Rabies Virus Antibodies from Oral Vaccination as a Correlate of Protection against Lethal Infection in Wildlife

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Literature Review
2.2. Animal Studies
2.3. Animals
2.4. Vaccine
2.5. Serum Samples
2.6. Serology
2.7. Statistical Analysis
3. Results
3.1. Literature Review
3.1.1. Serological Methods and Study Design Summary
3.1.2. Serological Results Summary
3.2. Animal Studies
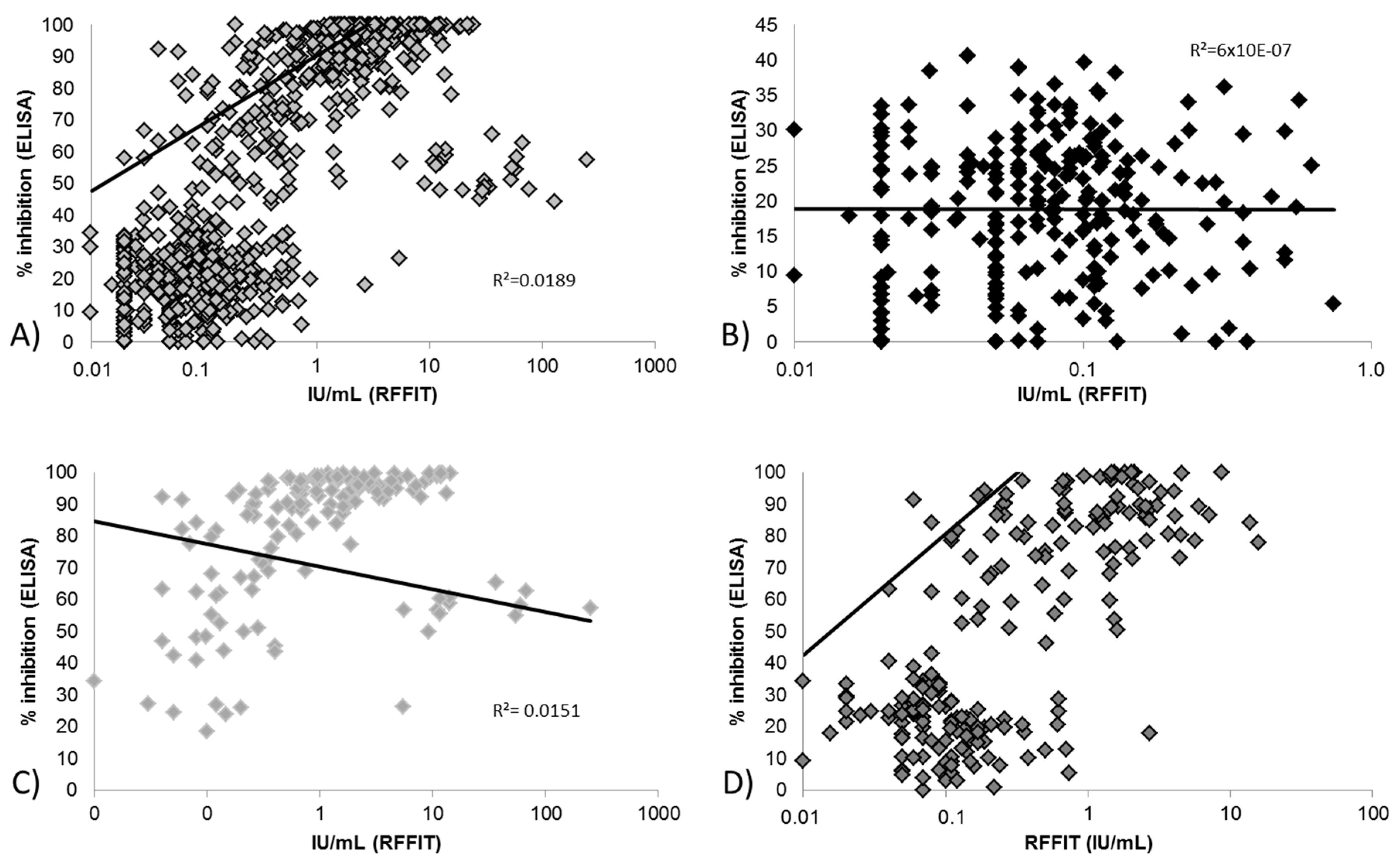
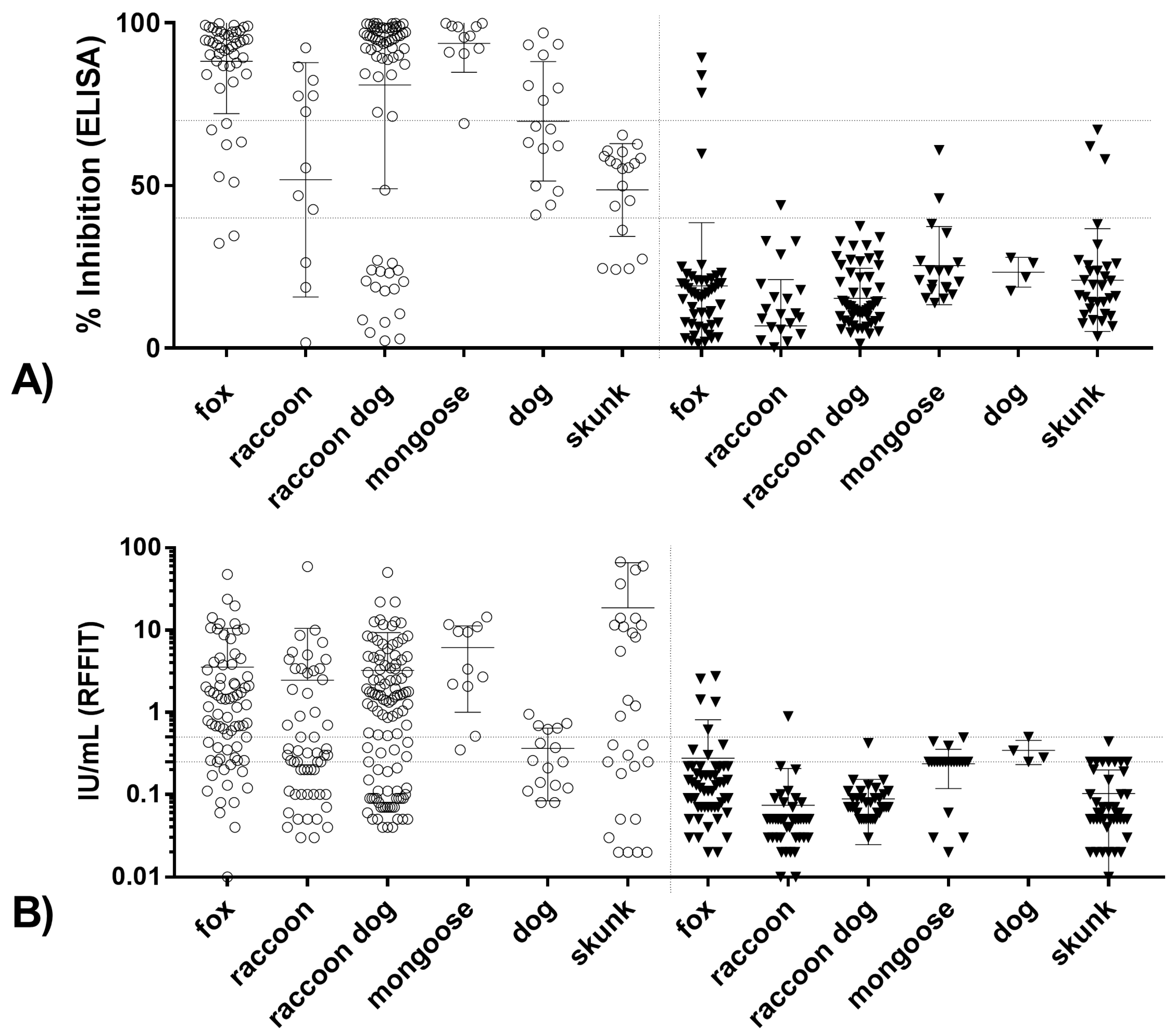
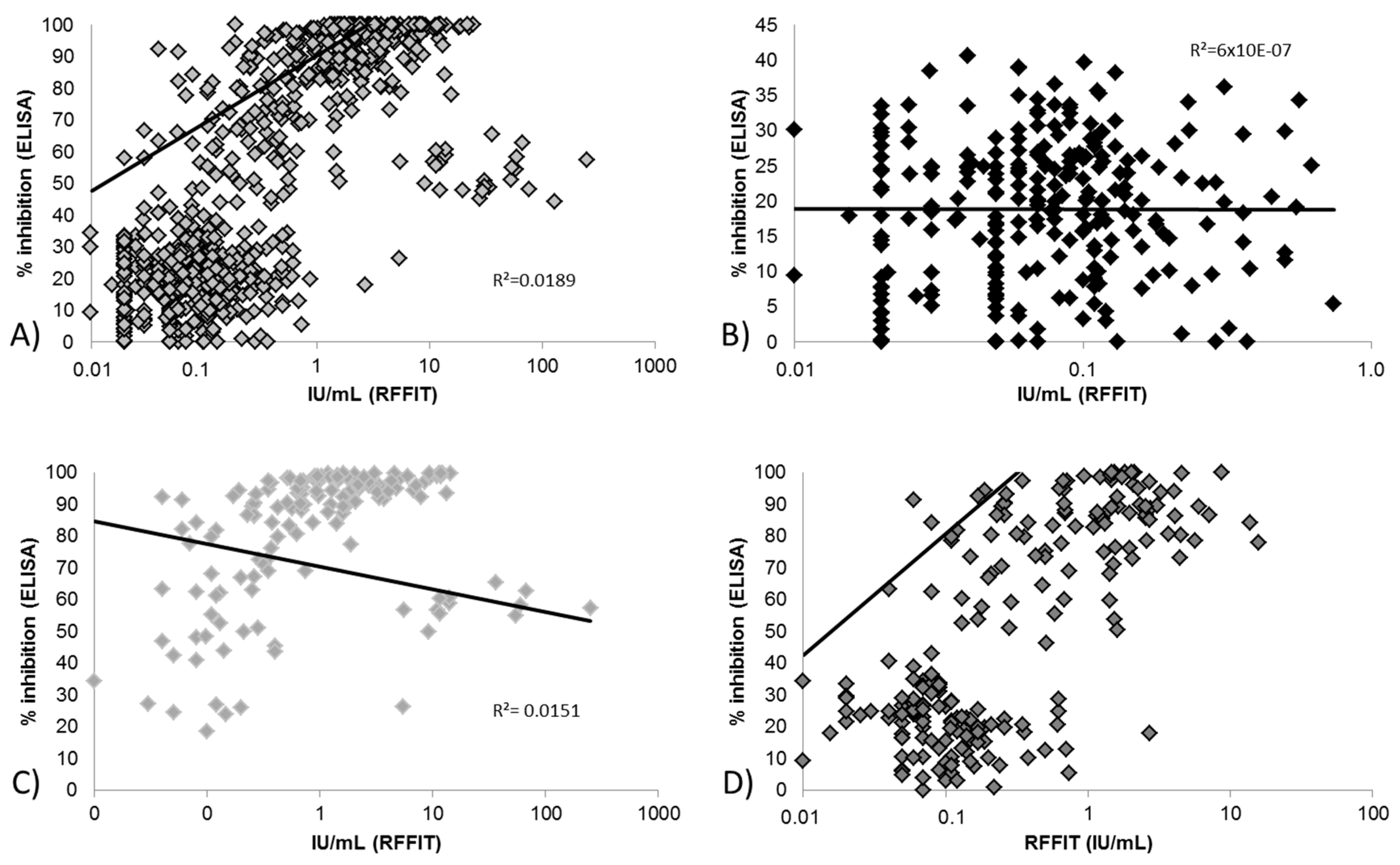
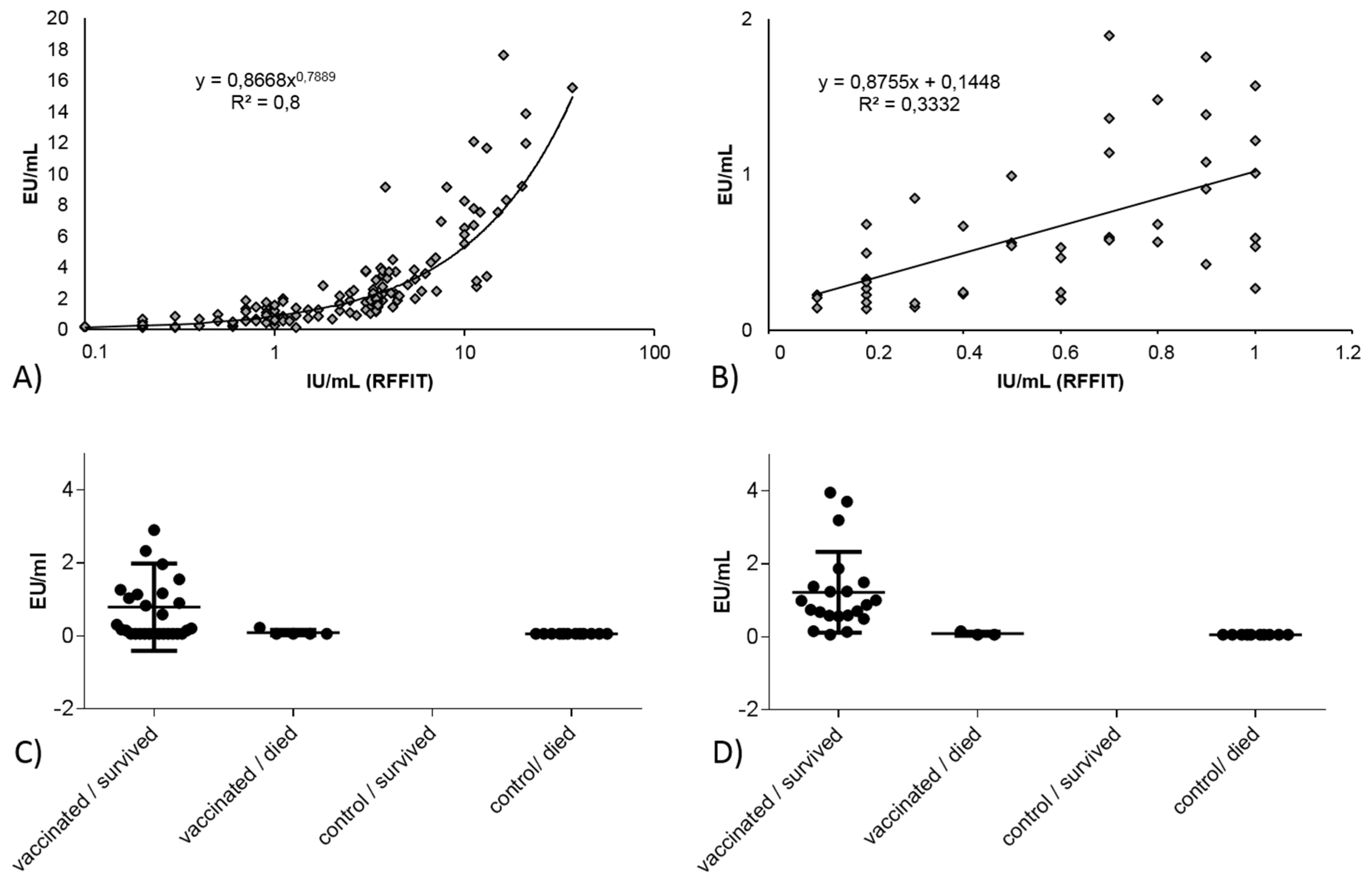
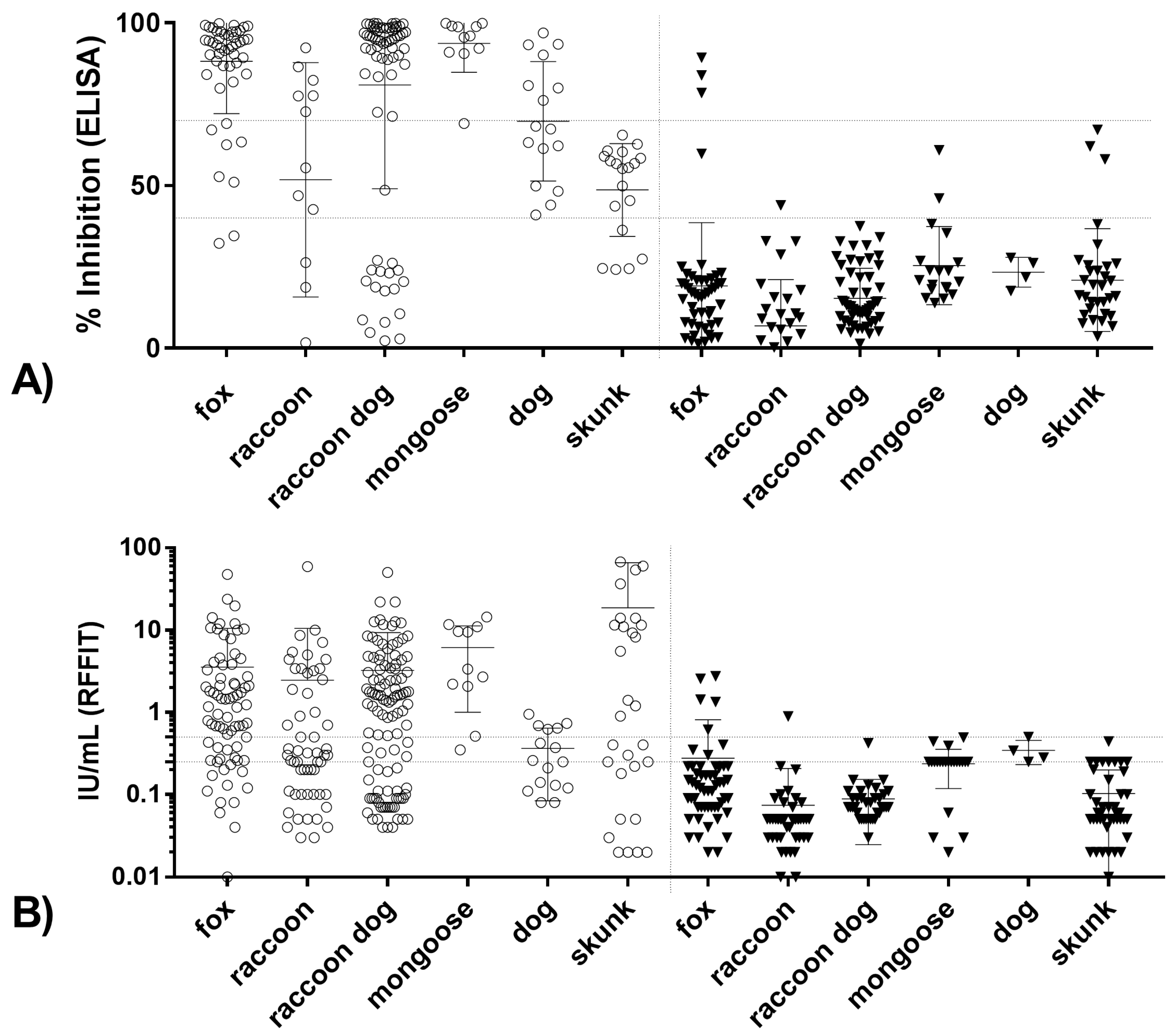
3.2.1. Correlation between Blocking ELISA and RFFIT Values and Test Agreement
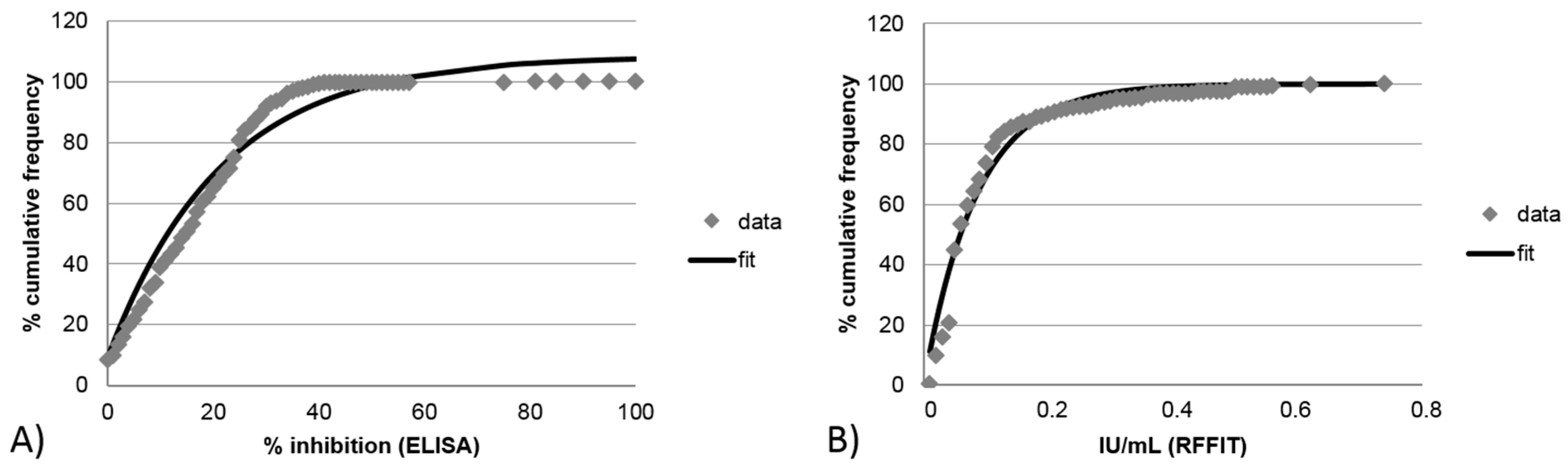
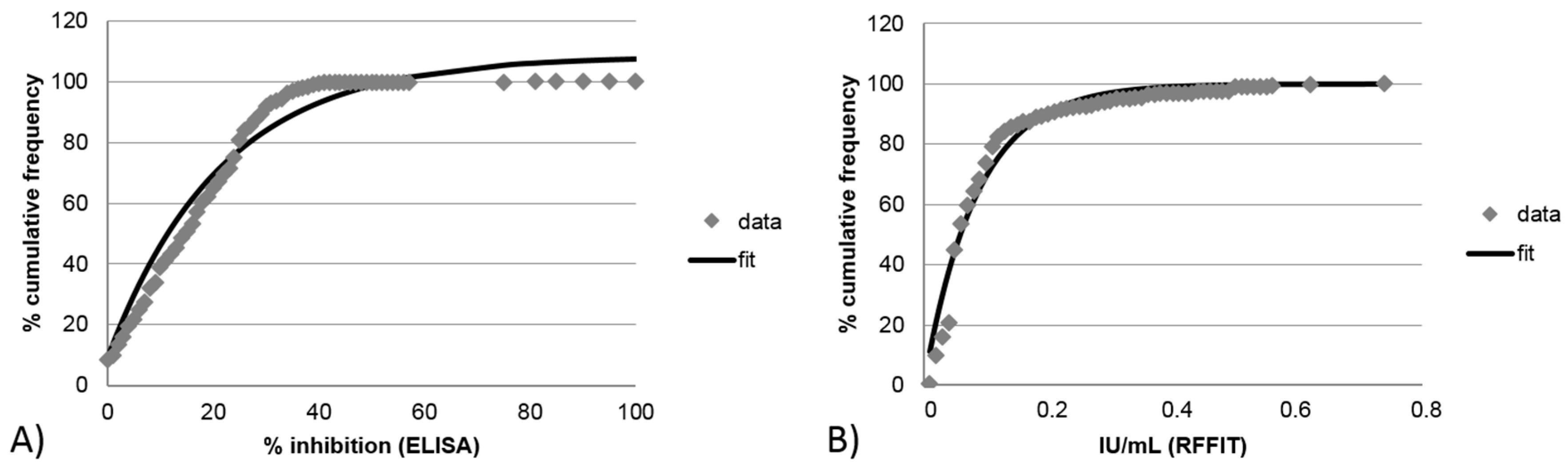
3.2.2. Limit of Detection for Blocking ELISA and RFFIT
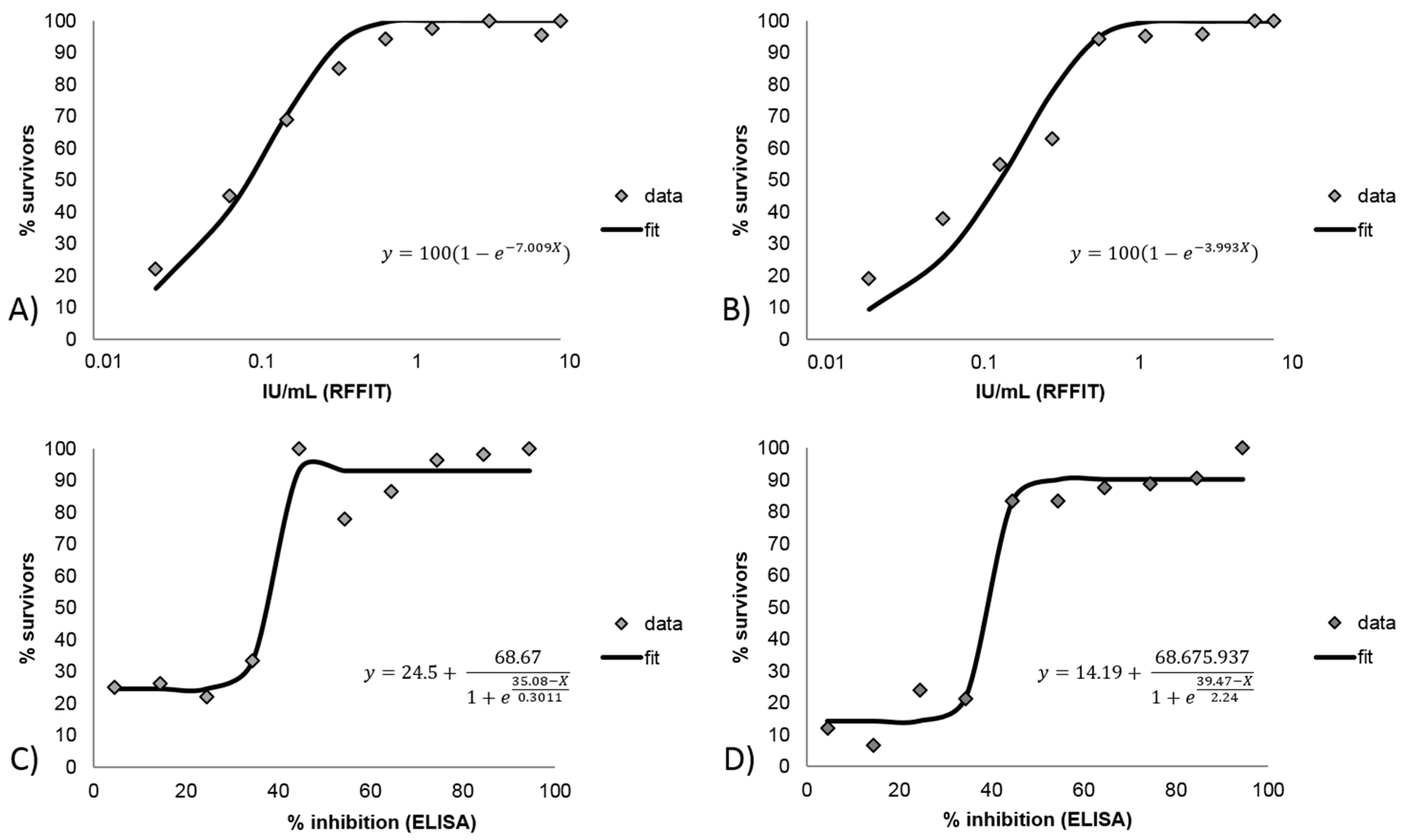
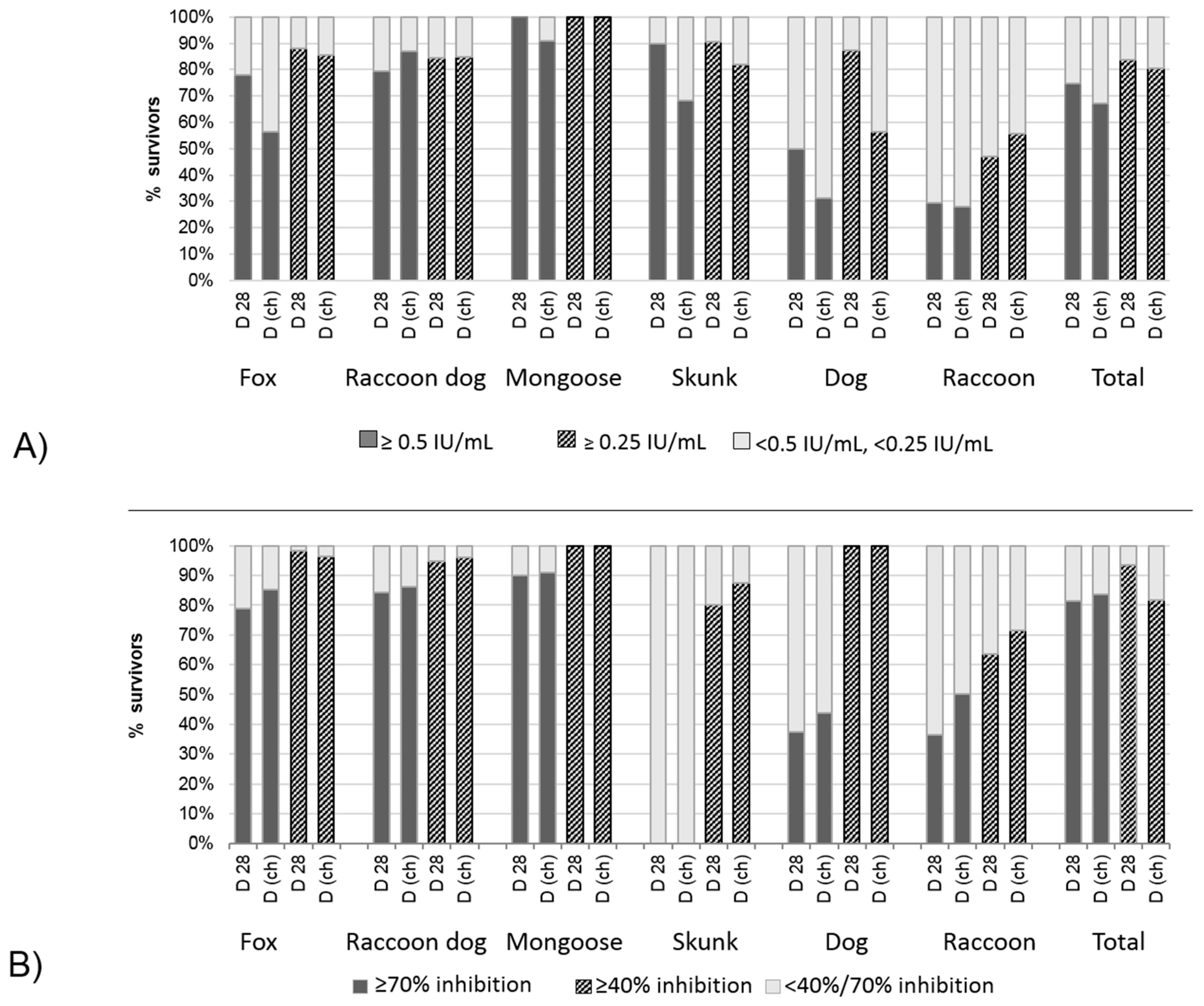
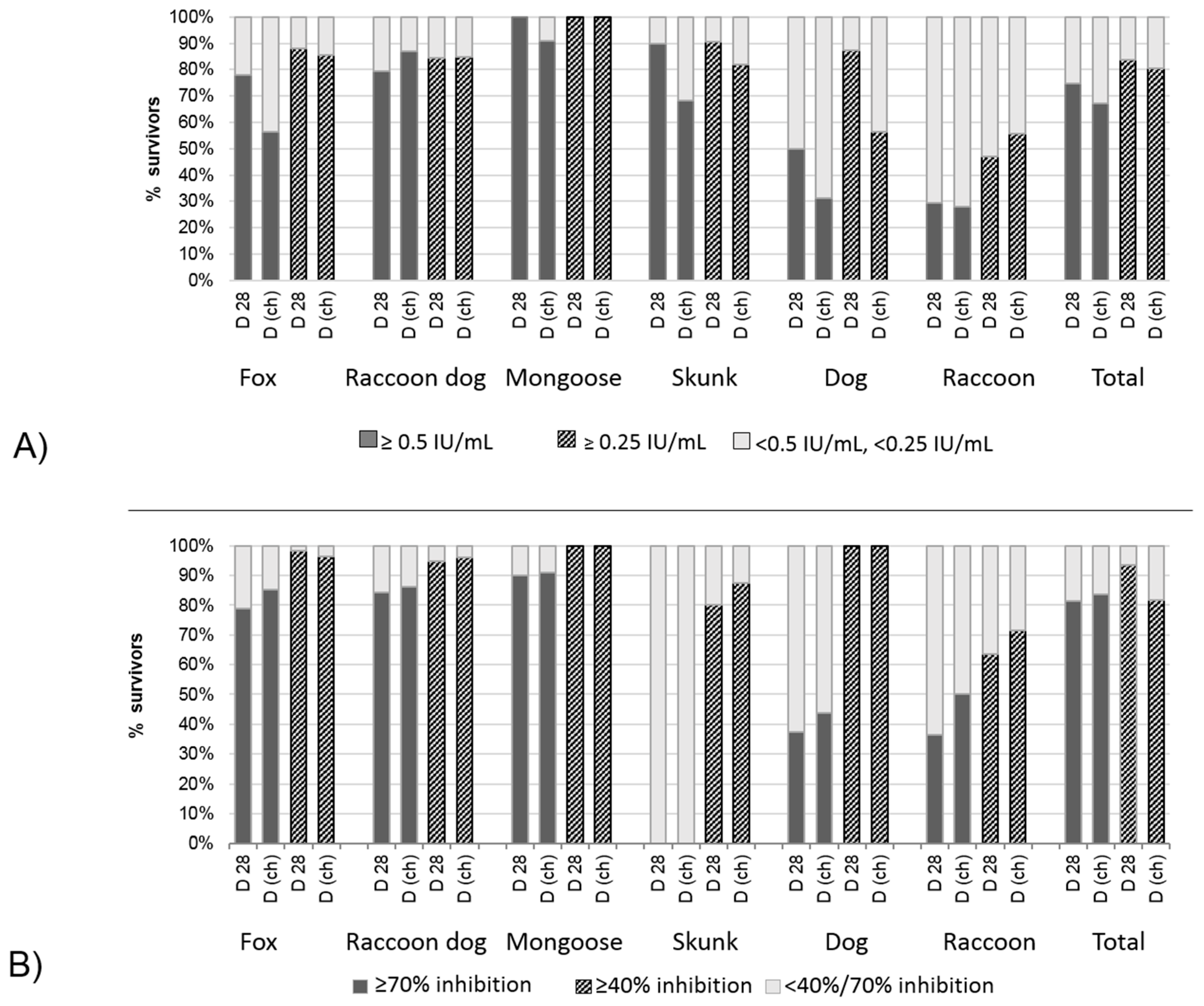
3.2.3. Correlation and Agreement of Test Results Obtained in Blocking ELISA and RFFIT with Survival/Death
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dietzschold, B.; Ruppecht, C.E.; Fu, Z.F.; Koprowski, H. Rhabdoviruses. In Field’s Virology, 3rd ed.; Fields, B.N, Knipe, D.M., Howley, P.M., Eds.; Raven Press: Philadelphia, PA, USA, 1996; pp. 1137–1159. [Google Scholar]
- Dietzschold, B.; Li, J.; Faber, M.; Schnell, M. Concepts in the pathogenesis of rabies. Future Med. 2008, 3, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Steck, F.; Wandeler, A.; Bichsel, P.; Capt, S.; Hafliger, U.; Schneider, L. Oral immunization of foxes against rabies. Laboratory and field studies. Comp. Immunol. Microbiol. Infect. Dis. 1982, 5, 165–171. [Google Scholar] [CrossRef]
- Wandeler, A.I.; Capt, S.; Kappeler, A.; Hauser, R. Oral immunization of wildlife against rabies: Concept and first field experiments. Rev. Infect. Dis. 1988, 10 (Suppl. 4), S649–S653. [Google Scholar] [CrossRef] [PubMed]
- Fooks, A.; Horton, D.; Muller, T.; Freuling, C.; Rupprecht, C. Rabies. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals (Mammals, Birds, Bees), 7th ed.; Office International Des Epizooties: Paris, France, 2012; pp. 263–282. [Google Scholar]
- Bunn, T.O.; Ridpath, H.D. The Relationship Between Rabies Antibody Titers in Dogs and Cats and Protection from Challenge; U.S. Department of Health, Education and Welfare, Public Health: Washington, DC, USA, 1984; pp. 43–45.
- Aubert, M.F. Practical significance of rabies antibodies in cats and dogs. Rev. Sci. Tech. 1992, 11, 735–760. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.M.; Hanlon, C.A. Rabies-specific antibodies: Measuring surrogates of protection against a fatal disease. PLoS Negl. Trop. Dis. 2010, 4, e595. [Google Scholar] [CrossRef] [PubMed]
- Sikes, R.K.; Peacock, G.V.; Acha, P.; Arko, R.J.; Dierks, R. Rabies vaccines: Duration-of-immunity study in dogs. J. Am. Vet. Med. Assoc. 1971, 159, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.M.; Wilkerson, M.J.; Davis, R.D.; Wyatt, C.R.; Briggs, D.J. Detection of cellular immunity to rabies antigens in human vaccinees. J. Clin. Immunol. 2006, 26, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Haralambieva, I.H.; Ovsyannikova, I.G.; Pankratz, V.S.; Kennedy, R.B.; Jacobson, R.M.; Poland, G.A. The genetic basis for interindividual immune response variation to measles vaccine: New understanding and new vaccine approaches. Expert. Rev. Vaccines 2013, 12, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Ovsyannikova, I.G.; Schaid, D.J.; Larrabee, B.R.; Haralambieva, I.H.; Kennedy, R.B.; Poland, G.A. A large population-based association study between HLA and KIR genotypes and measles vaccine antibody responses. PLoS ONE 2017, 12, e0171261. [Google Scholar] [CrossRef] [PubMed]
- Schaid, D.J.; Haralambieva, I.H.; Larrabee, B.R.; Ovsyannikova, I.G.; Kennedy, R.B.; Poland, G.A. Heritability of vaccine-induced measles neutralizing antibody titers. Vaccine 2017, 35, 1390–1394. [Google Scholar] [CrossRef] [PubMed]
- Kuwert, E.; Barsenbach, J.; Werner, J.; Mardus, I.; Scheiermann, N.; Grosse-Wilde, H. Early/high and late/low responders among HDCS vaccinees. In Cell Culture Rabies Vaccines and their Protective Effect in Man; Wiktor, T.J., Koprowski, H., Eds.; International Green Cross: Geneva, Switzerland, 1981; pp. 160–168. [Google Scholar]
- Feyssaguet, M.; Dacheux, L.; Audry, L.; Compoint, A.; Morize, J.L.; Blanchard, I.; Bourhy, H. Multicenter comparative study of a new ELISA, Platelia Rabies II, for the detection and titration of anti-rabies glycoprotein antibodies and comparison with the rapid fluorescent focus inhibition test (RFFIT) on human samples from vaccinated and non-vaccinated people. Vaccine 2007, 25, 2244–2251. [Google Scholar] [PubMed]
- Welch, R.J.; Anderson, B.L.; Litwin, C.M. An evaluation of two commercially available ELISAs and one in-house reference laboratory ELISA for the determination of human anti-rabies virus antibodies. J. Med. Microbiol. 2009, 58, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Cliquet, F.; McElhinney, L.M.; Servat, A.; Boucher, J.M.; Lowings, J.P.; Goddard, T.; Mansfield, K.L.; Fooks, A.R. Development of a qualitative indirect ELISA for the measurement of rabies virus-specific antibodies from vaccinated dogs and cats. J. Virol. Methods 2004, 117, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.M.; Pralle, S.; Engelman, L.; Hartschuh, H.; Smith, M. Rabies vaccine response measurement is assay dependent. Biologicals 2016, 44, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Artois, M.; Masson, E.; Barrat, J.; Aubert, M.F. Efficacy of three oral rabies vaccine-baits in the red fox: A comparison. Vet. Microbiol. 1993, 38, 167–172. [Google Scholar] [CrossRef]
- Black, J.G.; Lawson, K.F. Further studies of sylvatic rabies in the fox (Vulpes vulpes). Vaccination by the oral route. Can. Vet. J. 1973, 14, 206–211. [Google Scholar] [PubMed]
- Black, J.G.; Lawson, K.F. The safety and efficacy of immunizing foxes (Vulpes vulpes) using bait containing attenuated rabies virus vaccine. Can. J. Comp. Med. 1980, 44, 169–176. [Google Scholar] [PubMed]
- Blancou, J.; Kieny, M.P.; Lathe, R.; Lecocq, J.P.; Pastoret, P.P.; Soulebot, J.P.; Desmettre, P. Oral vaccination of the fox against rabies using a live recombinant vaccinia virus. Nature 1986, 322, 373–375. [Google Scholar] [CrossRef] [PubMed]
- Brochier, B.M.; Languet, B.; Blancou, J.; Kieny, M.P.; Lecocq, J.P.; Costy, F.; Desmettre, P.; Pastoret, P.P. Use of recombinant vaccinia-rabies virus for oral vaccination of fox cubs (Vulpes vulpes, l) against rabies. Vet. Microbiol. 1988, 18, 103–108. [Google Scholar] [CrossRef]
- Brown, L.J.; Rosatte, R.C.; Fehlner-Gardiner, C.; Taylor, J.S.; Davies, J.C.; Donovan, D. Immune response and protection in raccoons (Procyon lotor) following consumption of baits containing ONRAB®, a human adenovirus rabies glycoprotein recombinant vaccine. J. Wildl. Dis. 2012, 48, 1010–1020. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.J.; Rosatte, R.C.; Fehlner-Gardiner, C.; Ellison, J.A.; Jackson, F.R.; Bachmann, P.; Taylor, J.S.; Franka, R.; Donovan, D. Oral vaccination and protection of striped skunks (Mephitis mephitis) against rabies using ONRAB®. Vaccine 2014, 32, 3675–3679. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.J.; Rosatte, R.C.; Fehlner-Gardiner, C.; Bachmann, P.; Ellison, J.A.; Jackson, F.R.; Taylor, J.S.; Davies, C.; Donovan, D. Oral vaccination and protection of red foxes (Vulpes vulpes) against rabies using ONRAB, an adenovirus-rabies recombinant vaccine. Vaccine 2014, 32, 984–989. [Google Scholar] [CrossRef] [PubMed]
- Follmann, E.; Ritter, D.; Swor, R.; Dunbar, M.; Hueffer, K. Preliminary evaluation of Raboral V-RG® oral rabies vaccine in arctic foxes (Vulpes lagopus). J. Wildl. Dis. 2011, 47, 1032–1035. [Google Scholar] [CrossRef] [PubMed]
- Follmann, E.H.; Ritter, D.G.; Donald, W.H. Oral vaccination of captive arctic foxes with lyophilized SAG2 rabies vaccine. J. Wildl. Dis. 2004, 40, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Follmann, E.H.; Ritter, D.G.; Baer, G.M. Oral rabies vaccination of arctic foxes (Alopex lagopus) with an attenuated vaccine. Vaccine 1992, 10, 305–308. [Google Scholar] [CrossRef]
- Follmann, E.H.; Ritter, D.G.; Baer, G.M. Immunization of arctic foxes (Alopex lagopus) with oral rabies vaccine. J. Wildl. Dis. 1988, 24, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Lawson, K.F.; Chiu, H.; Crosgrey, S.J.; Matson, M.; Casey, G.A.; Campbell, J.B. Duration of immunity in foxes vaccinated orally with era vaccine in a bait. Can. J. Vet. Res. 1997, 61, 39–42. [Google Scholar] [PubMed]
- Le Blois, H.; Tuffereau, C.; Blancou, J.; Artois, M.; Aubert, A.; Flamand, A. Oral immunization of foxes with avirulent rabies virus mutants. Vet. Microbiol. 1990, 23, 259–266. [Google Scholar]
- Neubert, A.; Schuster, P.; Müller, T.; Vos, A.; Pommerening, E. Immunogenicity and efficacy of the oral rabies vaccine SAD B19 in foxes. J. Vet. Med. B Infect. Dis. Vet. Public Health 2001, 48, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Tolson, N.D.; Charlton, K.M.; Lawson, K.F.; Campbell, J.B.; Stewart, R.B. Studies of ERA/BHK-21 rabies vaccine in skunks and mice. Can. J. Vet. Res. 1988, 52, 58–62. [Google Scholar] [PubMed]
- Tolson, N.D.; Charlton, K.M.; Casey, G.A.; Knowles, M.K.; Rupprecht, C.E.; Lawson, K.F.; Campbell, J.B. Immunization of foxes against rabies with a vaccinia recombinant virus expressing the rabies glycoprotein. Arch. Virol. 1988, 102, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Tolson, N.D.; Charlton, K.M.; Stewart, R.B.; Campbell, J.B.; Wiktor, T.J. Immune response in skunks to a vaccinia virus recombinant expressing the rabies virus glycoprotein. Can. J. Vet. Res. 1987, 51, 363–366. [Google Scholar] [PubMed]
- Vos, A.; Conzelmann, K.K.; Finke, S.; Müller, T.; Teifke, J.; Fooks, A.R.; Neubert, A. Immunogenicity studies in carnivores using a rabies virus construct with a site-directed deletion in the phosphoprotein. Adv. Prev. Med. 2011, 2011, 898171. [Google Scholar] [CrossRef] [PubMed]
- Cliquet, F.; Barrat, J.; Guiot, A.L.; Cael, N.; Boutrand, S.; Maki, J.; Schumacher, C.L. Efficacy and bait acceptance of vaccinia vectored rabies glycoprotein vaccine in captive foxes (Vulpes vulpes), raccoon dogs (Nyctereutes procyonoides) and dogs (Canis familiaris). Vaccine 2008, 26, 4627–4638. [Google Scholar] [CrossRef] [PubMed]
- Cliquet, F.; Gurbuxani, J.P.; Pradhan, H.K.; Pattnaik, B.; Patil, S.S.; Regnault, A.; Begouen, H.; Guiot, A.L.; Sood, R.; Mahl, P.; et al. The safety and efficacy of the oral rabies vaccine SAG2 in indian stray dogs. Vaccine 2007, 25, 3409–3418. [Google Scholar] [CrossRef] [PubMed]
- Charlton, K.M.; Artois, M.; Prevec, L.; Campbell, J.B.; Casey, G.A.; Wandeler, A.I.; Armstrong, J. Oral rabies vaccination of skunks and foxes with a recombinant human adenovirus vaccine. Arch. Virol. 1992, 123, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Fekadu, M.; Nesby, S.L.; Shaddock, J.H.; Schumacher, C.L.; Linhart, S.B.; Sanderlin, D.W. Immunogenicity, efficacy and safety of an oral rabies vaccine (SAG-2) in dogs. Vaccine 1996, 14, 465–468. [Google Scholar] [CrossRef]
- Fekadu, M.; Shaddock, J.H.; Sumner, J.W.; Sanderlin, D.W.; Knight, J.C.; Esposito, J.J.; Baer, G.M. Oral vaccination of skunks with raccoon poxvirus recombinants expressing the rabies glycoprotein or the nucleoprotein. J. Wildl. Dis. 1991, 27, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Grosenbaugh, D.A.; Maki, J.L.; Rupprecht, C.E.; Wall, D.K. Rabies challenge of captive striped skunks (Mephitis mephitis) following oral administration of a live vaccinia-vectored rabies vaccine. J. Wildl. Dis. 2007, 43, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Hanlon, C.A.; Niezgoda, M.; Morrill, P.; Rupprecht, C.E. Oral efficacy of an attenuated rabies virus vaccine in skunks and raccoons. J. Wildl. Dis. 2002, 38, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Henderson, H.; Jackson, F.; Bean, K.; Panasuk, B.; Niezgoda, M.; Slate, D.; Li, J.; Dietzschold, B.; Mattis, J.; Rupprecht, C.E. Oral immunization of raccoons and skunks with a canine adenovirus recombinant rabies vaccine. Vaccine 2009, 27, 7194–7197. [Google Scholar] [CrossRef] [PubMed]
- Blanton, J.D.; Meadows, A.; Murphy, S.M.; Manangan, J.; Hanlon, C.A.; Faber, M.L.; Dietzschold, B.; Rupprecht, C.E. Vaccination of small asian mongoose (Herpestes javanicus) against rabies. J. Wildl. Dis. 2006, 42, 663–666. [Google Scholar] [CrossRef] [PubMed]
- Blanton, J.D.; Self, J.; Niezgoda, M.; Faber, M.L.; Dietzschold, B.; Rupprecht, C. Oral vaccination of raccoons (Procyon lotor) with genetically modified rabies virus vaccines. Vaccine 2007, 25, 7296–7300. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, C.E.; Hanlon, C.A.; Blanton, J.; Manangan, J.; Morrill, P.; Murphy, S.; Niezgoda, M.; Orciari, L.A.; Schumacher, C.L.; Dietzschold, B. Oral vaccination of dogs with recombinant rabies virus vaccines. Virus Res. 2005, 111, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.J.; Rosatte, R.C.; Fehlner-Gardiner, C.; Knowles, M.K.; Bachmann, P.; Davies, J.C.; Wandeler, A.; Sobey, K.; Donovan, D. Immunogenicity and efficacy of two rabies vaccines in wild-caught, captive raccoons. J. Wildl. Dis. 2011, 47, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Esposito, J.J.; Knight, J.C.; Shaddock, J.H.; Novembre, F.J.; Baer, G.M. Successful oral rabies vaccination of raccoons with raccoon poxvirus recombinants expressing rabies virus glycoprotein. Virology 1988, 165, 313–316. [Google Scholar] [CrossRef]
- Orciari, L.A.; Niezgoda, M.; Hanlon, C.A.; Shaddock, J.H.; Sanderlin, D.W.; Yager, P.A.; Rupprecht, C.E. Rapid clearance of SAG-2 rabies virus from dogs after oral vaccination. Vaccine 2001, 19, 4511–4518. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, L.; Zhou, S.; Wang, Z.; Ruan, J.; Tang, L.; Jia, Z.; Cui, M.; Zhao, L.; Fu, Z.F. Recombinant rabies virus expressing dog GM-CSF is an efficacious oral rabies vaccine for dogs. Oncotarget 2015, 6, 38504–38516. [Google Scholar] [PubMed]
- Rupprecht, C.E.; Wiktor, T.J.; Johnston, D.H.; Hamir, A.N.; Dietzschold, B.; Wunner, W.H.; Glickman, L.T.; Koprowski, H. Oral immunization and protection of raccoons (Procyon lotor) with a vaccinia-rabies glycoprotein recombinant virus vaccine. Proc. Natl. Acad. Sci. USA 1986, 83, 7947–7950. [Google Scholar] [CrossRef] [PubMed]
- Faber, M.; Faber, M.L.; Papaneri, A.; Bette, M.; Weihe, E.; Dietzschold, B.; Schnell, M.J. A single amino acid change in rabies virus glycoprotein increases virus spread and enhances virus pathogenicity. J. Virol. 2005, 79, 14141–14148. [Google Scholar] [CrossRef] [PubMed]
- Faber, M.; Pulmanausahakul, R.; Hodawadekar, S.S.; Spitsin, S.; McGettigan, J.P.; Schnell, M.J.; Dietzschold, B. Overexpression of the rabies virus glycoprotein results in enhancement of apoptosis and antiviral immune response. J. Virol. 2002, 76, 3374–3381. [Google Scholar] [CrossRef] [PubMed]
- Faber, M.; Faber, M.L.; Li, J.; Preuss, M.A.; Schnell, M.J.; Dietzschold, B. Dominance of a nonpathogenic glycoprotein gene over a pathogenic glycoprotein gene in rabies virus. J. Virol. 2007, 81, 7041–7047. [Google Scholar] [CrossRef] [PubMed]
- Rieder, M.; Brzozka, K.; Pfaller, C.K.; Cox, J.H.; Stitz, L.; Conzelmann, K.K. Genetic dissection of interferon-antagonistic functions of rabies virus phosphoprotein: Inhibition of interferon regulatory factor 3 activation is important for pathogenicity. J. Virol. 2011, 85, 842–852. [Google Scholar] [CrossRef] [PubMed]
- Mebatsion, T. Extensive attenuation of rabies virus by simultaneously modifying the dynein light chain binding site in the P protein and replacing Arg333 in the G protein. J. Virol. 2001, 75, 11496–11502. [Google Scholar] [CrossRef] [PubMed]
- Visser, N.; Mebatsion, T.; Kilari, S. Oral rabies vaccine: Developing a safer and efficacious oral vaccine for dogs. In Proceedings of the SEARG Meeting, Windhoek, Namibia, January 2006; pp. 22–26. [Google Scholar]
- Prevec, L.; Campbell, J.B.; Christie, B.S.; Belbeck, L.; Graham, F.L. A recombinant human adenovirus vaccine against rabies. J. Infect. Dis. 1990, 161, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Yarosh, O.K.; Wandeler, A.I.; Graham, F.L.; Campbell, J.B.; Prevec, L. Human adenovirus type 5 vectors expressing rabies glycoprotein. Vaccine 1996, 14, 1257–1264. [Google Scholar] [CrossRef]
- Cox, J.H.; Schneider, L.G. Prophylactic immunization of humans against rabies by intradermal inoculation of human diploid cell culture vaccine. J. Clin. Microbiol. 1976, 3, 96–101. [Google Scholar] [PubMed]
- Müller, T.; Selhorst, T.; Burow, J.; Schameitat, A.; Vos, A. Cross-reactive antigenicity in orally vaccinated foxes and raccoon dogs against European bat lyssavirus type 1 and 2. Dev. Biol. 2006, 125, 195–204. [Google Scholar]
- Smith, J.S.; Yager, P.A.; Baer, G.M. A rapid reproducible test for determining rabies neutralizing antibody. Bull. World Health Organ. 1973, 48, 535–541. [Google Scholar] [PubMed]
- Smith, J.S.; Yager, P.A.; Baer, G.M. A rapid fluorescent focus inhibition test (RFFIT) for determining rabies virus-neutralizing antibody. In Laboratory Techniques in Rabies, 4th ed.; Meslin, F.X., Kaplan, M.M., Koprowski, H., Eds.; World Health Organization: Geneva, Switzerland, 1996; pp. 181–192. [Google Scholar]
- Habel, K. Habel, K. Habel test for potency. In Laboratory Techniques in Rabies, 4th ed.; Meslin, F.X., Kaplan, M.M., Koprowski, H., Eds.; World Health Organization: Geneva, Switzerland, 1996; pp. 369–373. [Google Scholar]
- Kostense, S.; Moore, S.; Companjen, A.; Bakker, A.B.; Marissen, W.E.; von, E.R.; Weverling, G.J.; Hanlon, C.; Goudsmit, J. Validation of the rapid fluorescent focus inhibition test (RFFIT) for rabies virus neutralizing antibodies in clinical samples. Antimicrob. Agents Chemother. 2012, 56, 3524–3530. [Google Scholar] [CrossRef] [PubMed]
- Mojzis, M.; Korytar, P.; Jerg, S. Development and Validation of ELISA Test for Detection of Rabies Anti-Glycoprotein Antibodies. In Internationals Conference on Rabies in the Americas (RITA XIX); Centers for Disease Control and Prevention: Atlanta, GA, USA, 2008; pp. 48–49. [Google Scholar]
- Wasniewski, M.; Guiot, A.L.; Schereffer, J.L.; Tribout, L.; Mahar, K.; Cliquet, F. Evaluation of an ELISA to detect rabies antibodies in orally vaccinated foxes and raccoon dogs sampled in the field. J. Virol. Methods 2013, 187, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Cliquet, F.; Sagne, L.; Schereffer, J.L.; Aubert, M.F. ELISA test for rabies antibody titration in orally vaccinated foxes sampled in the fields. Vaccine 2000, 18, 3272–3279. [Google Scholar] [CrossRef]
- Zalan, E.; Wilson, C.; Pukitis, D. A microtest for the quantitation of rabies virus neutralizing antibodies. J. Biol. Stand. 1979, 7, 213–220. [Google Scholar] [CrossRef]
- Rabies vaccine, live virus. In 9, Regulations; Code of Federal Regulations (annual edition); U.S. National Archives and Records Administration: City of College Park, Maryland, MD, USA, 2007; p. 4.
- Council of Europe. Rabies vaccine (live, oral) for foxes and raccoon dogs-0746. In European Pharmacopoeia 8.0; EDQM: Strasbourg, France, 2014; pp. 1011–1012. [Google Scholar]
- Lafon, M. Evasive strategies in rabies virus infection. Adv. Virus Res. 2011, 79, 33–53. [Google Scholar] [PubMed]
- Dietzschold, B.; Morimoto, K.; Hooper, D.C. Mechanisms of virus-induced neuronal damage and the clearance of viruses from the cns. Curr. Top. Microbiol. Immunol. 2001, 253, 145–155. [Google Scholar] [PubMed]
- Morimoto, K.; Hooper, D.C.; Spitsin, S.; Koprowski, H.; Dietzschold, B. Pathogenicity of different rabies virus variants inversely correlates with apoptosis and rabies virus glycoprotein expression in infected primary neuron cultures. J. Virol. 1999, 73, 510–518. [Google Scholar] [PubMed]
- Knowlton, F.F.; Roetto, M.; Briggs, D. Serological responses of coyotes to two commercial rabies vaccines. J. Wildl. Dis. 2001, 37, 798–802. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, C.E.; Barrett, J.; Briggs, D.; Cliquet, F.; Fooks, A.R.; Lumlertdacha, B.; Meslin, F.X.; Muler, T.; Nel, L.H.; Schneider, C.; et al. Can rabies be eradicated? Dev. Biol. 2008, 131, 95–121. [Google Scholar]
- Moore, S.M.; Gordon, C.R.; Hanlon, C.A. Measures of rabies immunity. In Rabies, 3rd ed.; Jackson, A.C., Ed.; Elsevier: San Diego, CA, USA, 2013; pp. 461–496. [Google Scholar]
- Smith, J. Rabies serology. In The Natural History of Rabies, 2nd ed.; Baer, G.M., Ed.; 1991; pp. 235–252. [Google Scholar]
- Moore, S.M.; Ricke, T.A.; Davis, R.D.; Briggs, D.J. The influence of homologous vs. Heterologous challenge virus strains on the serological test results of rabies virus neutralizing assays. Biologicals 2005, 33, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.J.; Ma, X.J.; Wang, L.H.; Li, H.; Shen, X.X.; Yu, P.C.; Tang, Q.; Liang, G.D. Preparation and initial application of a monoclonal antibody specific for a newly discovered conserved linear epitope of rabies virus nucleoprotein. Biomed. Environ. Sci. 2012, 25, 98–103. [Google Scholar] [PubMed]
- Wasniewski, M.; Almeida, I.; Baur, A.; Bedekovic, T.; Boncea, D.; Chaves, L.B.; David, D.; De, B.P.; Dobrostana, M.; Giraud, P.; et al. First international collaborative study to evaluate rabies antibody detection method for use in monitoring the effectiveness of oral vaccination programmes in fox and raccoon dog in europe. J. Virol. Methods 2016, 238, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Cleaveland, S.; Barrat, J.; Barrat, M.J.; Selve, M.; Kaare, M.; Esterhuysen, J. A rabies serosurvey of domestic dogs in rural Tanzania: Results of a rapid fluorescent focus inhibition test (RFFIT) and a liquid-phase blocking ELISA used in parallel. Epidemiol. Infect. 1999, 123, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Fehlner-Gardiner, C.; Rudd, R.; Donovan, D.; Slate, D.; Kempf, L.; Badcock, J. Comparing ONRAB® and Raboral V-RG® oral rabies vaccine field performance in raccoons and striped skunks, New Brunswick, Canada, and Maine, USA. J. Wildl. Dis. 2012, 48, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, A.T.; Fooks, A.R.; Hayman, D.T.; Horton, D.L.; Müller, T.; Plowright, R.; Peel, A.J.; Bowen, R.; Wood, J.L.; Mills, J.; et al. Deciphering serology to understand the ecology of infectious diseases in wildlife. Ecohealth 2013, 10, 298–313. [Google Scholar] [CrossRef] [PubMed]
- Fry, T.L.; Vandalen, K.K.; Shriner, S.A.; Moore, S.M.; Hanlon, C.A.; Vercauteren, K.C. Humoral immune response to oral rabies vaccination in raccoon kits: Problems and implications. Vaccine 2013, 31, 2811–2815. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Animal Species | Number Vaccinated | Number Controls | Challenge After … Days | Challenge Dose (MICLD50 log) | Challenge Virus Strain | Facility Where Research Was Done | Reference |
|---|---|---|---|---|---|---|---|---|
| 1 | arctic fox | 6 | 5 | 56 | 5.7 | fox | 5 | 30 |
| 2 | arctic fox | 8 | 4 | 112 | 5.7 | canine/fox | 5 | 27 |
| 3 | arctic fox | 10 | 4 | 56 | 3.9 | fox | 5 | 28 |
| 4 | arctic fox | 6 | 6 | 56 | 5.7 | fox | 5 | 29 |
| 5 | domestic dog | 18 | 6 | 30 | 3.8 | dog | 10 | 38 |
| 6 | domestic dog | 21 | 10 | 109 | 6.5 | dog/Ariana | 10 | 39 |
| 7 | domestic dog | 40 | 10 | 180 | 6.5 | dog | 11 | 41 |
| 8 | domestic dog | 12 | 6 | 28 | 7.4 | dog | 11 | 51 |
| 9 | domestic dog | 30 | 12 | 35 | 7.4 | dog | 11 | 48 |
| 10 | domestic dog | 12 | 7 | 35 | 3.7 | unknown | 13 | 52 |
| 11 | mongoose | 5 | 5 | 28 | 5 | skunk | 11 | 46 |
| 12 | raccoon | 5 | 5 | 30 | 4.9 | raccoon | 11 | 44 |
| 13 | raccoon | 7 | 7 | 28 | 4.9 | raccoon | 11 | 45 |
| 14 | raccoon | 14 | 5 | 56 | 4.9 | raccoon | 11 | 47 |
| 15 | raccoon | 61 | 19 | 442 | 6.7 | raccoon | 4 | 49 |
| 16 | raccoon | 30 | 10 | 350 | 5.9 | raccoon | 4 | 24 |
| 17 | raccoon | 10 | 5 | 60 | * | raccoon | 11 | 49 |
| 18 | raccoon | 6 | 13 | 28/65 | 5.5 | dog | 12 | 53 |
| 5 | raccoon dog | 6 | 9 | 124 | 2.8 | coyote | 10 | 38 |
| 19 | red fox | 36 | 6 | 34 | 4.7 | fox | 1 | 19 |
| 20 | red fox | 84 | 31 | 30 | 4 | fox | 2 | 20 |
| 21 | red fox | 66 | 25 | 730 | 4 | fox | 2 | 21 |
| 22 | red fox | 27 | 4 | 28 | 3.8 | fox | 1 | 22 |
| 23 | red fox | 13 | 4 | 28/180/360 | 3.2 | fox | 3 | 23 |
| 24 | red fox | 25 | 9 | 547 | 4.9 | fox | 4 | 26 |
| 25 | red fox | 18 | 6 | 2490 | 6.7 | fox | 4 | 31 |
| 26 | red fox | 25 | 0 | 28 | 5.7 | fox | 6 | 32 |
| 27 | red fox | 27 | 14 | 190 | 5.3 | canine/coyote | 7 | 33 |
| 28 | red fox | 8 | 2 | 45 | 5.1 | canine/coyote | 9 | 37 |
| 16 | red fox | 6 | 14 | 97 | 3 | fox | 10 | 38 |
| 29 | red fox | 16 | 8 | 107 | 3.3 | fox | 8 | 40 |
| 30 | red fox | 16 | 8 | 16 | 3.2 | fox | 8 | 35 |
| 28 | striped skunk | 3 | 2 | 45 | 5.1 | canine/coyote | 9 | 37 |
| 31 | striped skunk | 54 | 10 | 247 | 4.9 | arctic fox | 4 | 25 |
| 29 | striped skunk | 32 | 8 | 90/107 | 7.8 | skunk | 8 | 40 |
| 32 | striped skunk | 6 | 9 | 16 | 5.5 | skunk | 11 | 42 |
| 33 | striped skunk | 24 | 6 | 116 | 6.3 | skunk | 11 | 43 |
| 12 | striped skunk | 5 | 5 | 30 | 5 | skunk | 11 | 44 |
| 13 | striped skunk | 17 | 6 | 35 | 4.2 | skunk | 11 | 45 |
| 34 | striped skunk | 26 | 8 | 90 | 6.3 | skunk | 8 | 36 |
| 35 | striped skunk | 32 | 8 | 90 | 5.3 | skunk | 8 | 34 |
| Study | Animal Species | Vacc. | Ctls. | Challenge after … Days | Challenge Dose (MICLD50 log) | Challenge Virus Strain | Facil. | Approval | Date (dd/mm/yy) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | domestic dog | 16 | 4 | 56 | 3.6 | dog | IDT | 42502-3-710 IDT | 2/26/2014 |
| 2 | mongoose | 15 | 4 | 56 | 4.2 | dog | IDT | 42502-3-693 IDT | 24/06/2013 |
| 3 | mongoose | 11 | 3 | 42 | 4 | dog | IDT | 42502-3-693 IDT | 6/24/2013 |
| 4 | raccoon | 5 | 3 | 49 | 4.7 | coyote | IDT | 42502-3-582 IDT | 28/10/2009 |
| 5 | raccoon | 3 | - | 56 | 5 | coyote | IDT | 42502-3-582 IDT | 28/10/2009 |
| 6 | raccoon | 20 | 4 | 56 | 4.6 | dog | IDT | 42502-3-669 IDT | 11/23/2012 |
| 7 | raccoon | - | 12 | - | 4.2–6.2 | raccoon | IDT | 42502-3-726 IDT | 29/07/2014 |
| 8 | raccoon | 11 | 6 | 180 | 6.2 | dog(fox) | NWRC | QA2278A | 4/21/2014 |
| 9 | raccoon | 23 | 6 | 180 | 5.9 | dog(fox) | NWRC | QA2278B | 4/21/2014 |
| 10 | raccoon dog | 6 | 2 | 56 | 5 | coyote | IDT | 42502-3-669 IDT | 11/23/2012 |
| 11 | raccoon dog | 30 | 12 | 190 | 0.7 | fox | FLI | 7221.3-2-007/14 | 4/1/2014 |
| 12 | raccoon dog | - | 22 | - | 2.3–3.0 | fox | IDT | 42502-3-741 IDT | 8/17/2015 |
| 13 | raccoon dog | 16 | 2 | 28 | 2.0–2.7 | fox | IDT | 42502-3-761 IDT | 17/08/2015 |
| 14 | raccoon dog | - | 20 | - | 1.7–2.7 | fox | IDT | 42502-3-761 IDT | 17/08/2015 |
| 15 | raccoon dog | 30 | 12 | 183/184 | 3 | fox | FLI | 7221.3-2-018/15 | 17/06/2015 |
| 16 | red fox | 12 | 3 | 98 | 5 | coyote | IDT | Unknown * | Unknown * |
| 17 | red fox | 6 | 2 | 62 | 5 | coyote | IDT | 42502-3-582 IDT | 28/10/2009 |
| 18 | red fox | 6 | 2 | 56 | 5 | coyote | IDT | 42502-3-669 IDT | 11/23/2012 |
| 19 | red fox | 6 | 4 | 56 | 6.7 | dog | IDT | 42502-3-669 IDT | 11/23/2012 |
| 20 | red fox | 9 | 2 | 56 | 4.6 | fox | IDT | 42502-3-669 IDT | 11/23/2012 |
| 21 | red fox | - | 18 | - | 2.0–4.7 | fox | IDT | 42502-3-735 IDT | 17/08/2015 |
| 22 | red fox | 20 | 4 | 50 | 3 | fox | FLI | 7221.3-2-005/16 | 1/28/2016 |
| 23 | red fox | 30 | 12 | 190 | 0.7 | fox | FLI | 7221.3-2-007/14 | 4/1/2014 |
| 24 | striped skunk | 8 | 2 | 42 | 5 | coyote | IDT | 42502-3-582 IDT | 28/10/2009 |
| 25 | striped skunk | 5 | 3 | 58 | 4.7 | coyote | IDT | 42502-3-582 IDT | 28/10/2009 |
| 26 | striped skunk | 6 | 3 | 56 | 5 | coyote | IDT | 42502-3-582 IDT | 28/10/2009 |
| 27 | striped skunk | 16 | 4 | 28/56 | 4.6 | dog | IDT | 42502-3-669 IDT | 11/23/2012 |
| 28 | striped skunk | 20 | 4 | 335 | 5.9 | dog(fox) | NWRC | QA2278 | 8/15/2014 |
| Diagnostic Test | OR | (95% CI) | Se | (95% CI) | Sp | (95% CI) | PPV | (95% CI) | NPV | (95% CI) | p Value | Kappa | (95% CI) | Agreement |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| blocking ELISA (70% Inhib) | 18.54 | 4.982–78.59 | 0.4873 | 0.4486–0.5263 | 0.9512 | 0.8386–0.9913 | 0.9935 | 0.9768–0.9989 | 0.1074 | 0.07959–0.1435 | <0.0001 | 0.016 | 0.062–0.126 | Poor |
| blocking ELISA (40% Inhib) | 56.39 | 21.5–148 | 0.9063 | 0.8567–0.9399 | 0.8537 | 0.7156–0.9312 | 0.9667 | 0.9292–0.9846 | 0.6604 | 0.5259–0.7731 | <0.0001 | 0.681 | 0.564–0.799 | Good |
| Fox | 126 | 14.48–1383 | 0.9825 | 0.9071–0.9991 | 0.6923 | 0.4237–0.8732 | 0.9333 | 0.8407–0.9738 | 0.9 | 0.5958–0.9949 | <0.0001 | 0.741 | 0.527–0.955 | good |
| Raccoon dog | ∞ | 12.29–∞ | 0.8795 | 0.7922–0.9332 | 1 | 0.6756–1 | 1 | 0.95–1 | 0.4444 | 0.2456–0.6628 | <0.0001 | 0.562 | 0.332–0.792 | moderate |
| Mongoose | ∞ | 9.259–∞ | 1 | 0.7225–1 | 1 | 0.6457–1 | 1 | 0.7225–1 | 1 | 0.6457–1 | <0.0001 | 1.000 | 1.000-1.000 | perfect |
| Skunk | 6 | 0.8095–41.37 | 0.8 | 0.5481–0.9295 | 0.6 | 0.2307–0.9289 | 0.8571 | 0.6006–0.9746 | 0.5 | 0.1876–0.8124 | 0.1313 | 0.375 | 0.071–0.821 | fair |
| Dog | N/A | N/A | 0.375 | 0.1848–0.6136 | N/A | N/A | 1 | 0.6097–1 | 0 | 0–0.2775 | >0.9999 | 0.000 | 0.000-0.000 | poor |
| Raccoon | ∞ | 2.432–∞ | 0.6364 | 0.3538–0.8483 | 1 | 0.6756–1 | 1 | 0.6457–1 | 0.6667 | 0.3906–0.8619 | 0.0128 | -0.390 | -0.676–-0.104 | worse |
| RFFIT (0.5 IU/mL) | 32.1 | 10.2–101.1 | 0.7171 | 0.6519–0.7743 | 0.9268 | 0.8057–0.9748 | 0.98 | 0.9429–0.9945 | 0.3958 | 0.3038–0.4958 | <0.0001 | 0.419 | 0.312–0.526 | Moderate |
| Fox | 16.25 | 3.735–60.62 | 0.8298 | 0.6986–0.9111 | 0.7692 | 0.4974–0.9182 | 0.9286 | 0.8099–0.9754 | 0.5556 | 0.3372–0.7544 | 0.0001 | 0.526 | 0.285–0.767 | moderate |
| Raccoon dog | ∞ | 7.174–∞ | 0.7952 | 0.6962–0.868 | 1 | 0.6756–1 | 1 | 0.945–1 | 0.32 | 0.1721–0.5159 | <0.0001 | 0.406 | 0.201–0.611 | moderate |
| Mongoose | ∞ | 1.121–∞ | 0.3793 | 0.2269–0.56 | 1 | 0.6457–1 | 1 | 0.7412–1 | 0.28 | 0.1428–0.4758 | 0.0756 | 0.192 | 0.037–0.348 | poor |
| Skunk | ∞ | 4.573–∞ | 0.8571 | 0.6536–0.9502 | 1 | 0.5655–1 | 1 | 0.8241–1 | 0.625 | 0.3057–0.8632 | 0.0009 | 0.698 | 0.391–1.000 | good |
| Dog | N/A | N/A | 0.5 | 0.28–0.72 | N/A | N/A | 1 | 0.6756–1 | 0 | 0–0.3244 | >0.9999 | 0.000 | N/A | poor |
| Raccoon | ∞ | 0.6622–∞ | 0.2941 | 0.1328–0.5313 | 1 | 0.6756–1 | 1 | 0.5655–1 | 0.4 | 0.2188–0.6134 | 0.1399 | 0.211 | 0.013–0.408 | fair |
| RFFIT (0.25 IU/mL) | 24.83 | 9.924–59.72 | 0.8098 | 0.7505–0.8576 | 0.8537 | 0.7156–0.9312 | 0.9651 | 0.926–0.9839 | 0.473 | 0.3634–0.5852 | <0.0001 | 0.502 | 0.382–0.622 | Moderate |
| Fox | 5.415 | 1.584–18.03 | 0.7719 | 0.6479–0.8616 | 0.6154 | 0.3552–0.8229 | 0.898 | 0.7824–0.9556 | 0.381 | 0.2075–0.5912 | 0.0152 | 0.313 | 0.072–0.554 | fair |
| Raccoon dog | ∞ | 9.542–∞ | 0.8434 | 0.7502–0.9061 | 1 | 0.6756–1 | 1 | 0.948–1 | 0.381 | 0.2075–0.5912 | <0.0001 | 0.486 | 0.265–0.708 | moderate |
| Mongoose | ∞ | 2–∞ | 1 | 0.7412–1 | 0.8571 | 0.4869–0.9927 | 0.9167 | 0.6461–0.9957 | 1 | 0.6097–1 | 0.0004 | 0.880 | 0.653–1.000 | very good |
| Skunk | ∞ | 5.995–∞ | 0.9048 | 0.7109–0.9831 | 1 | 0.5655–1 | 1 | 0.8318–1 | 0.7143 | 0.3589–0.9492 | 0.0003 | 0.785 | 0.506–1.000 | good |
| Dog | N/A | N/A | 0.875 | 0.6398–0.9778 | N/A | N/A | 1 | 0.7847–1 | 0 | 0–0.8223 | >0.9999 | 0.000 | N/A | poor |
| Raccoon | ∞ | 1.618–∞ | 0.4706 | 0.2617–0.6904 | 1 | 0.6756–1 | 1 | 0.6756–1 | 0.4706 | 0.2617–0.6904 | 0.0261 | 0.363 | 0.106–0.619 | fair |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moore, S.M.; Gilbert, A.; Vos, A.; Freuling, C.M.; Ellis, C.; Kliemt, J.; Müller, T. Rabies Virus Antibodies from Oral Vaccination as a Correlate of Protection against Lethal Infection in Wildlife. Trop. Med. Infect. Dis. 2017, 2, 31. https://doi.org/10.3390/tropicalmed2030031
Moore SM, Gilbert A, Vos A, Freuling CM, Ellis C, Kliemt J, Müller T. Rabies Virus Antibodies from Oral Vaccination as a Correlate of Protection against Lethal Infection in Wildlife. Tropical Medicine and Infectious Disease. 2017; 2(3):31. https://doi.org/10.3390/tropicalmed2030031
Chicago/Turabian StyleMoore, Susan M., Amy Gilbert, Ad Vos, Conrad M. Freuling, Christine Ellis, Jeannette Kliemt, and Thomas Müller. 2017. "Rabies Virus Antibodies from Oral Vaccination as a Correlate of Protection against Lethal Infection in Wildlife" Tropical Medicine and Infectious Disease 2, no. 3: 31. https://doi.org/10.3390/tropicalmed2030031
APA StyleMoore, S. M., Gilbert, A., Vos, A., Freuling, C. M., Ellis, C., Kliemt, J., & Müller, T. (2017). Rabies Virus Antibodies from Oral Vaccination as a Correlate of Protection against Lethal Infection in Wildlife. Tropical Medicine and Infectious Disease, 2(3), 31. https://doi.org/10.3390/tropicalmed2030031