Cross-Generational Effects of Heat Stress on Fitness and Wolbachia Density in Aedes aegypti Mosquitoes




Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Mosquito Strains and Colony Maintenance
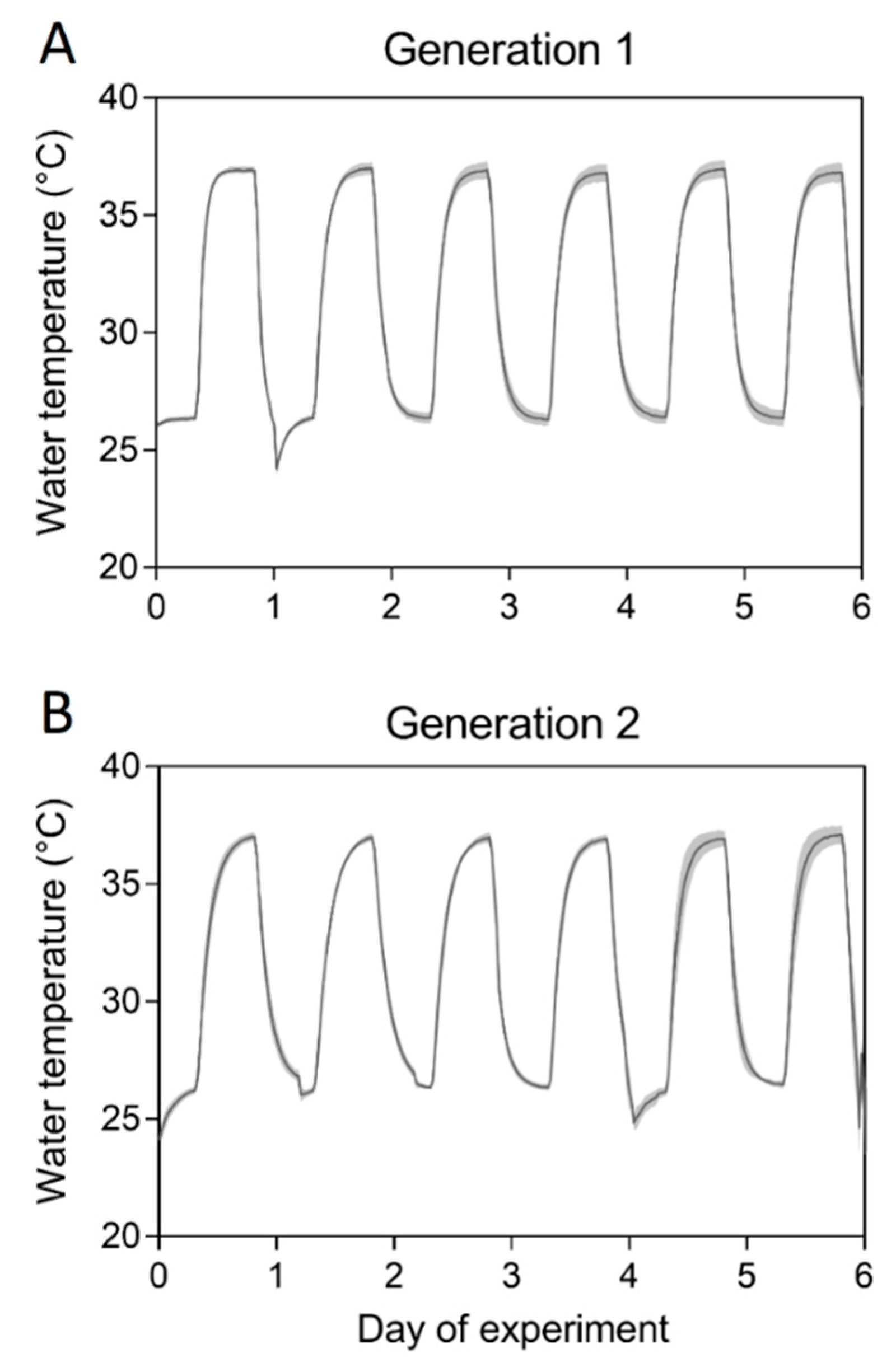
2.3. Mosquito Rearing at Constant and Cyclical Temperatures
2.4. Adult Starvation Tolerance
2.5. Wing Length
2.6. Wolbachia Density
2.7. Statistical Analysis
3. Results
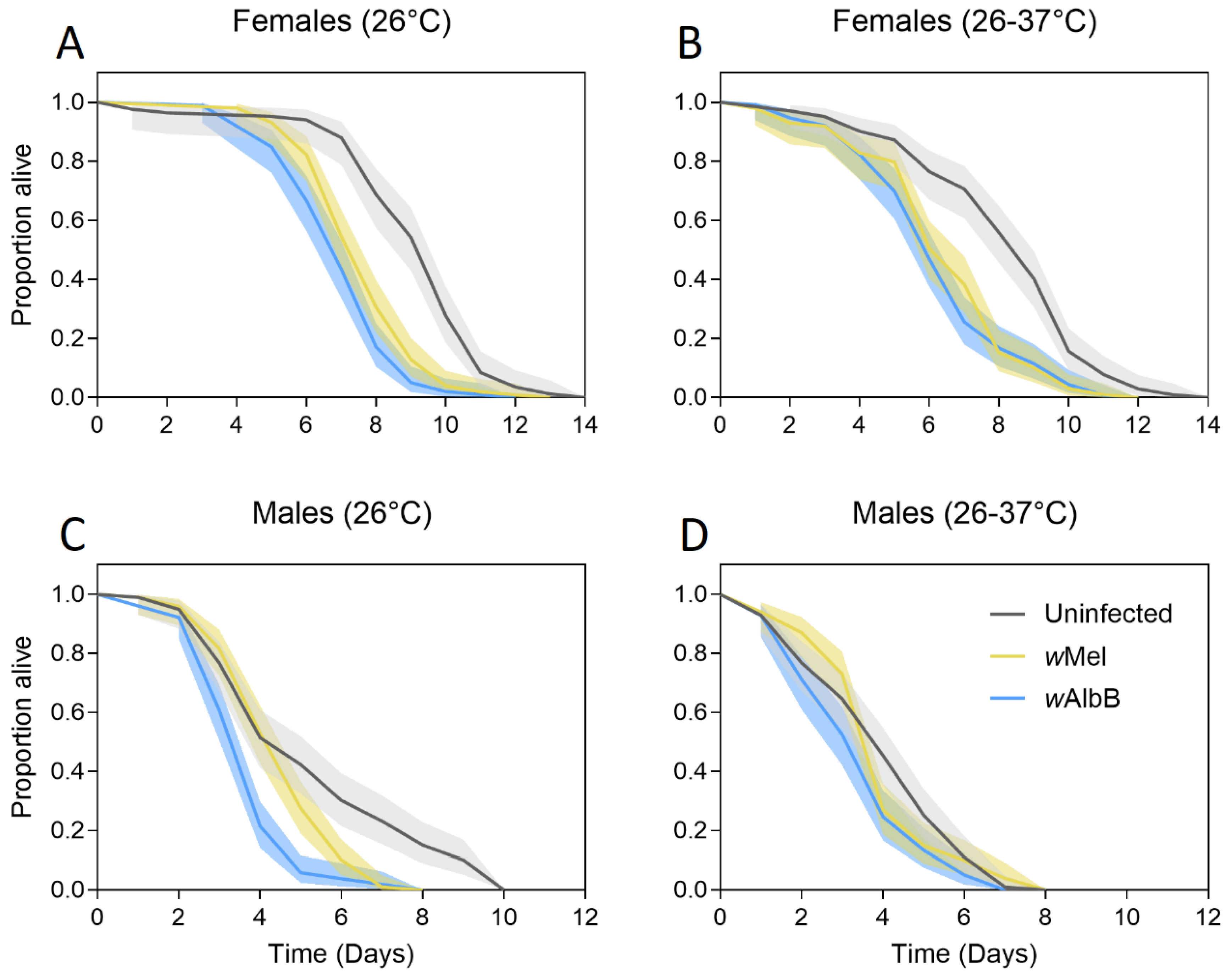
3.1. Adult Starvation Survival
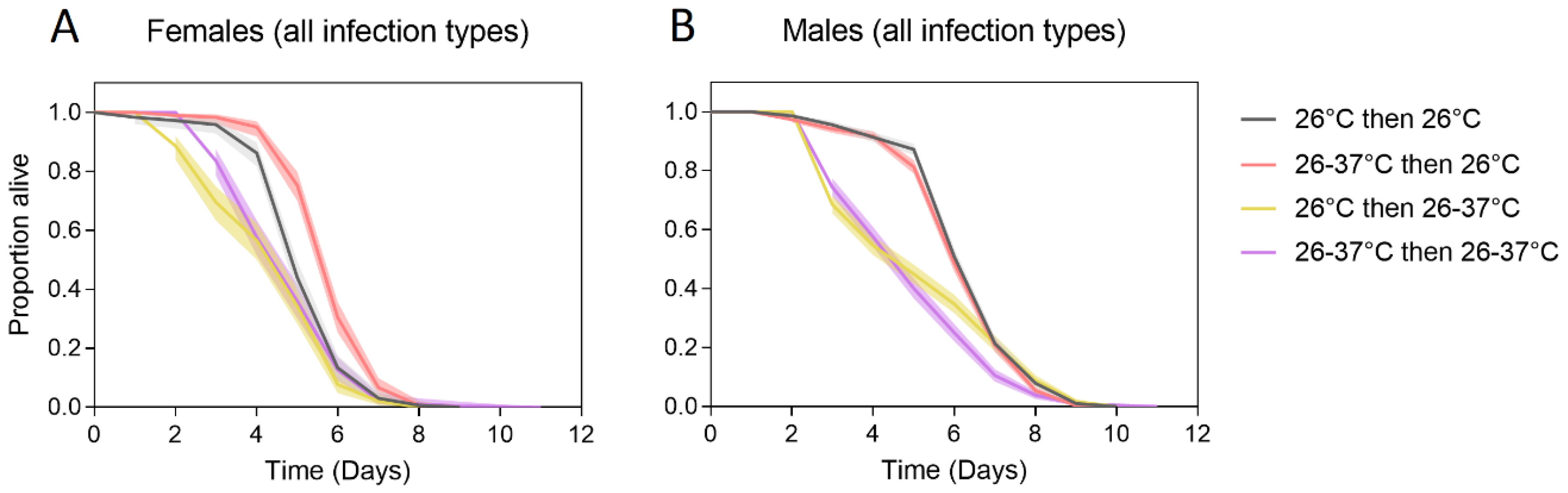
3.1.1. Generation 1
3.1.2. Generation 2
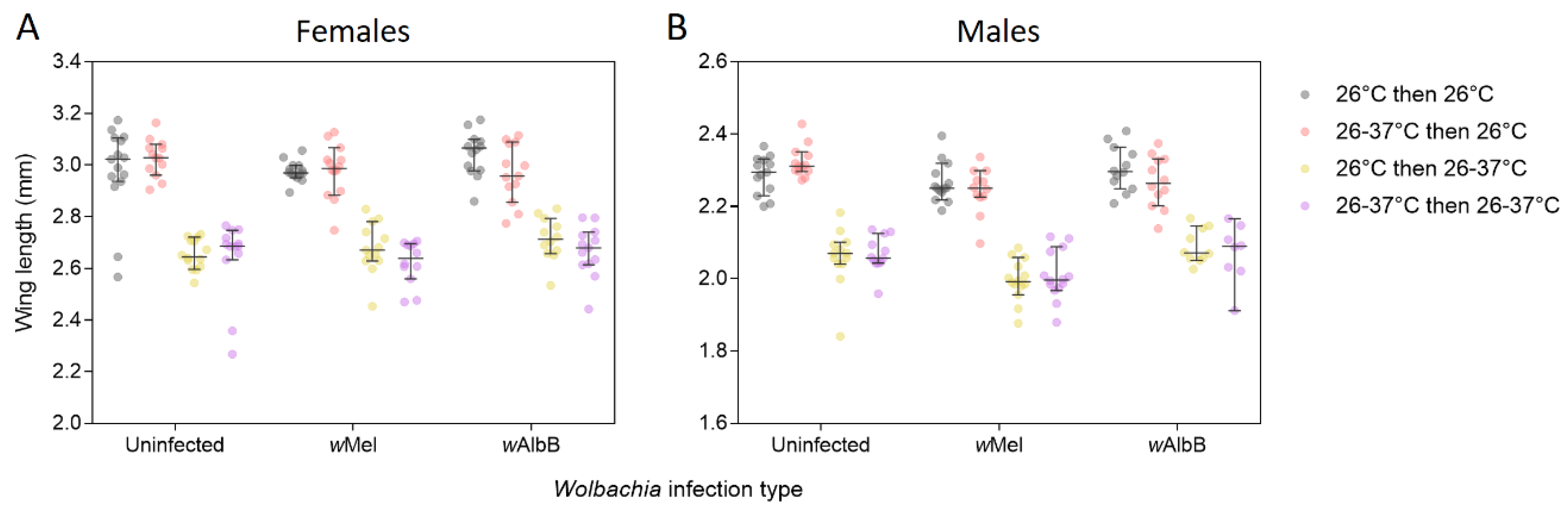
3.2. Wing Length
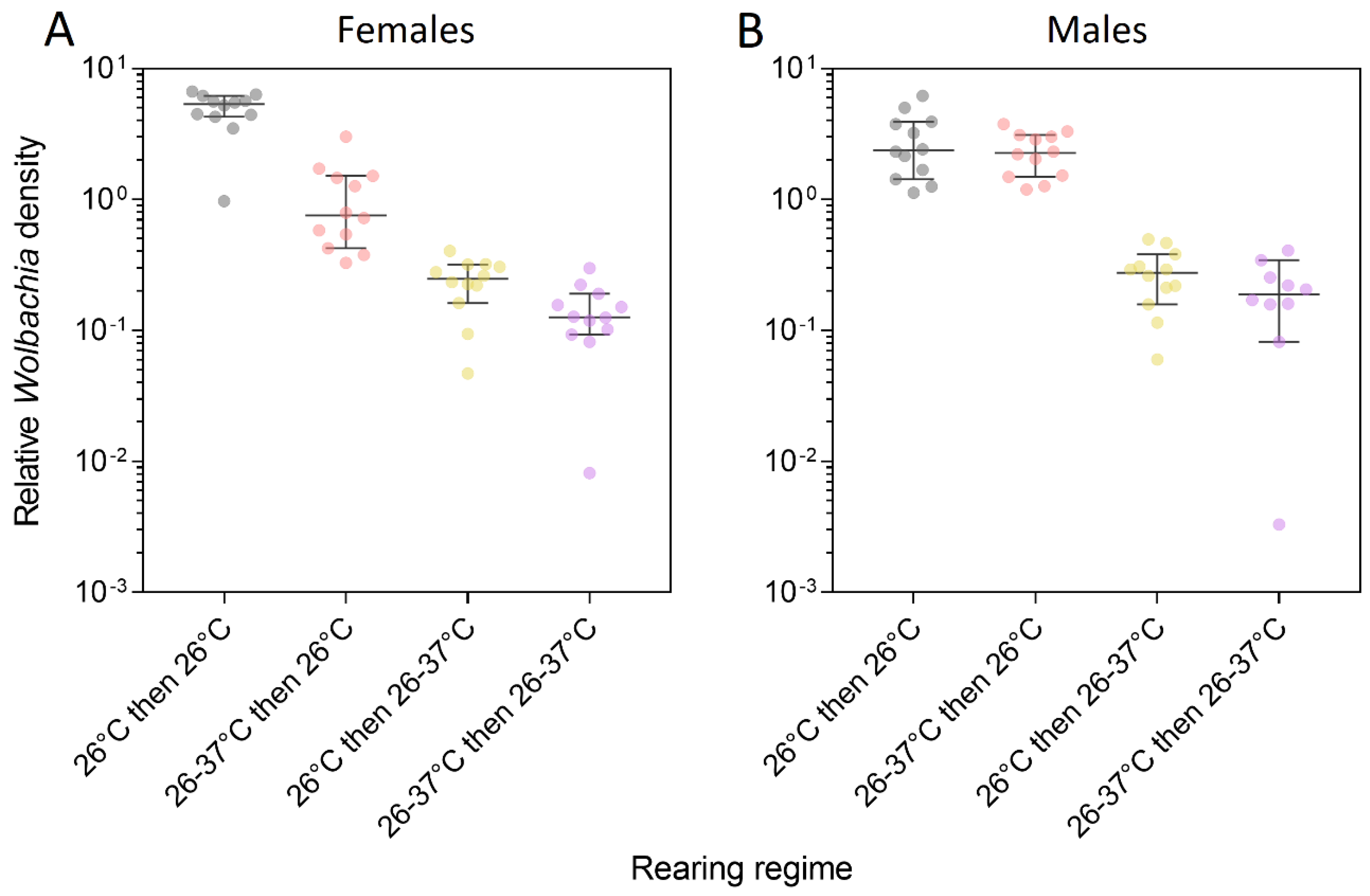
3.3. Wolbachia Density
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

References
- Mackenzie, J.S.; Gubler, D.J.; Petersen, L.R. Emerging flaviviruses: The spread and resurgence of Japanese Encephalitis, West Nile and dengue viruses. Nat. Med. 2004, 10, S98–S109. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Loroño-Pino, M.A.; Cropp, C.B.; Farfán, J.A.; Vorndam, A.V.; Rodríguez-Angulo, E.M.; Rosado-Paredes, E.P.; Flores-Flores, L.F.; Beaty, B.J.; Gubler, D.J. Common occurrence of concurrent infections by multiple dengue virus serotypes. Am. J. Trop. Med. Hyg. 1999, 61, 725–730. [Google Scholar] [CrossRef]
- Monath, T.P. Dengue: The risk to developed and developing countries. Proc. Natl. Acad. Sci. USA 1994, 91, 2395–2400. [Google Scholar] [CrossRef] [PubMed]
- Carrington, L.B.; Simmons, C.P. Human to mosquito transmission of dengue viruses. Front. Immunol. 2014, 5, 290. [Google Scholar] [CrossRef]
- Gubler, D.J. Dengue and dengue hemorrhagic fever. Semin. Pediatr. Infect. Dis. 1997, 8, 3–9. [Google Scholar] [CrossRef]
- Klowden, M.J.; Lea, A.O. Blood meal size as a factor affecting continued host-seeking by Aedes aegypti (L.)*. Am. J. Trop. Med. Hyg. 1978, 27, 827–831. [Google Scholar] [CrossRef]
- Devine, G.J.; Perea, E.Z.; Killeen, G.F.; Stancil, J.D.; Clark, S.J.; Morrison, A.C. Using adult mosquitoes to transfer insecticides to Aedes aegypti larval habitats. Proc. Natl. Acad. Sci. USA 2009, 106, 11530–11534. [Google Scholar] [CrossRef] [PubMed]
- Moyes, C.L.; Vontas, J.; Martins, A.J.; Ng, L.C.; Koou, S.Y.; Dusfour, I.; Raghavendra, K.; Pinto, J.; Corbel, V.; David, J.-P. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS Negl. Trop. Dis. 2017, 11, e0005625. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Ross, P.A.; Rašić, G. Wolbachia strains for disease control: Ecological and evolutionary considerations. Evol. Appl. 2015, 8, 751–768. [Google Scholar] [CrossRef] [PubMed]
- Molloy, J.C.; Sommer, U.; Viant, M.R.; Sinkins, S.P. Wolbachia modulates lipid metabolism in Aedes albopictus mosquito cells. Appl. Environ. Microbiol. 2016, 82, 3109–3120. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.; Johnson, P.H.; Moreira, L.A.; Iturbe-Ormaetxe, I.; Frentiu, F.D.; McMeniman, C.J.; Leong, Y.S.; Dong, Y.; Axford, J.; Kriesner, P.; et al. The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature 2011, 476, 450–453. [Google Scholar] [CrossRef] [PubMed]
- Bian, G.; Xu, Y.; Lu, P.; Xie, Y.; Xi, Z. The endosymbiotic bacterium Wolbachia induces resistance to dengue virus in Aedes aegypti. PLoS Pathog. 2010, 6, e1000833. [Google Scholar] [CrossRef] [PubMed]
- Geoghegan, V.; Stainton, K.; Rainey, S.M.; Ant, T.H.; Dowle, A.A.; Larson, T.; Hester, S.; Charles, P.D.; Thomas, B.; Sinkins, S.P. Perturbed cholesterol and vesicular trafficking associated with dengue blocking in Wolbachia-infected Aedes aegypti cells. Nat. Commun. 2017, 8, 526. [Google Scholar] [CrossRef] [PubMed]
- McMeniman, C.J.; Lane, R.V.; Cass, B.N.; Fong, A.W.C.; Sidhu, M.; Wang, Y.-F.; O’Neill, S.L. Stable introduction of a life-shortening Wolbachia infection into the mosquito Aedes aegypti. Science 2009, 323, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Khoo, C.C.; Dobson, S.L. Wolbachia establishment and invasion in an Aedes aegypti laboratory population. Science 2005, 310, 326–328. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Montgomery, B.L.; Popovici, J.; Iturbe-Ormaetxe, I.; Johnson, P.H.; Muzzi, F.; Greenfield, M.; Durkan, M.; Leong, Y.S.; Dong, Y.; et al. Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature 2011, 476, 454–457. [Google Scholar] [CrossRef]
- Richardson, K.M.; Hoffmann, A.A.; Johnson, P.; Ritchie, S.R.; Kearney, M.R. A replicated comparison of breeding-container suitability for the dengue vector Aedes aegypti in tropical and temperate Australia. Austral Ecol. 2013, 38, 219–229. [Google Scholar] [CrossRef]
- Ulrich, J.N.; Beier, J.C.; Devine, G.J.; Hugo, L.E. Heat sensitivity of wMel Wolbachia during Aedes aegypti development. PLoS Negl. Trop. Dis. 2016, 10, e0004873. [Google Scholar] [CrossRef]
- Ross, P.A.; Wiwatanaratanabutr, I.; Axford, J.K.; White, V.L.; Endersby-Harshman, N.M.; Hoffmann, A.A. Wolbachia infections in Aedes aegypti differ markedly in their response to cyclical heat stress. PLoS Pathog. 2017, 13, e1006006. [Google Scholar] [CrossRef]
- Lu, P.; Bian, G.; Pan, X.; Xi, Z. Wolbachia induces density-dependent inhibition to dengue virus in mosquito cells. PLoS Negl. Trop. Dis. 2012, 6, e1754. [Google Scholar] [CrossRef] [PubMed]
- Osborne, S.E.; Iturbe-Ormaetxe, I.; Brownlie, J.C.; O’Neill, S.L.; Johnson, K.N. Antiviral protection and the importance of Wolbachia density and tissue tropism in Drosophila simulans. Appl. Environ. Microbiol. 2012, 78, 6922–6929. [Google Scholar] [CrossRef] [PubMed]
- Axford, J.K.; Ross, P.A.; Yeap, H.L.; Callahan, A.G.; Hoffmann, A.A. Fitness of wAlbB Wolbachia infection in Aedes aegypti: Parameter estimates in an outcrossed background and potential for population invasion. Am. J. Trop. Med. Hyg. 2016, 94, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.A.; Axford, J.K.; Richardson, K.M.; Endersby-Harshman, N.M.; Hoffmann, A.A. Maintaining Aedes aegypti mosquitoes infected with Wolbachia. J. Vis. Exp. 2017, 126, e56124. [Google Scholar]
- Hoffmann, A.A.; Harshman, L.G. Desiccation and starvation resistance in Drosophila: Patterns of variation at the species, population and intrapopulation levels. Heredity 1999, 83, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Yeap, H.L.; Endersby, N.M.; Johnson, P.H.; Ritchie, S.A.; Hoffmann, A.A. Body size and wing shape measurements as quality indicators of Aedes aegypti mosquitoes destined for field release. Am. J. Trop. Med. Hyg. 2013, 89, 78–92. [Google Scholar] [CrossRef]
- Lee, S.F.; White, V.L.; Weeks, A.R.; Hoffmann, A.A.; Endersby, N.M. High-throughput PCR assays to monitor Wolbachia infection in the dengue mosquito (Aedes aegypti) and Drosophila simulans. Appl. Environ. Microbiol. 2012, 78, 4740–4743. [Google Scholar] [CrossRef]
- Ross, P.A. Environmental Impacts on the Use of Modified Mosquitoes for Arbovirus Control. Ph.D. Thesis, The University of Melbourne, Melbourne, Australia, 2018. [Google Scholar]
- Ross, P.A.; Hoffmann, A.A. Continued susceptibility of the wMel Wolbachia infection in Aedes aegypti to heat stress following field deployment and selection. Insects 2018, 9, 78. [Google Scholar] [CrossRef]
- Ant, T.H.; Herd, C.S.; Geoghegan, V.; Hoffmann, A.A.; Sinkins, S.P. The Wolbachia strain wAu provides highly efficient virus transmission blocking in Aedes aegypti. PLoS Pathog. 2018, 14, e1006815. [Google Scholar] [CrossRef]
- Li, Y.-Y.; Floate, K.; Fields, P.; Pang, B.-P. Review of treatment methods to remove Wolbachia bacteria from arthropods. Symbiosis 2014, 62, 1–15. [Google Scholar] [CrossRef]
- Jia, F.-X.; Yang, M.-S.; Yang, W.-J.; Wang, J.-J. Influence of continuous high temperature conditions on Wolbachia infection frequency and the fitness of Liposcelis tricolor (Psocoptera: Liposcelididae). Environ. Entomol. 2009, 38, 1365–1372. [Google Scholar] [CrossRef]
- Van Opijnen, T.; Breeuwer, J. High temperatures eliminate Wolbachia, a cytoplasmic incompatibility inducing endosymbiont, from the two-spotted spider mite. Exp. Appl. Acarol. 1999, 23, 871–881. [Google Scholar] [CrossRef]
- Mouton, L.; Henri, H.; Bouletreau, M.; Vavre, F. Effect of temperature on Wolbachia density and impact on cytoplasmic incompatibility. Parasitology 2006, 132, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Chrostek, E.; Marialva, M.S.; Esteves, S.S.; Weinert, L.A.; Martinez, J.; Jiggins, F.M.; Teixeira, L. Wolbachia variants induce differential protection to viruses in Drosophila melanogaster: A phenotypic and phylogenomic analysis. PLoS Genet. 2013, 9, e1003896. [Google Scholar] [CrossRef]
- Martinez, J.; Tolosana, I.; Ok, S.; Smith, S.; Snoeck, K.; Day, J.P.; Jiggins, F.M. Symbiont strain is the main determinant of variation in Wolbachia-mediated protection against viruses across Drosophila species. Mol. Ecol. 2017, 26, 4072–4084. [Google Scholar] [CrossRef] [PubMed]
- Veneti, Z.; Clark, M.E.; Zabalou, S.; Karr, T.L.; Savakis, C.; Bourtzis, K. Cytoplasmic incompatibility and sperm cyst infection in different Drosophila-Wolbachia associations. Genetics 2003, 164, 545–552. [Google Scholar] [PubMed]
- Bordenstein, S.R.; Marshall, M.L.; Fry, A.J.; Kim, U.; Wernegreen, J.J. The tripartite associations between bacteriophage, Wolbachia, and arthropods. PLoS Pathog. 2006, 2, e43. [Google Scholar]
- Nguyen, T.H.; Le Nguyen, H.; Nguyen, T.Y.; Vu, S.N.; Tran, N.D.; Le, T.; Vien, Q.M.; Bui, T.; Le, H.T.; Kutcher, S. Field evaluation of the establishment potential of wMelpop Wolbachia in Australia and Vietnam for dengue control. Parasit Vectors 2015, 8, 563. [Google Scholar] [CrossRef]
- Schmidt, T.L.; Barton, N.H.; Rašić, G.; Turley, A.P.; Montgomery, B.L.; Iturbe-Ormaetxe, I.; Cook, P.E.; Ryan, P.A.; Ritchie, S.A.; Hoffmann, A.A. Local introduction and heterogeneous spatial spread of dengue-suppressing Wolbachia through an urban population of Aedes aegypti. PLoS Biol. 2017, 15, e2001894. [Google Scholar] [CrossRef]
- O’Neill, S.L.; Ryan, P.A.; Turley, A.P.; Wilson, G.; Retzki, K.; Iturbe-Ormaetxe, I.; Dong, Y.; Kenny, N.; Paton, C.J.; Ritchie, S.A. Scaled deployment of Wolbachia to protect the community from dengue and other Aedes transmitted arboviruses. Gates Open Res. 2018, 2, 36. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Iturbe-Ormaetxe, I.; Callahan, A.G.; Phillips, B.L.; Billington, K.; Axford, J.K.; Montgomery, B.; Turley, A.P.; O’Neill, S.L. Stability of the wMel Wolbachia infection following invasion into Aedes aegypti populations. PLoS Negl. Trop. Dis. 2014, 8, e3115. [Google Scholar] [CrossRef]
- Joubert, D.A.; Walker, T.; Carrington, L.B.; De Bruyne, J.T.; Kien, D.H.T.; Hoang, N.L.T.; Chau, N.V.V.; Iturbe-Ormaetxe, I.; Simmons, C.P.; O’Neill, S.L. Establishment of a Wolbachia superinfection in Aedes aegypti mosquitoes as a potential approach for future resistance management. PLoS Pathog. 2016, 12, e1005434. [Google Scholar] [CrossRef]
- Yeap, H.L.; Mee, P.; Walker, T.; Weeks, A.R.; O’Neill, S.L.; Johnson, P.; Ritchie, S.A.; Richardson, K.M.; Doig, C.; Endersby, N.M. Dynamics of the ‘popcorn’ Wolbachia infection in outbred Aedes aegypti informs prospects for mosquito vector control. Genetics 2011, 187, 583–595. [Google Scholar] [CrossRef] [PubMed]
- McMeniman, C.J.; O’Neill, S.L. A virulent Wolbachia infection decreases the viability of the dengue vector Aedes aegypti during periods of embryonic quiescence. PLoS Negl. Trop. Dis. 2010, 4, e748. [Google Scholar] [CrossRef]
- Harcombe, W.; Hoffmann, A. Wolbachia effects in Drosophila melanogaster: In search of fitness benefits. J. Invertebr. Pathol. 2004, 87, 45–50. [Google Scholar] [CrossRef]
- Hamm, C.A.; Begun, D.J.; Vo, A.; Smith, C.C.; Saelao, P.; Shaver, A.O.; Jaenike, J.; Turelli, M. Wolbachia do not live by reproductive manipulation alone: Infection polymorphism in Drosophila suzukii and D. subpulchrella. Mol. Ecol. 2014, 23, 4871–4885. [Google Scholar] [CrossRef]
- Ross, P.A.; Endersby, N.M.; Hoffmann, A.A. Costs of three Wolbachia infections on the survival of Aedes aegypti larvae under starvation conditions. PLoS Negl. Trop. Dis. 2016, 10, e0004320. [Google Scholar] [CrossRef]
- Johnson, B.J.; Mitchell, S.N.; Paton, C.J.; Stevenson, J.; Staunton, K.M.; Snoad, N.; Beebe, N.; White, B.J.; Ritchie, S.A. Use of rhodamine B to mark the body and seminal fluid of male Aedes aegypti for mark-release-recapture experiments and estimating efficacy of sterile male releases. PLoS Negl. Trop. Dis. 2017, 11, e0005902. [Google Scholar] [CrossRef]
- Evans, O.; Caragata, E.P.; McMeniman, C.J.; Woolfit, M.; Green, D.C.; Williams, C.R.; Franklin, C.E.; O’Neill, S.L.; McGraw, E.A. Increased locomotor activity and metabolism of Aedes aegypti infected with a life-shortening strain of Wolbachia pipientis. J. Exp. Biol. 2009, 212, 1436–1441. [Google Scholar] [CrossRef] [PubMed]
- Caragata, E.P.; Rancès, E.; O’Neill, S.L.; McGraw, E.A. Competition for amino acids between Wolbachia and the mosquito host, Aedes aegypti. Microb. Ecol. 2014, 67, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Edman, J.D.; Strickman, D.; Kittayapong, P.; Scott, T.W. Female Aedes aegypti (Diptera: Culicidae) in Thailand rarely feed on sugar. J. Med. Entomol. 1992, 29, 1035–1038. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infection Type | Generation | |
|---|---|---|
| 1 | 2 | |
| Uninfected | 26 °C | 26 °C |
| 26 °C | 26–37 °C | |
| 26–37 °C | 26 °C | |
| 26–37 °C | 26–37 °C | |
| wMel | 26 °C | 26 °C |
| 26 °C | 26–37 °C | |
| 26–37 °C | 26 °C | |
| 26–37 °C | 26–37 °C | |
| wAlbB | 26 °C | 26 °C |
| 26 °C | 26–37 °C | |
| 26–37 °C | 26 °C | |
| 26–37 °C | 26–37 °C | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foo, I.J.-H.; Hoffmann, A.A.; Ross, P.A. Cross-Generational Effects of Heat Stress on Fitness and Wolbachia Density in Aedes aegypti Mosquitoes. Trop. Med. Infect. Dis. 2019, 4, 13. https://doi.org/10.3390/tropicalmed4010013
Foo IJ-H, Hoffmann AA, Ross PA. Cross-Generational Effects of Heat Stress on Fitness and Wolbachia Density in Aedes aegypti Mosquitoes. Tropical Medicine and Infectious Disease. 2019; 4(1):13. https://doi.org/10.3390/tropicalmed4010013
Chicago/Turabian StyleFoo, Isabelle Jia-Hui, Ary A. Hoffmann, and Perran A. Ross. 2019. "Cross-Generational Effects of Heat Stress on Fitness and Wolbachia Density in Aedes aegypti Mosquitoes" Tropical Medicine and Infectious Disease 4, no. 1: 13. https://doi.org/10.3390/tropicalmed4010013
APA StyleFoo, I. J.-H., Hoffmann, A. A., & Ross, P. A. (2019). Cross-Generational Effects of Heat Stress on Fitness and Wolbachia Density in Aedes aegypti Mosquitoes. Tropical Medicine and Infectious Disease, 4(1), 13. https://doi.org/10.3390/tropicalmed4010013