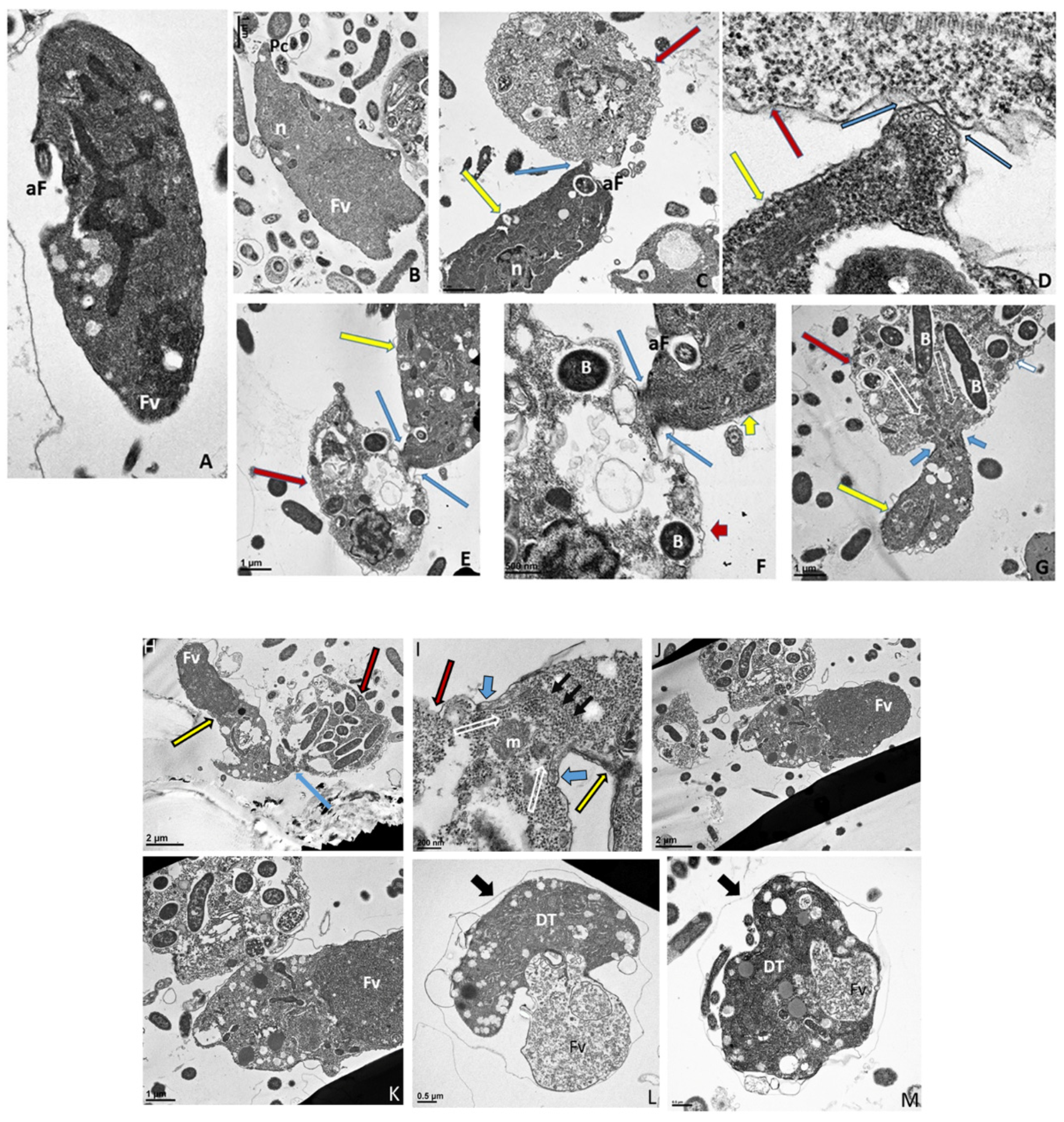
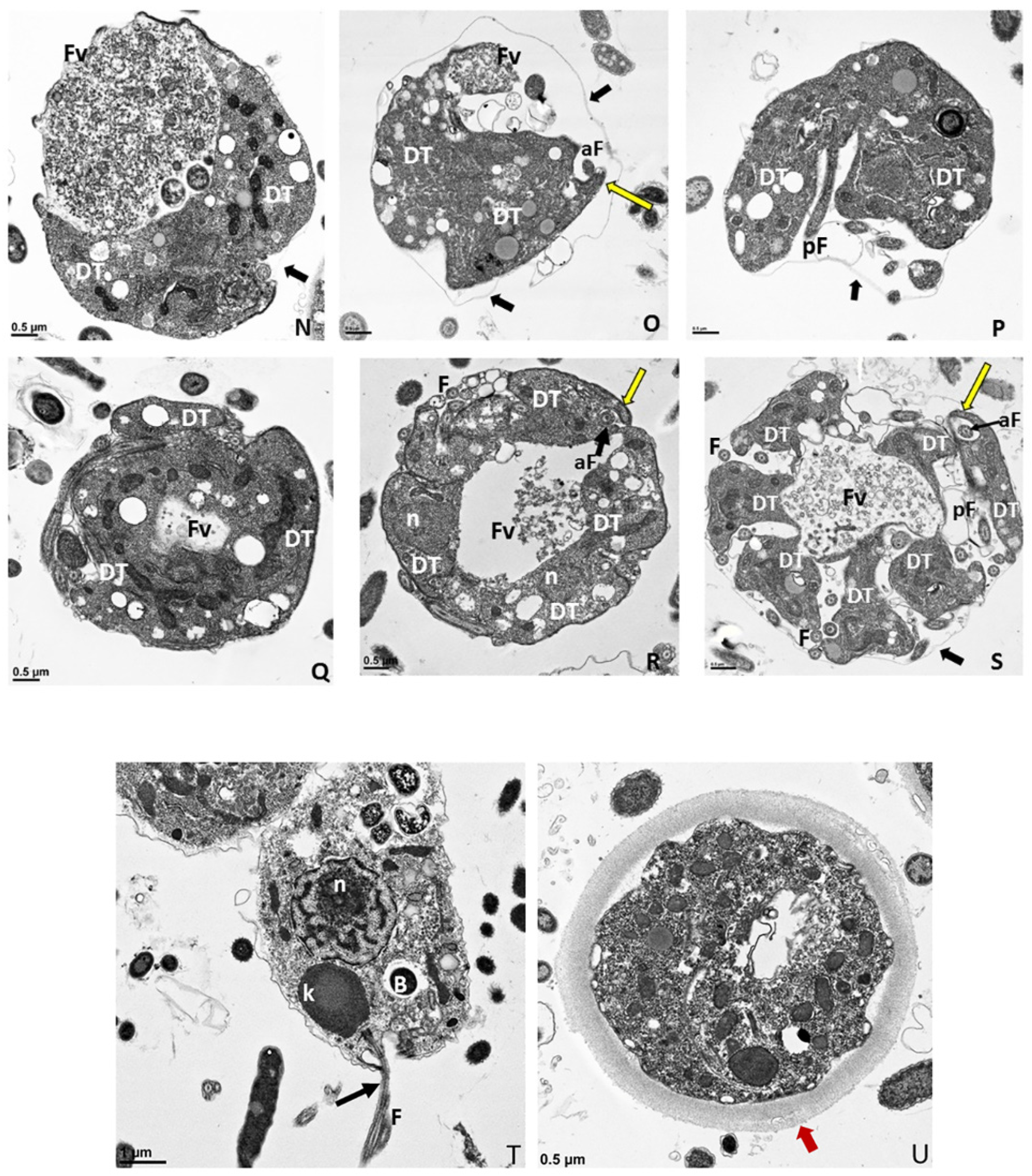
Figure 6.
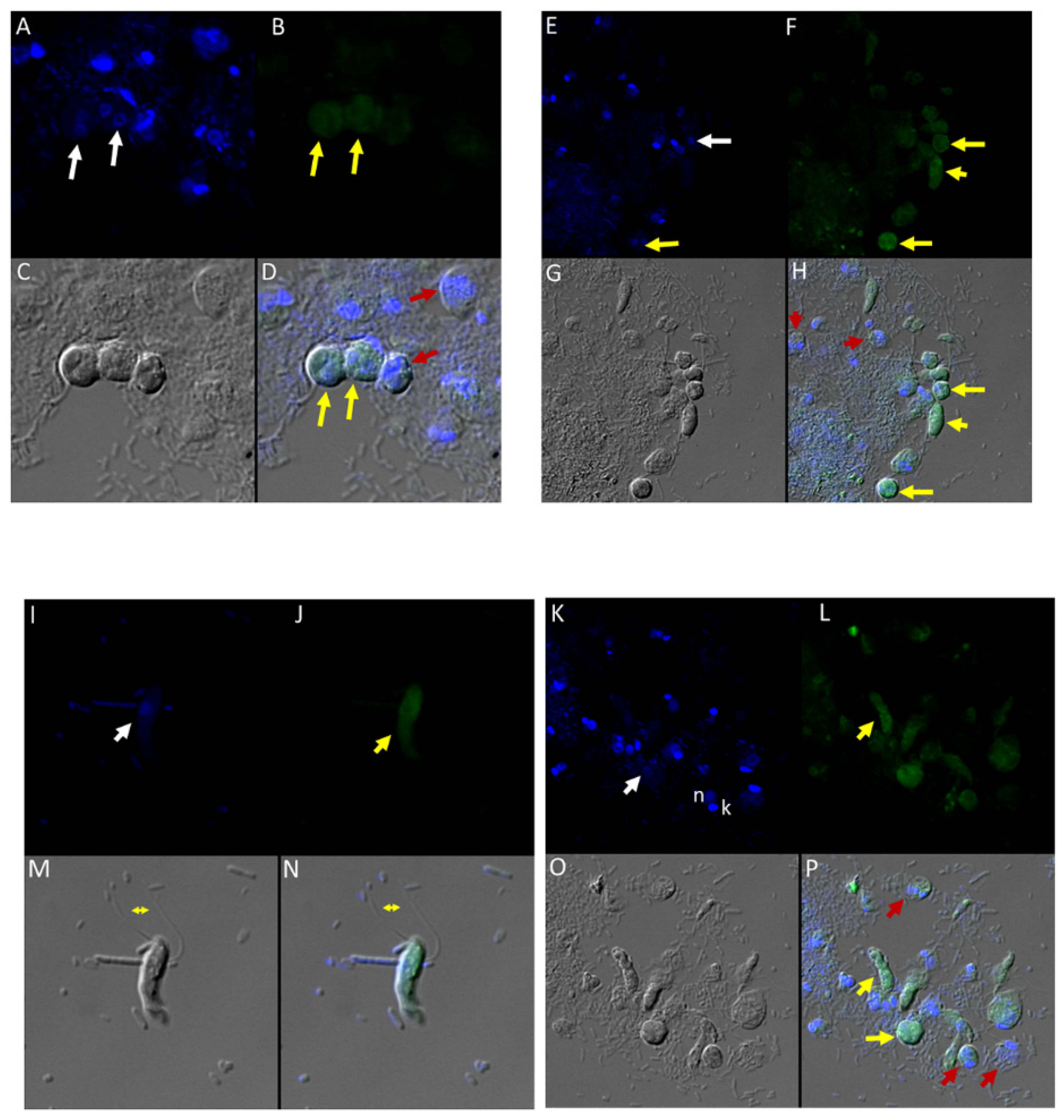
(A–G): Transmission electron micrographs of Colpodella sp. (ATCC 50594) life cycle stages. (A,B), Trophozoites. (C), attachment (D, enlarged) of Colpodella sp. (ATCC 50594) trophozoite (red arrow) to P. caudatus showing two points of attachment to the plasma membrane of P. caudatus (yellow arrow). Microtubules and organelles are shown at the point of attachment. (E), (enlarged F) microtubules spread out at point of attachment. (G), opening shown with cytoplasmic contents of prey aspirated into Colpodella sp. (ATCC 50594) predator. Open arrows show direction of flow of cytoplasmic contents. Bacteria (B) in prey were identified. Blue arrows shown points of attachment in predator-prey pairs undergoing myzocytosis. Anterior flagella (aF), developing trophozoite (DT), flagella (F), food vacuole (Fv), nucleus (n), pseudo-conoid (PC), posterior flagella (pF). Scale bars, (A) (2 µm), (B) (1 µm), (C) (2 µm), (D) (500 nm), (E) (1 µm), (F) (500 nm), and (G) (1 µm). (H–M): Transmission electron micrographs of Colpodella sp. (ATCC 50594) life cycle stages. (H) (enlarged in I) shows attachment of Colpodella sp. (ATCC 50594) trophozoites (yellow arrow) to P. caudatus (red arrow) containing rod shaped bacteria in the cytoplasm. Blue arrow shows point of attachment between predator and prey. The plasma membrane of Colpodella sp. (ATCC 50594) is seen surrounding the plasma membrane of P. caudatus (three small black arrows) being pulled into the cytoplasm of Colpodella sp. (ATCC 50594). Open white arrows show the direction of flow of the cytoplasmic contents from the prey. (J), (enlarged in K) shows the posterior food vacuole of Colpodella sp. (ATCC 50594) enlarged after feeding accompanied by disintegration of the anterior end with loss of flagella and apical organelles. (L), young cyst of Colpodella sp. (ATCC 50594) with developing trophozoite (DT) and remnant food vacuole. M, young cyst with developing trophozoite and remnant food vacuole. Anterior flagella (aF), developing trophozoite (DT), flagella (F), food vacuole (Fv), mitochondria (m), nucleus (n), pseudo-conoid (PC), posterior flagella (pF). Scale bars, (H) (2 µm), (I) (200 nm), (J) (2 µm), (K) (1 µm), (L,M) (0.5 µm). (N–S): Transmission electron micrographs of Colpodella sp. (ATCC 50594) life cycle stages. N-P, shows Colpodella sp. (ATCC 50594) mature cysts containing two developing trophozoites with remnant food vacuoles (N,O), black arrows identify the thin cyst wall and the developing pseudo-conoid (yellow arrow) was identified in a developing trophozoite. (Q–S) show different number of developing trophozoites in cysts of Colpodella sp. (ATCC 50594). (Q), cyst with three DT, (R), cyst with four DT and (S), cyst with seven DT. One trophozoite is more mature than the other six, and contains a developed pseudo-conoid (yellow arrow), in an asymmetric and asynchronous development of the developing trophozoites. Remnant food vacuoles were identified in panels Q-S. Black arrows identify the thin cyst wall of Colpodella sp. (ATCC 50594) cysts. Anterior flagella (aF), developing trophozoite (DT), flagella (F), food vacuole (Fv), mitochondria (m), nucleus (n), pseudo-conoid (PC), posterior flagella (pF). Scale bars, N to S (0.5 µm). (T–U): (T), Trophozoite of P. caudatus showing the nucleus (n), kinetoplast (k), flagella (F) and black arrow is paraflagellar rod. (U), Cyst of P. caudatus showing the thick cyst wall (red arrowhead). Bacteria (B) were identified in the cytoplasm of P. caudatus. Scale bars, (T) (1 µm) and (U) (0.5 µm).
Figure 6.
(A–G): Transmission electron micrographs of Colpodella sp. (ATCC 50594) life cycle stages. (A,B), Trophozoites. (C), attachment (D, enlarged) of Colpodella sp. (ATCC 50594) trophozoite (red arrow) to P. caudatus showing two points of attachment to the plasma membrane of P. caudatus (yellow arrow). Microtubules and organelles are shown at the point of attachment. (E), (enlarged F) microtubules spread out at point of attachment. (G), opening shown with cytoplasmic contents of prey aspirated into Colpodella sp. (ATCC 50594) predator. Open arrows show direction of flow of cytoplasmic contents. Bacteria (B) in prey were identified. Blue arrows shown points of attachment in predator-prey pairs undergoing myzocytosis. Anterior flagella (aF), developing trophozoite (DT), flagella (F), food vacuole (Fv), nucleus (n), pseudo-conoid (PC), posterior flagella (pF). Scale bars, (A) (2 µm), (B) (1 µm), (C) (2 µm), (D) (500 nm), (E) (1 µm), (F) (500 nm), and (G) (1 µm). (H–M): Transmission electron micrographs of Colpodella sp. (ATCC 50594) life cycle stages. (H) (enlarged in I) shows attachment of Colpodella sp. (ATCC 50594) trophozoites (yellow arrow) to P. caudatus (red arrow) containing rod shaped bacteria in the cytoplasm. Blue arrow shows point of attachment between predator and prey. The plasma membrane of Colpodella sp. (ATCC 50594) is seen surrounding the plasma membrane of P. caudatus (three small black arrows) being pulled into the cytoplasm of Colpodella sp. (ATCC 50594). Open white arrows show the direction of flow of the cytoplasmic contents from the prey. (J), (enlarged in K) shows the posterior food vacuole of Colpodella sp. (ATCC 50594) enlarged after feeding accompanied by disintegration of the anterior end with loss of flagella and apical organelles. (L), young cyst of Colpodella sp. (ATCC 50594) with developing trophozoite (DT) and remnant food vacuole. M, young cyst with developing trophozoite and remnant food vacuole. Anterior flagella (aF), developing trophozoite (DT), flagella (F), food vacuole (Fv), mitochondria (m), nucleus (n), pseudo-conoid (PC), posterior flagella (pF). Scale bars, (H) (2 µm), (I) (200 nm), (J) (2 µm), (K) (1 µm), (L,M) (0.5 µm). (N–S): Transmission electron micrographs of Colpodella sp. (ATCC 50594) life cycle stages. N-P, shows Colpodella sp. (ATCC 50594) mature cysts containing two developing trophozoites with remnant food vacuoles (N,O), black arrows identify the thin cyst wall and the developing pseudo-conoid (yellow arrow) was identified in a developing trophozoite. (Q–S) show different number of developing trophozoites in cysts of Colpodella sp. (ATCC 50594). (Q), cyst with three DT, (R), cyst with four DT and (S), cyst with seven DT. One trophozoite is more mature than the other six, and contains a developed pseudo-conoid (yellow arrow), in an asymmetric and asynchronous development of the developing trophozoites. Remnant food vacuoles were identified in panels Q-S. Black arrows identify the thin cyst wall of Colpodella sp. (ATCC 50594) cysts. Anterior flagella (aF), developing trophozoite (DT), flagella (F), food vacuole (Fv), mitochondria (m), nucleus (n), pseudo-conoid (PC), posterior flagella (pF). Scale bars, N to S (0.5 µm). (T–U): (T), Trophozoite of P. caudatus showing the nucleus (n), kinetoplast (k), flagella (F) and black arrow is paraflagellar rod. (U), Cyst of P. caudatus showing the thick cyst wall (red arrowhead). Bacteria (B) were identified in the cytoplasm of P. caudatus. Scale bars, (T) (1 µm) and (U) (0.5 µm).
![Tropicalmed 06 00127 g006a]()
![Tropicalmed 06 00127 g006b]()
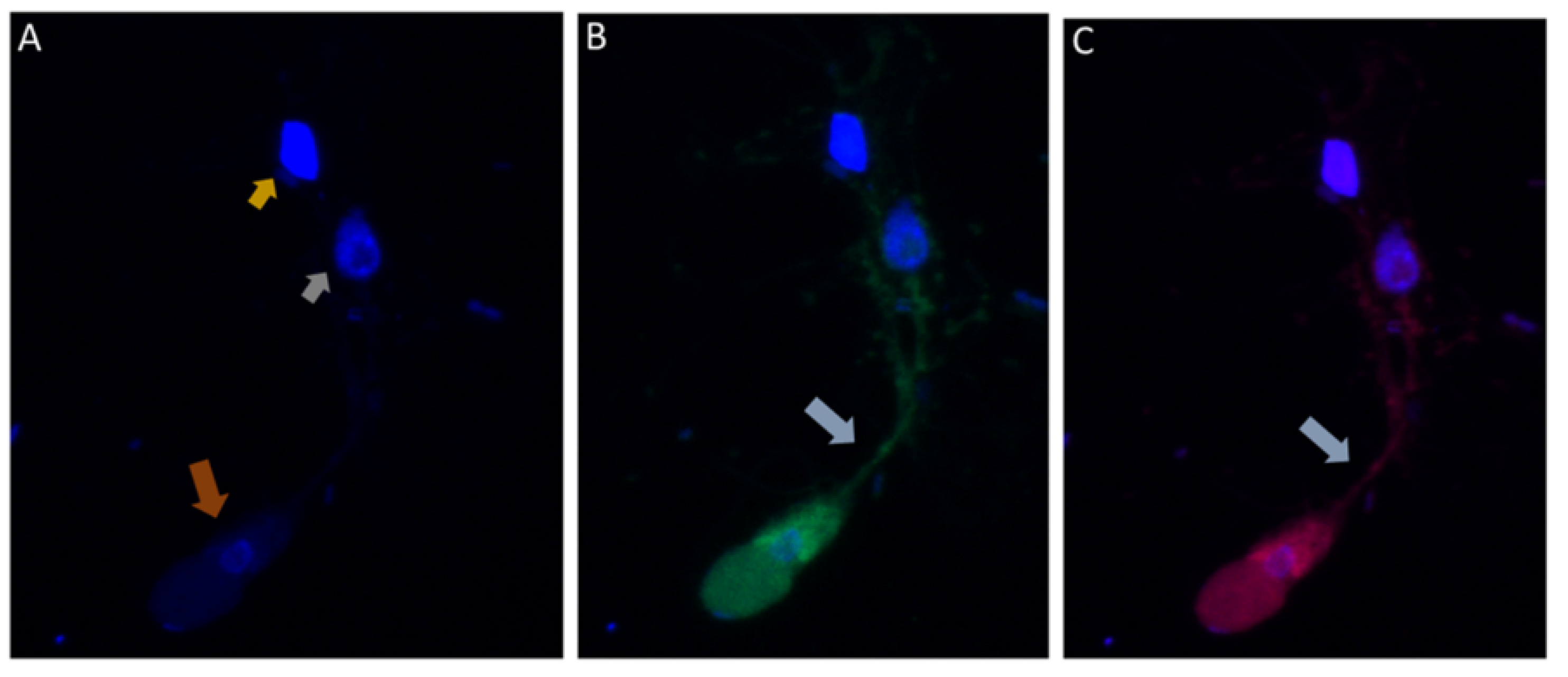
Figure 7.
(
A–
E): Immunofluorescence microscopy of
Colpodella sp. (ATCC 50594) trophozoite in myzocytosis using antiserum 686 (1:100 dilution) specific for
P. falciparum rhoptry protein, RhopH3 (green) and antiserum FL (1:100 dilution) specific for
P. berghei RhopH3 (red). DAPI and DIC images are also shown. (
A). DAPI staining showing nucleus of
Colpodella sp. trophozoite (red arrow), nucleus (grey arrowhead) and kinetoplast (gold arrowhead) of
P. caudatus trophozoite. (
B). Antiserum 686 cross reacted with
Colpodella sp. (ATCC 50594) proteins in the cytoplasm and in the tubular tether used for attachment with the prey. (
C). Antiserum FL cross reacted with
Colpodella sp. (ATCC 50594) proteins in the cytoplasm and in the tubular tether used for attachment with the prey and is colocalized with antiserum 686. (
D). merge of DAPI, 686 and FL showing antibody colocalization of antibody reactivity. (
E). merge of DAPI, 686, FL and DIC showing antibody reactivity and morphology of
Colpodella sp. (ATCC 50594) and
P. caudatus trophozoites in myzocytosis. Antibody reactivity is seen on the tubular tether and within the
P. caudatus prey in the section closest to the predator (movie in
Supplementary Figure S2).
Figure 7.
(
A–
E): Immunofluorescence microscopy of
Colpodella sp. (ATCC 50594) trophozoite in myzocytosis using antiserum 686 (1:100 dilution) specific for
P. falciparum rhoptry protein, RhopH3 (green) and antiserum FL (1:100 dilution) specific for
P. berghei RhopH3 (red). DAPI and DIC images are also shown. (
A). DAPI staining showing nucleus of
Colpodella sp. trophozoite (red arrow), nucleus (grey arrowhead) and kinetoplast (gold arrowhead) of
P. caudatus trophozoite. (
B). Antiserum 686 cross reacted with
Colpodella sp. (ATCC 50594) proteins in the cytoplasm and in the tubular tether used for attachment with the prey. (
C). Antiserum FL cross reacted with
Colpodella sp. (ATCC 50594) proteins in the cytoplasm and in the tubular tether used for attachment with the prey and is colocalized with antiserum 686. (
D). merge of DAPI, 686 and FL showing antibody colocalization of antibody reactivity. (
E). merge of DAPI, 686, FL and DIC showing antibody reactivity and morphology of
Colpodella sp. (ATCC 50594) and
P. caudatus trophozoites in myzocytosis. Antibody reactivity is seen on the tubular tether and within the
P. caudatus prey in the section closest to the predator (movie in
Supplementary Figure S2).
![Tropicalmed 06 00127 g007a]()
![Tropicalmed 06 00127 g007b]()
Figure 8.
(A–H): Immunofluorescence microscopy of Colpodella sp. (ATCC 50594) trophozoites and cysts using anti-IMC3 antibody. (A,E); two-nuclei cysts of Colpodella sp. (ATCC 50594) were identified by DAPI staining (white arrows, panel A), single nucleus cyst is shown in panel (E) (white arrow). (B,F); Antibody reactivity was observed on cysts (yellow arrows) and trophozoites (yellow arrowheads) of Colpodella sp. (ATCC 50594). (C,G); DIC microscopy showing the morphology or trophozoites and cysts of predator and prey. (D,H); Merged antibody, DAPI and DIC images showing no reactivity of anti-IMC3 antibodies with P. caudatus cysts (red arrows, panel D) and trophozoites (red arrowheads, panel H), antibody reactivity with Colpodella sp. (ATCC 50594) cysts (panels D and H, yellow arrows). (I–P): Immunofluorescence microscopy of Colpodella sp. (ATCC 50594) trophozoites and cysts using anti-IMC3 FLR antibody. (I,K); DAPI stained nuclei of Colpodella sp. (ATCC 50594) trophozoites (white arrows, panels I and K) and P. caudatus trophozoites showing nucleus (n) and kinetoplast (k) (panel K). (J,L); anti-IMC3 FLR antibody reactivity is shown on trophozoites and cysts of Colpodella sp. (ATCC 50594) (yellow arrowheads and arrow). (N,P); Merging of antibody, DAPI and DIC image. Antibody reactivity was not observed on P. caudatus trophozoites (red arrowheads, panel P) while ant-IMC3 FLR reactivity was observed on trophozoites and cysts of Colpodella sp. (ATCC 50594) (yellow arrow and arrowhead). Flagella of Colpodella sp. (ATCC 50594) were identified (double arrow, panels M and N).
Figure 8.
(A–H): Immunofluorescence microscopy of Colpodella sp. (ATCC 50594) trophozoites and cysts using anti-IMC3 antibody. (A,E); two-nuclei cysts of Colpodella sp. (ATCC 50594) were identified by DAPI staining (white arrows, panel A), single nucleus cyst is shown in panel (E) (white arrow). (B,F); Antibody reactivity was observed on cysts (yellow arrows) and trophozoites (yellow arrowheads) of Colpodella sp. (ATCC 50594). (C,G); DIC microscopy showing the morphology or trophozoites and cysts of predator and prey. (D,H); Merged antibody, DAPI and DIC images showing no reactivity of anti-IMC3 antibodies with P. caudatus cysts (red arrows, panel D) and trophozoites (red arrowheads, panel H), antibody reactivity with Colpodella sp. (ATCC 50594) cysts (panels D and H, yellow arrows). (I–P): Immunofluorescence microscopy of Colpodella sp. (ATCC 50594) trophozoites and cysts using anti-IMC3 FLR antibody. (I,K); DAPI stained nuclei of Colpodella sp. (ATCC 50594) trophozoites (white arrows, panels I and K) and P. caudatus trophozoites showing nucleus (n) and kinetoplast (k) (panel K). (J,L); anti-IMC3 FLR antibody reactivity is shown on trophozoites and cysts of Colpodella sp. (ATCC 50594) (yellow arrowheads and arrow). (N,P); Merging of antibody, DAPI and DIC image. Antibody reactivity was not observed on P. caudatus trophozoites (red arrowheads, panel P) while ant-IMC3 FLR reactivity was observed on trophozoites and cysts of Colpodella sp. (ATCC 50594) (yellow arrow and arrowhead). Flagella of Colpodella sp. (ATCC 50594) were identified (double arrow, panels M and N).
![Tropicalmed 06 00127 g008]()
Figure 9.
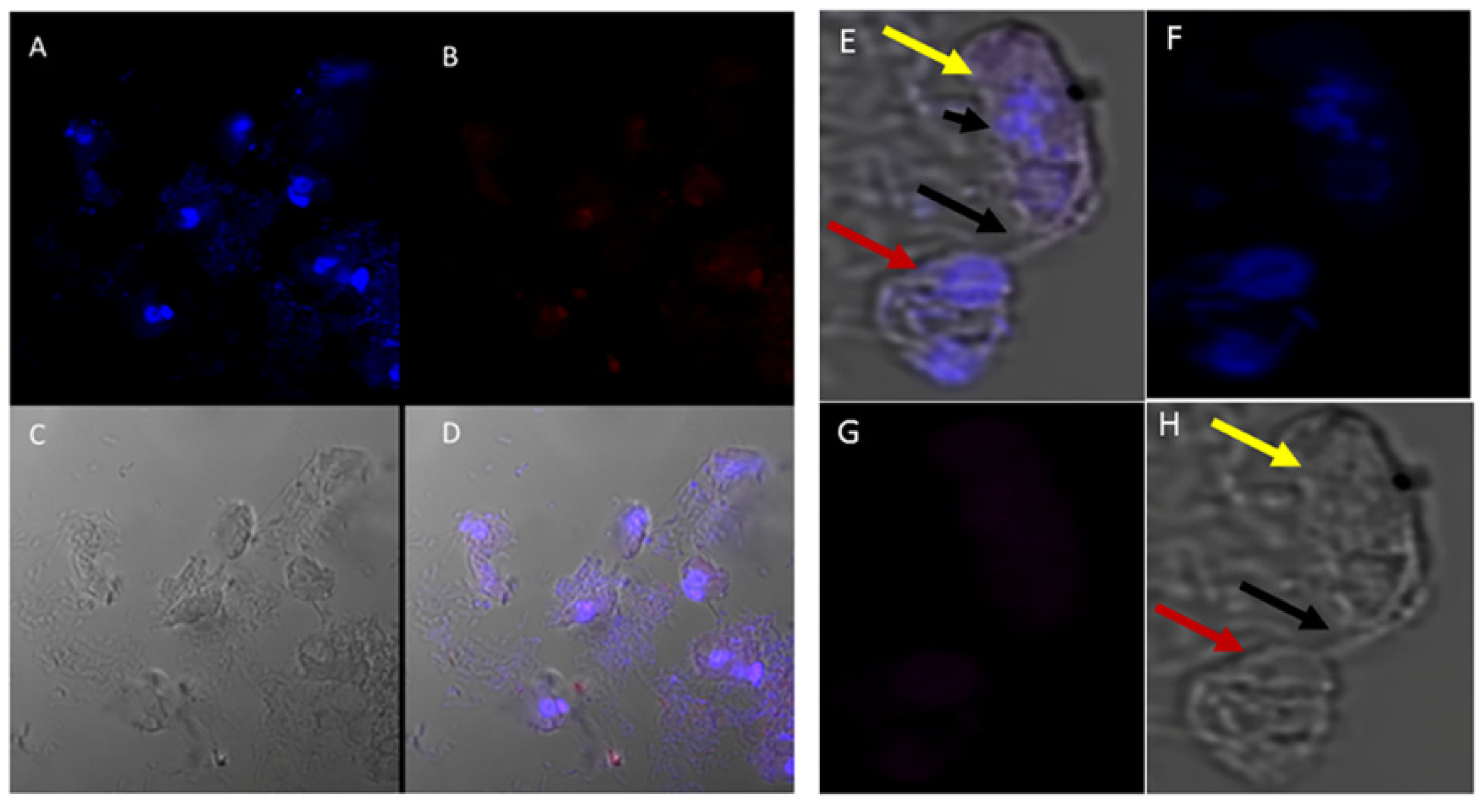
(A–H): Immunofluorescence microscopy of Colpodella sp. (ATCC 50594) trophozoites and cysts using anti-IMC7 and anti-Py 235 antibodies. (A), DAPI stained nuclei of Colpodella sp. (ATCC 50594) and P. caudatus, (B), Background to no reactivity was observed with anti-IMC7. Panels (C,D) shown DIC microscopy and merged Antibody, DAPI and DIC images, respectively. (E), Merged anti-Py235 antibody, DAPI, DIC image of Colpodella sp. (ATCC 50594) trophozoite (yellow arrow) and P. caudatus (red arrow) in myzocytosis. The tubular tether (black arrow) joining predator and prey is shown and DAPI stained aspirated cytoplasmic contents from the prey is shown (black arrowhead). (G), no antibody reactivity was observed and (H), DIC microscopy of Colpodella sp. (ATCC 50594) trophozoite and P. caudatus.
Figure 9.
(A–H): Immunofluorescence microscopy of Colpodella sp. (ATCC 50594) trophozoites and cysts using anti-IMC7 and anti-Py 235 antibodies. (A), DAPI stained nuclei of Colpodella sp. (ATCC 50594) and P. caudatus, (B), Background to no reactivity was observed with anti-IMC7. Panels (C,D) shown DIC microscopy and merged Antibody, DAPI and DIC images, respectively. (E), Merged anti-Py235 antibody, DAPI, DIC image of Colpodella sp. (ATCC 50594) trophozoite (yellow arrow) and P. caudatus (red arrow) in myzocytosis. The tubular tether (black arrow) joining predator and prey is shown and DAPI stained aspirated cytoplasmic contents from the prey is shown (black arrowhead). (G), no antibody reactivity was observed and (H), DIC microscopy of Colpodella sp. (ATCC 50594) trophozoite and P. caudatus.
Figure 10.
(A–H): Immunofluorescence microscopy of Colpodella sp. (ATCC 50594) trophozoites and cysts using anti-AMA-1antibodies. (A,E); DAPI stained nuclei of Colpodella sp. (ATCC 50594) (white arrow) and P. caudatus (red arrow) showing DAPI stained nucleus (n) and kinetoplast (k). (B,F) shows antibody reactivity with Colpodella sp. (ATCC 50594) trophozoites (yellow arrow) with intense antibody reactivity at the anterior end of the trophozoites (yellow arrowhead), panels (B,D,F,H). No antibody reactivity was observed on P. caudatus trophozoites (red arrows). (C,G); DIC microscopy shown the morphology of Colpodella sp. (ATCC 50594) and P. caudatus. (D,H); shows merged images of Antibody, DAPI and DIC. Grey arrowhead identified DAPI stained aspirated cytoplasmic contents of P. caudatus.
Figure 10.
(A–H): Immunofluorescence microscopy of Colpodella sp. (ATCC 50594) trophozoites and cysts using anti-AMA-1antibodies. (A,E); DAPI stained nuclei of Colpodella sp. (ATCC 50594) (white arrow) and P. caudatus (red arrow) showing DAPI stained nucleus (n) and kinetoplast (k). (B,F) shows antibody reactivity with Colpodella sp. (ATCC 50594) trophozoites (yellow arrow) with intense antibody reactivity at the anterior end of the trophozoites (yellow arrowhead), panels (B,D,F,H). No antibody reactivity was observed on P. caudatus trophozoites (red arrows). (C,G); DIC microscopy shown the morphology of Colpodella sp. (ATCC 50594) and P. caudatus. (D,H); shows merged images of Antibody, DAPI and DIC. Grey arrowhead identified DAPI stained aspirated cytoplasmic contents of P. caudatus.
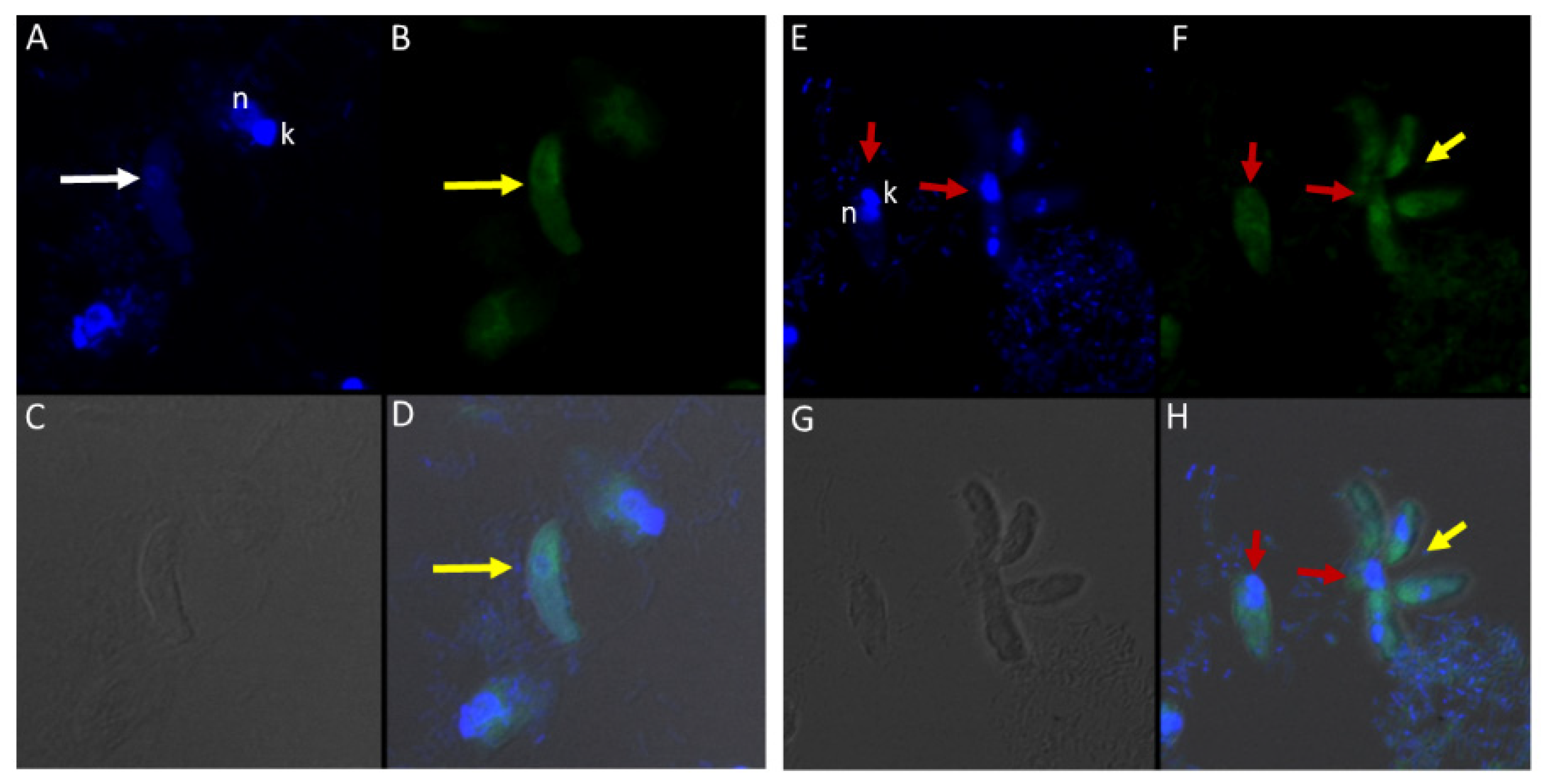
Figure 11.
(A–H): Immunofluorescence microscopy of Colpodella sp. (ATCC 50594) trophozoites and cysts using anti-EBA175 antibodies. (A,E); DAPI stained nuclei of Colpodella sp. (ATCC 50594) (white arrow) and P. caudatus trophozoites (red arrow) showing nucleus (n) and kinetoplast (k). (B,F); anti-EBA175 reactivity with single Colpodella sp. trophozoite and multiple trophozoites feeding on a single P. caudatus. Yellow arrowhead shows cluster of four Colpodella sp. (ATCC 50594) trophozoites. (C,G); show DIC microscopy of Colpodella sp. (ATCC 50594) and P. caudatus trophozoites. (D,H); Merged image of anti-EBA175, DAPI and DIC.
Figure 11.
(A–H): Immunofluorescence microscopy of Colpodella sp. (ATCC 50594) trophozoites and cysts using anti-EBA175 antibodies. (A,E); DAPI stained nuclei of Colpodella sp. (ATCC 50594) (white arrow) and P. caudatus trophozoites (red arrow) showing nucleus (n) and kinetoplast (k). (B,F); anti-EBA175 reactivity with single Colpodella sp. trophozoite and multiple trophozoites feeding on a single P. caudatus. Yellow arrowhead shows cluster of four Colpodella sp. (ATCC 50594) trophozoites. (C,G); show DIC microscopy of Colpodella sp. (ATCC 50594) and P. caudatus trophozoites. (D,H); Merged image of anti-EBA175, DAPI and DIC.
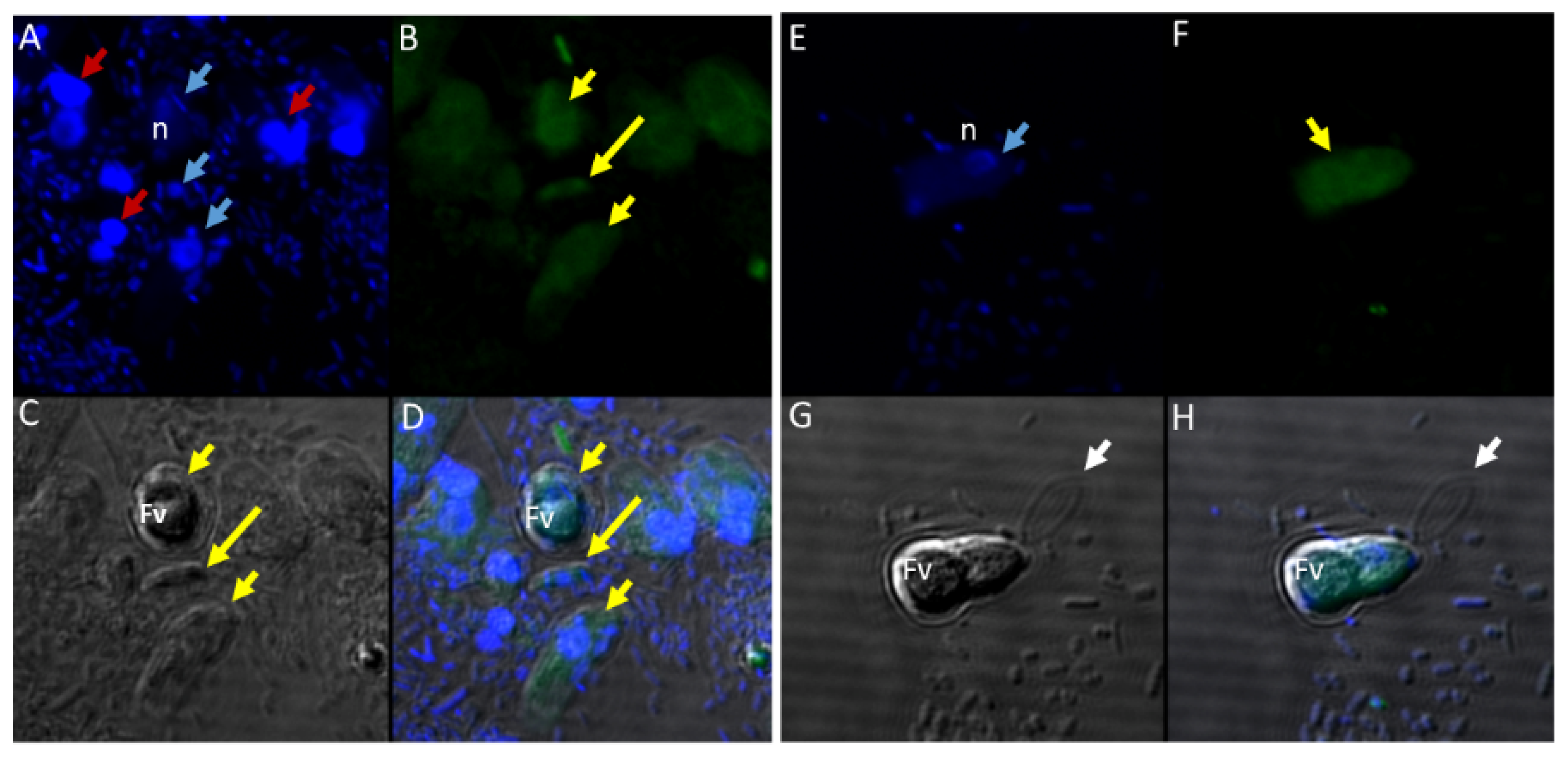
Figure 12.
(A–H): Immunofluorescence microscopy of Colpodella sp. (ATCC 50594) trophozoites and cysts using anti-Plasmepsin II antibodies. (A,E); DAPI stained nuclei of Colpodella sp. (ATCC 50594) pre-cysts (blue arrowheads) and P. caudatus trophozoites (red arrowheads). (B,F); anti-plasmepsin II antibody reactivity was observed with pre-cysts of Colpodella sp. (ATCC 50594) with enlarged food vacuoles (Fv) after myzocytosis. Reactivity was seen in the cytoplasm at the anterior end and (F,C,G); DIC microscopy showing morphology of Colpodella sp. (ATCC 50594) and P. caudatus. (D,H); Merged image of anti-plasmepsin II, DAPI and DIC microscopy. Yellow arrows identified a young trophozoite that reacted with anti-plasmepsin II. Some background antibody reactivity was observed on some P. caudatus trophozoites.
Figure 12.
(A–H): Immunofluorescence microscopy of Colpodella sp. (ATCC 50594) trophozoites and cysts using anti-Plasmepsin II antibodies. (A,E); DAPI stained nuclei of Colpodella sp. (ATCC 50594) pre-cysts (blue arrowheads) and P. caudatus trophozoites (red arrowheads). (B,F); anti-plasmepsin II antibody reactivity was observed with pre-cysts of Colpodella sp. (ATCC 50594) with enlarged food vacuoles (Fv) after myzocytosis. Reactivity was seen in the cytoplasm at the anterior end and (F,C,G); DIC microscopy showing morphology of Colpodella sp. (ATCC 50594) and P. caudatus. (D,H); Merged image of anti-plasmepsin II, DAPI and DIC microscopy. Yellow arrows identified a young trophozoite that reacted with anti-plasmepsin II. Some background antibody reactivity was observed on some P. caudatus trophozoites.
Figure 13.
(A–H): Normal and rabbit serum negative controls. (A–D), normal mouse serum 1:100. There was no reactivity with proteins of Colpodella sp. (ATCC 50594) and P. caudatus. (A), merged image showing a Colpodella sp. (ATCC 50594) trophozoite (yellow arrow) attached to a P. caudatus trophozoite (red arrow). (B). DAPI staining shows the nuclei in both cells. The DAPI stained cytoplasmic contents aspirated from the prey can be visualized as blue in the posterior food vacuole of Colpodella sp. (ATCC 50594) (grey arrowhead). (C). There was no reactivity with NMS. (D). DIC showing the morphology of the cells in myzocytosis. (E–H), Normal rabbit serum, 1:100 was used as a negative control. (E), merged image of antibody, DAPI and DIC showing no reactivity with proteins of Colpodella sp. (ATCC 50594) and P. caudatus. The merged image shows two separate attachments of Colpodella sp. (ATCC 50594) and P. caudatus trophozoites. (F), DAPI staining shows the nucleus of Colpodella sp. (ATCC 50594) and P. caudatus. (C). Background to no reactivity was observed on cells with NRS. (D). DIC images show the attachment of Colpodella sp. (ATCC 50594) and the tubular tethers (black arrows).
Figure 13.
(A–H): Normal and rabbit serum negative controls. (A–D), normal mouse serum 1:100. There was no reactivity with proteins of Colpodella sp. (ATCC 50594) and P. caudatus. (A), merged image showing a Colpodella sp. (ATCC 50594) trophozoite (yellow arrow) attached to a P. caudatus trophozoite (red arrow). (B). DAPI staining shows the nuclei in both cells. The DAPI stained cytoplasmic contents aspirated from the prey can be visualized as blue in the posterior food vacuole of Colpodella sp. (ATCC 50594) (grey arrowhead). (C). There was no reactivity with NMS. (D). DIC showing the morphology of the cells in myzocytosis. (E–H), Normal rabbit serum, 1:100 was used as a negative control. (E), merged image of antibody, DAPI and DIC showing no reactivity with proteins of Colpodella sp. (ATCC 50594) and P. caudatus. The merged image shows two separate attachments of Colpodella sp. (ATCC 50594) and P. caudatus trophozoites. (F), DAPI staining shows the nucleus of Colpodella sp. (ATCC 50594) and P. caudatus. (C). Background to no reactivity was observed on cells with NRS. (D). DIC images show the attachment of Colpodella sp. (ATCC 50594) and the tubular tethers (black arrows).
![Tropicalmed 06 00127 g013]()
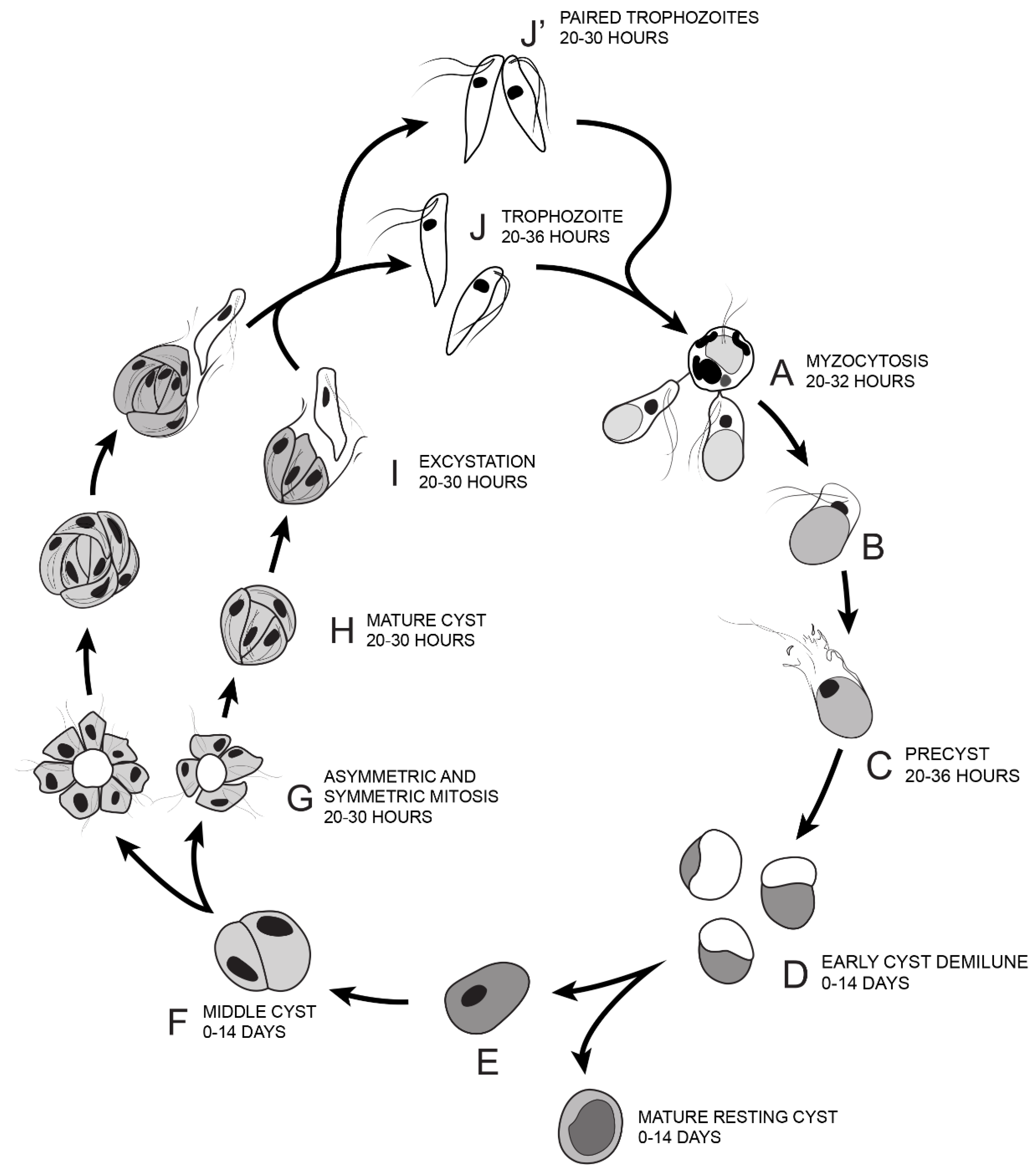
Figure 14.
Illustration of the life cycle of Colpodella sp. (ATCC 50594) cultured in Hay medium. (A). Once trophozoites egress from cysts they begin feeding on P. caudatus prey. Two trophozoites are depicted feeding in the process of myzocytosis. Active cultures with predator-prey attachments are seen 20–32 h after subculture. The posterior food vacuole in Colpodella sp. (ATCC 50594) enlarges during feeding. (B). Encystment begins after feeding. (C). Pre-cyst showing anterior portion of trophozoite with a frayed appearance. Pre-cysts are seen 20–36 h after subculture. (D). Early demilune cysts can be seen in culture during the active phase of the culture or in resting cultures (0–14 days after subculture). (E). Mature cyst stages found in active and resting cultures can be seen in culture 0–14 days after subculture. Mature Colpodella sp. (ATCC 50594) cysts with two or more nuclei are observed in active cultures, as transient cysts are formed at this stage. The one nucleus cyst is predominant in the resting culture. (F). Two-nuclei cysts of Colpodella sp. (ATCC 50594) can be seen in culture, 0–14 days after subculture. (G). Following mitosis, development of the four-nuclei cyst is most common. Asymmetric cysts containing three, five and seven nuclei were observed in culture, in addition to symmetric cysts containing four, six or eight nuclei. Four and seven nuclei cysts are depicted and can be seen in culture 20–30 h after subculture. (H). Mature cysts with young trophozoites can be seen 20–30 h after subculture. (I). Excystation (egress) and release of young trophozoites. (J). Juvenile trophozoites can egress individually at 20–36 h after subculture or rarely, juvenile trophozoites in pairs still attached can egress from cysts and complete cytokinesis outside the cyst (J’ seen 20–30 h after subculture). Paired trophozoites can egress from symmetric or asymmetric cysts. Free swimming young trophozoites egressed from cysts attach to prey to repeat the life cycle. The life cycle of Colpodella sp. (ATCC 50594) lasts 36 h.
Figure 14.
Illustration of the life cycle of Colpodella sp. (ATCC 50594) cultured in Hay medium. (A). Once trophozoites egress from cysts they begin feeding on P. caudatus prey. Two trophozoites are depicted feeding in the process of myzocytosis. Active cultures with predator-prey attachments are seen 20–32 h after subculture. The posterior food vacuole in Colpodella sp. (ATCC 50594) enlarges during feeding. (B). Encystment begins after feeding. (C). Pre-cyst showing anterior portion of trophozoite with a frayed appearance. Pre-cysts are seen 20–36 h after subculture. (D). Early demilune cysts can be seen in culture during the active phase of the culture or in resting cultures (0–14 days after subculture). (E). Mature cyst stages found in active and resting cultures can be seen in culture 0–14 days after subculture. Mature Colpodella sp. (ATCC 50594) cysts with two or more nuclei are observed in active cultures, as transient cysts are formed at this stage. The one nucleus cyst is predominant in the resting culture. (F). Two-nuclei cysts of Colpodella sp. (ATCC 50594) can be seen in culture, 0–14 days after subculture. (G). Following mitosis, development of the four-nuclei cyst is most common. Asymmetric cysts containing three, five and seven nuclei were observed in culture, in addition to symmetric cysts containing four, six or eight nuclei. Four and seven nuclei cysts are depicted and can be seen in culture 20–30 h after subculture. (H). Mature cysts with young trophozoites can be seen 20–30 h after subculture. (I). Excystation (egress) and release of young trophozoites. (J). Juvenile trophozoites can egress individually at 20–36 h after subculture or rarely, juvenile trophozoites in pairs still attached can egress from cysts and complete cytokinesis outside the cyst (J’ seen 20–30 h after subculture). Paired trophozoites can egress from symmetric or asymmetric cysts. Free swimming young trophozoites egressed from cysts attach to prey to repeat the life cycle. The life cycle of Colpodella sp. (ATCC 50594) lasts 36 h.
![Tropicalmed 06 00127 g014]()
Table 1.
Resting cysts of Colpodella sp. (ATCC 50594) and trophozoites on days 5 and 7.
Table 1.
Resting cysts of Colpodella sp. (ATCC 50594) and trophozoites on days 5 and 7.
| Day & Slide # | Early Cyst | Mature Cyst | Colpodella sp. Trophozoite | Parabodo Trophozoite | Colpodella sp. Precyst |
|---|
| Day 5 #1 | 19 | 81 | 0 | 1 | 2 |
| Day 5 #2 | 12 | 88 | 0 | 0 | 1 |
| Day 5 Avg. | 15 | 85 | 0 | 1 | 2 |
| Day 7 #1 | 6 | 94 | 1 | 0 | 0 |
| Day 7 #2 | 9 | 91 | 0 | 2 | 1 |
| Day 7 Avg. | 7 | 93 | 1 | 1 | 1 |
Table 2.
Antibodies used in immunofluorescence and confocal microscopy of Colpodella sp. (ATCC 50594).
Table 2.
Antibodies used in immunofluorescence and confocal microscopy of Colpodella sp. (ATCC 50594).
| Antibodies | Antigen Specificty | Antibody Reactivity |
|---|
| Antiserum 686 | Plasmodium falciparum 110 kDa RhopH3 rhoptry protein | Apical and cytoplasmic reactivity in trophozoites |
| Anti-AMA-1 | P. falciparum apical membrane antigen-1, microneme protein | Apical reactivity in trophozoites |
| Anti-EBA-175 | P. falciparum erythrocyte binding antigen 175 kDa, microneme protein | Apical reactivity in trophozoites |
| Anti-IMC3 | Toxoplasma gondii inner membrane complex protein 3 | Diffuse reactivity on trophozoites and cysts of Colpodella sp. (50594) |
| Anti-IMC3FLR | T. gondii inner membrane complex protein 3 | Diffuse reactivity on trophozoites and cysts of Colpodella sp. (ATCC 50594) |
| Anti-IMC7 | T. gondii inner membrane complex protein 7 | No reactivity |
| Anti-Plasmepsin II | P. falciparum plasmepsin II, food vacuole protein | Diffuse reactivity in food vacuole in pre-cyst |
| Anti-Py235 | P. yoelii 235 kDa rhoptry protein | No reactivity |
| Anti-RhopH3 FL | P. berghei 110 kDa RhopH3 rhoptry protein | Apical and cytoplasmic reactivity in trophozoites |


 ,
, 
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}