

Infectivity and Dissemination of Dengue Virus-1 in Different Aedes aegypti Populations Throughout Brazil

 , , , , , , ,
, , , , , , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
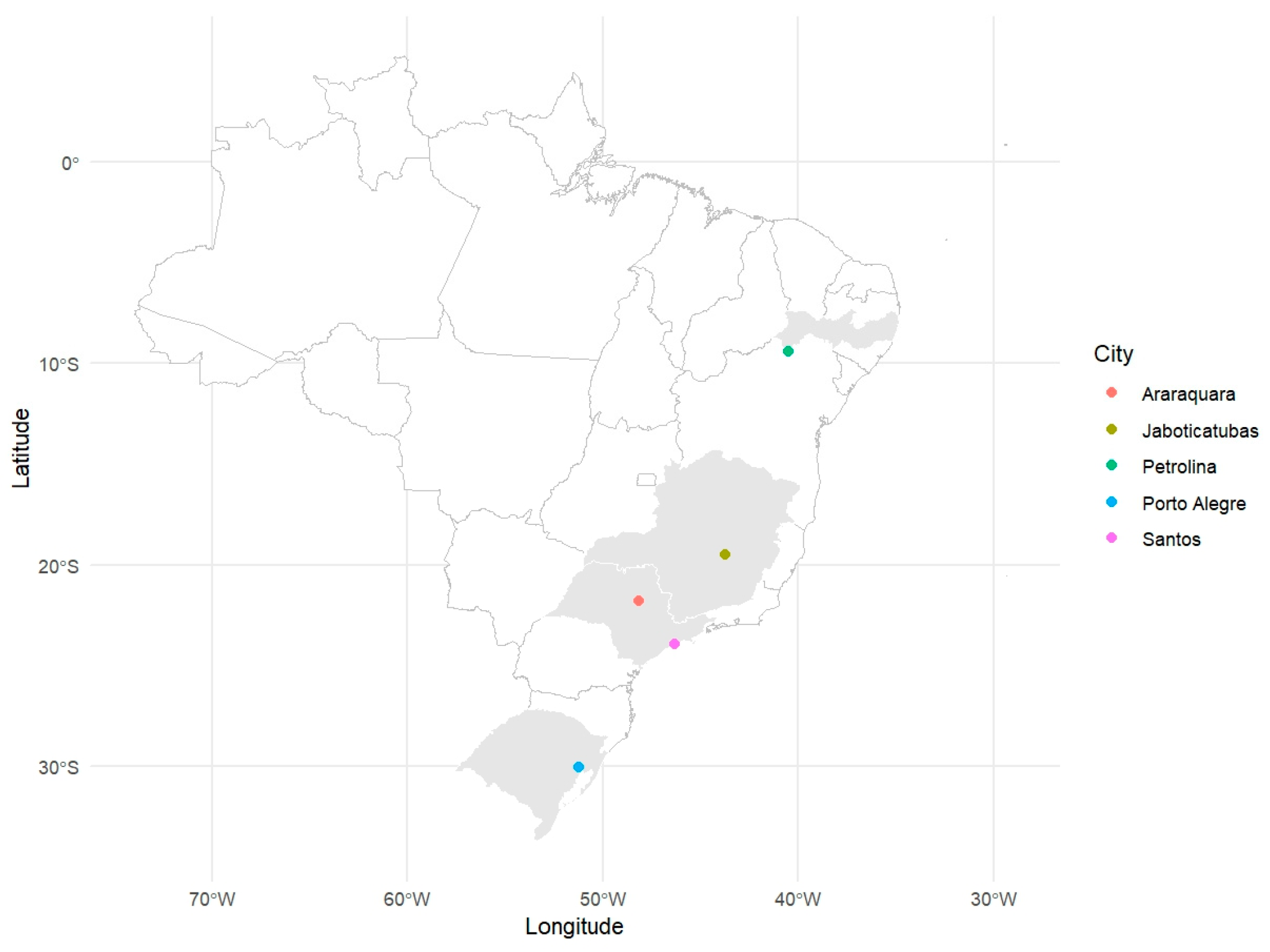
2.1. Mosquito Collection
2.2. Virus Strain
2.3. Mice Inoculation with DENV-1
2.4. Mosquito Infection with DENV-1
2.5. RNA Extraction and RT-qPCR
2.6. Statistical Analyses
3. Results
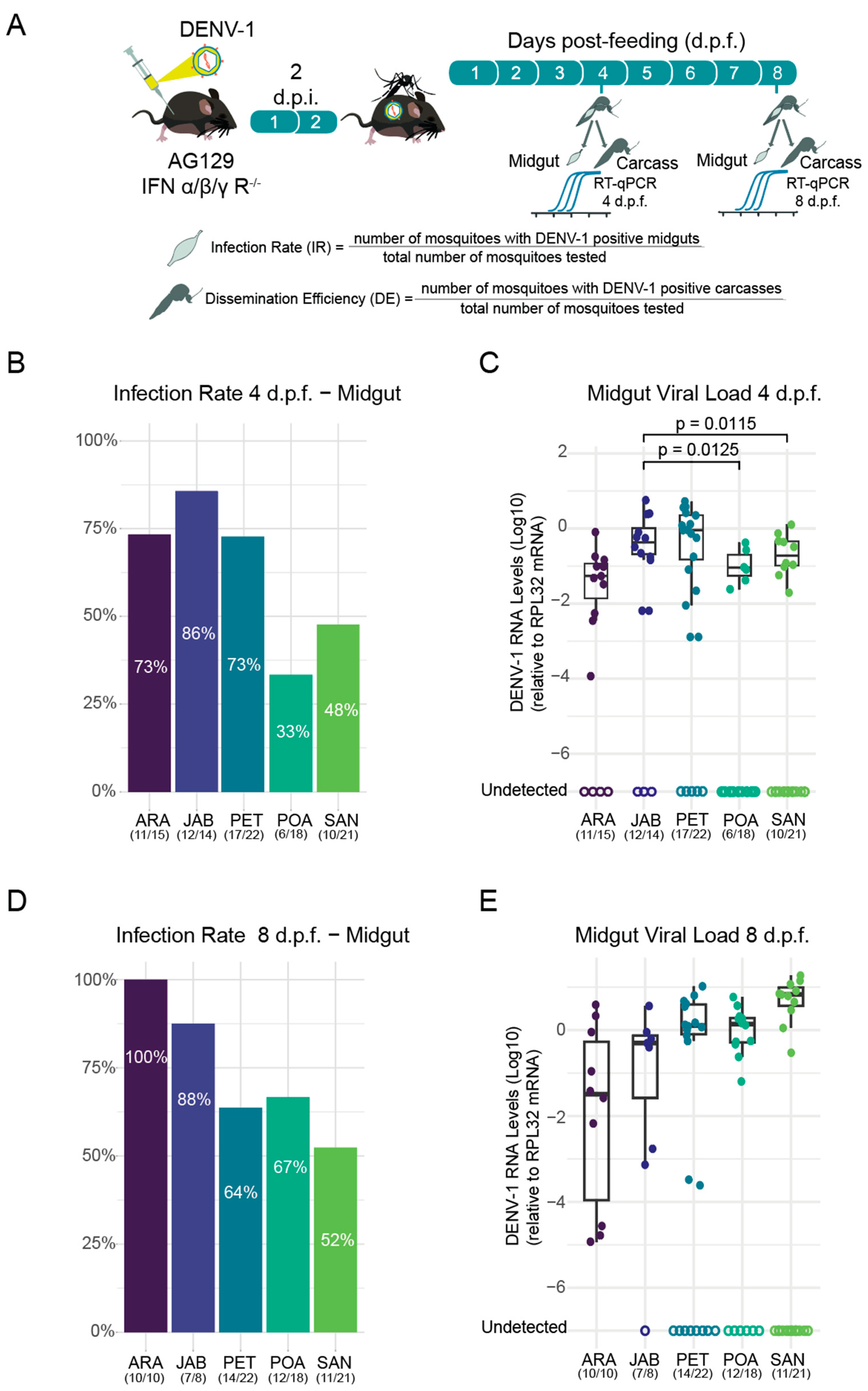
3.1. Differential Susceptibility of Brazilian Aedes aegypti Strains to DENV-1 Infection
3.2. Variation in DENV-1 Dissemination Rates Among Ae. aegypti Populations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Figueiredo, L.T.M. Emergent Arboviruses in Brazil. Rev. Soc. Bras. Med. Trop. 2007, 40, 224–229. [Google Scholar] [CrossRef]
- World Health Organization. Dengue and Severe Dengue; World Health Organization: Geneva, Switzerland, 2024; Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 24 February 2025).
- World Health Organization. Dengue—Global Situation; World Health Organization: Geneva, Switzerland, 2024; Available online: https://www.who.int/emergencies/disease-outbreak-news/item/2024-DON512 (accessed on 24 February 2025).
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The Global Distribution of the Arbovirus Vectors Aedes aegypti and Ae. Albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- Kok, B.H.; Lim, H.T.; Lim, C.P.; Lai, N.S.; Leow, C.Y.; Leow, C.H. Dengue Virus Infection—A Review of Pathogenesis, Vaccines, Diagnosis and Therapy. Virus Res. 2023, 324, 199018. [Google Scholar] [CrossRef]
- Postler, T.S.; Beer, M.; Blitvich, B.J.; Bukh, J.; de Lamballerie, X.; Drexler, J.F.; Imrie, A.; Kapoor, A.; Karganova, G.G.; Lemey, P.; et al. Renaming of the Genus Flavivirus to Orthoflavivirus and Extension of Binomial Species Names within the Family Flaviviridae. Arch. Virol. 2023, 168, 224. [Google Scholar] [CrossRef] [PubMed]
- Zanotto, P.M.; Gould, E.A.; Gao, G.F.; Harvey, P.H.; Holmes, E.C. Population Dynamics of Flaviviruses Revealed by Molecular Phylogenies. Proc. Natl. Acad. Sci. USA 1996, 93, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.; Kuhn, R.J. Structural Proteomics of Dengue Virus. Curr. Opin. Microbiol. 2008, 11, 369–377. [Google Scholar] [CrossRef]
- Lambrechts, L.; Scott, T.W.; Gubler, D.J. Consequences of the Expanding Global Distribution of Aedes Albopictus for Dengue Virus Transmission. PLoS Negl. Trop. Dis. 2010, 4, e646. [Google Scholar] [CrossRef]
- Weng, S.-C.; Tsao, P.-N.; Shiao, S.-H. Blood Glucose Promotes Dengue Virus Infection in the Mosquito Aedes aegypti. Parasites Vectors 2021, 14, 376. [Google Scholar] [CrossRef]
- Martina, B.E.E.; Koraka, P.; Osterhaus, A.D.M.E. Dengue Virus Pathogenesis: An Integrated View. Clin. Microbiol. Rev. 2009, 22, 564–581. [Google Scholar] [CrossRef]
- Raquin, V.; Lambrechts, L. Dengue Virus Replicates and Accumulates in Aedes aegypti Salivary Glands. Virology 2017, 507, 75–81. [Google Scholar] [CrossRef]
- Lambrechts, L.; Paaijmans, K.P.; Fansiri, T.; Carrington, L.B.; Kramer, L.D.; Thomas, M.B.; Scott, T.W. Impact of Daily Temperature Fluctuations on Dengue Virus Transmission by Aedes aegypti. Proc. Natl. Acad. Sci. USA 2011, 108, 7460–7465. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Kim, C.-H.; Chesser, L.; Ramirez, J.L.; Stone, C.M. Nutritional Stress Compromises Mosquito Fitness and Antiviral Immunity, While Enhancing Dengue Virus Infection Susceptibility. Commun. Biol. 2023, 6, 1123. [Google Scholar] [CrossRef]
- Gaye, A.; Wang, E.; Vasilakis, N.; Guzman, H.; Diallo, D.; Talla, C.; Ba, Y.; Dia, I.; Weaver, S.C.; Diallo, M. Potential for Sylvatic and Urban Aedes Mosquitoes from Senegal to Transmit the New Emerging Dengue Serotypes 1, 3 and 4 in West Africa. PLoS Negl. Trop. Dis. 2019, 13, e0007043. [Google Scholar] [CrossRef] [PubMed]
- Fansiri, T.; Fontaine, A.; Diancourt, L.; Caro, V.; Thaisomboonsuk, B.; Richardson, J.H.; Jarman, R.G.; Ponlawat, A.; Lambrechts, L. Genetic Mapping of Specific Interactions between Aedes aegypti Mosquitoes and Dengue Viruses. PLoS Genet. 2013, 9, e1003621. [Google Scholar] [CrossRef] [PubMed]
- Black, W.C.; Bennett, K.E.; Gorrochótegui-Escalante, N.; Barillas-Mury, C.V.; Fernández-Salas, I.; de Lourdes Muñoz, M.; Farfán-Alé, J.A.; Olson, K.E.; Beaty, B.J. Flavivirus Susceptibility in Aedes aegypti. Arch. Med. Res. 2002, 33, 379–388. [Google Scholar] [CrossRef]
- Failloux, A.-B.; Vazeille, M.; Rodhain, F. Geographic Genetic Variation in Populations of the Dengue Virus Vector Aedes aegypti. J. Mol. Evol. 2002, 55, 653–663. [Google Scholar] [CrossRef]
- Lequime, S.; Fontaine, A.; Ar Gouilh, M.; Moltini-Conclois, I.; Lambrechts, L. Genetic Drift, Purifying Selection and Vector Genotype Shape Dengue Virus Intra-Host Genetic Diversity in Mosquitoes. PLoS Genet. 2016, 12, e1006111. [Google Scholar] [CrossRef]
- Salazar, M.I.; Richardson, J.H.; Sánchez-Vargas, I.; Olson, K.E.; Beaty, B.J. Dengue Virus Type 2: Replication and Tropisms in Orally Infected Aedes aegypti Mosquitoes. BMC Microbiol. 2007, 7, 9. [Google Scholar] [CrossRef]
- Lourenço-de-Oliveira, R.; Vazeille, M.; De Filippis, A.M.B.; Failloux, A.B. Aedes aegypti in Brazil: Genetically Differentiated Populations with High Susceptibility to Dengue and Yellow Fever Viruses. Trans. R. Soc. Trop. Med. Hyg. 2004, 98, 43–54. [Google Scholar] [CrossRef]
- Chaves, B.A.; Godoy, R.S.M.; Campolina, T.B.; Júnior, A.B.V.; Paz, A.d.C.; Vaz, E.B.d.C.; Silva, B.M.; Nascimento, R.M.; Guerra, M.d.G.V.B.; Lacerda, M.V.G.; et al. Dengue Infection Susceptibility of Five Aedes aegypti Populations from Manaus (Brazil) after Challenge with Virus Serotypes 1–4. Viruses 2022, 14, 20. [Google Scholar] [CrossRef]
- Nogueira, R.M.R.; Araújo, J.M.G.; Schatzmayr, H.G. Dengue Viruses in Brazil, 1986–2006. Rev. Panam. Salud Publica 2007, 22, 358–363. [Google Scholar] [CrossRef]
- Breda, R.; Motta, A.S. Análise da influência de determinantes meteorológicos na periodicidade de epidemias de dengue em Porto Alegre. Ver. Inst. Adolfo Lutz. São Paulo 2024, 83, e39267. [Google Scholar] [CrossRef]
- dos Santos, S.D.; Ribeiro, M.C.S.A. Incidence of Dengue and Socioeconomic and Entomological Indicators in Santos, São Paulo, 2012–2016. Nurs. Ed. Bras. 2021, 24, 5229–5242. [Google Scholar] [CrossRef]
- De Freitas, J.R.; Santiago, E.J.P.; De Freitas, J.C.R.; Da Silva, A.S.A.; De Araújo Filho, R.N.; Piscoya, V.C.; Cunha Filho, M. Space-temporal analysis trend of the numbers of dengue cases in Pernambuco-Brazil. Res. Soc. Dev. 2020, 9, e526974427. [Google Scholar] [CrossRef]
- Bezerra, J.M.T.; de Sousa, S.C.; Tauil, P.L.; Carneiro, M.; Barbosa, D.S. Entry of dengue virus serotypes and their geographic distribution in Brazilian federative units: A systematic review. Rev. Bras. Epidemiol. 2021, 24, e210020. [Google Scholar] [CrossRef]
- Baldon, L.V.R.; de Mendonça, S.F.; Ferreira, F.V.; Rezende, F.O.; Amadou, S.C.G.; Leite, T.H.J.F.; Rocha, M.N.; Marques, J.T.; Moreira, L.A.; Ferreira, A.G.A. AG129 Mice as a Comprehensive Model for the Experimental Assessment of Mosquito Vector Competence for Arboviruses. Pathogens 2022, 11, 879. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control, 3rd ed.; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Olmo, R.P.; Ferreira, A.G.A.; Izidoro-Toledo, T.C.; Aguiar, E.R.G.R.; de Faria, I.J.S.; de Souza, K.P.R.; Osório, K.P.; Kuhn, L.; Hammann, P.; de Andrade, E.G.; et al. Control of dengue virus in the midgut of Aedes aegypti by ectopic expression of the dsRNA-binding protein Loqs2. Nat. Microbiol. 2018, 3, 1385–1393. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The Global Distribution and Burden of Dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Araújo, V.E.M.D.; Bezerra, J.M.T.; Amâncio, F.F.; Passos, V.M.D.A.; Carneiro, M. Aumento Da Carga de Dengue No Brasil e Unidades Federadas, 2000 e 2015: Análise Do Global Burden of Disease Study 2015. Rev. Bras. Epidemiol. 2017, 20, 205–216. [Google Scholar] [CrossRef]
- Villabona-Arenas, C.J.; De Oliveira, J.L.; Capra, C.D.S.; Balarini, K.; Loureiro, M.; Fonseca, C.R.T.P.; Passos, S.D.; Zanotto, P.M.D.A. Detection Of Four Dengue Serotypes Suggests Rise In Hyperendemicity In Urban Centers Of Brazil. PLoS Negl. Trop. Dis. 2014, 8, e2620. [Google Scholar] [CrossRef] [PubMed]
- Bastos, M.D.S.; Figueiredo, R.M.P.D.; Ramasawmy, R.; Itapirema, E.; Gimaque, J.B.L.; Santos, L.O.; Figueiredo, L.T.M.; Mourão, M.P.G. Simultaneous Circulation of All Four Dengue Serotypes in Manaus, State of Amazonas, Brazil in 2011. Rev. Soc. Bras. Med. Trop. 2012, 45, 393–394. [Google Scholar] [CrossRef] [PubMed]
- Gurgel-Gonçalves, R.; Oliveira, W.K.; Croda, J. The Greatest Dengue Epidemic in Brazil: Surveillance, Prevention, and Control. Rev. Soc. Bras. Med. Trop. 2024, 57, e002032024. [Google Scholar] [CrossRef]
- Carpenter, A.; Clem, R.J. Factors Affecting Arbovirus Midgut Escape in Mosquitoes. Pathogens 2023, 12, 220. [Google Scholar] [CrossRef] [PubMed]
- De Freitas, A.; Rezende, F.; De Mendonça, S.; Baldon, L.; Silva, E.; Ferreira, F.; Almeida, J.; Amadou, S.; Marçal, B.; Comini, S.; et al. The High Capacity of Brazilian Aedes aegypti Populations to Transmit a Locally Circulating Lineage of Chikungunya Virus. Viruses 2024, 16, 575. [Google Scholar] [CrossRef]
- Luna, E.J.A.; Figueiredo, G.M.; Levi, J.E.; Campos, S.R.S.L.C.; Felix, A.C.; Souza, N.S.E.; Figueiredo, W.M.; Costa, A.A.; Cardoso, M.R.A.; Pannuti, C.S. A Cohort Study to Assess the Incidence of Dengue, Brazil, 2014–2018. Acta Trop. 2020, 204, 105313. [Google Scholar] [CrossRef]
- Ferreira, A.C.; Chiaravalloti Neto, F.; Mondini, A. Dengue in Araraquara, State of São Paulo: Epidemiology, Climate and Aedes aegypti Infestation. Rev. Saúde Pública 2018, 52, 18. [Google Scholar] [CrossRef]
- De Souza, C.S.; Caleiro, G.S.; Claro, I.M.; De Jesus, J.G.; Coletti, T.M.; Da Silva, C.A.M.; Costa, Â.A.; Inenami, M.; Ribeiro, A.C.; Felix, A.C.; et al. Phylogenetics, Epidemiology and Temporal Patterns of Dengue Virus in Araraquara, São Paulo State. Viruses 2024, 16, 274. [Google Scholar] [CrossRef]
- Prefeitura de Jaboticatubas. Prefeitura Decreta Situação de Emergência Devido ao Aumento de Casos de Dengue no Município. 2024. Available online: https://www.jaboticatubas.mg.gov.br/index.php/mais-noticias/602-prefeitura-decreta-situa%C3%A7%C3%A3o-de-emerg%C3%AAncia-devido-ao-aumento-de-casos-de-dengue-no-munic%C3%ADpio.html (accessed on 24 February 2025).
- Adelino, T.É.R.; Giovanetti, M.; Fonseca, V.; Xavier, J.; De Abreu, Á.S.; Do Nascimento, V.A.; Demarchi, L.H.F.; Oliveira, M.A.A.; Da Silva, V.L.; De Mello, A.L.E.S.; et al. Field and Classroom Initiatives for Portable Sequence-Based Monitoring of Dengue Virus in Brazil. Nat. Commun. 2021, 12, 2296. [Google Scholar] [CrossRef]
- Do Nascimento, I.D.S.; Pastor, A.F.; Lopes, T.R.R.; Farias, P.C.S.; Gonçales, J.P.; Do Carmo, R.F.; Durães-Carvalho, R.; Da Silva, C.S.; Silva Júnior, J.V.J. Retrospective Cross-Sectional Observational Study on the Epidemiological Profile of Dengue Cases in Pernambuco State, Brazil, between 2015 and 2017. BMC Public Health 2020, 20, 923. [Google Scholar] [CrossRef] [PubMed]
- Carvalho-Leandro, D.; Ayres, C.F.J.; Guedes, D.R.D.; Suesdek, L.; Melo-Santos, M.A.V.; Oliveira, C.F.; Cordeiro, M.T.; Regis, L.N.; Marques, E.T.; Gil, L.H.; et al. Immune Transcript Variations among Aedes aegypti Populations with Distinct Susceptibility to Dengue Virus Serotype 2. Acta Trop. 2012, 124, 113–119. [Google Scholar] [CrossRef]
- Gregianini, T.S.; Tumioto-Giannini, G.L.; Favreto, C.; Plentz, L.C.; Ikuta, N.; Da Veiga, A.B.G. Dengue in Rio Grande Do Sul, Brazil: 2014 to 2016. Rev. Med. Virol. 2018, 28, e1960. [Google Scholar] [CrossRef] [PubMed]
- Da Cruz Ferreira, D.A.; Degener, C.M.; De Almeida Marques-Toledo, C.; Bendati, M.M.; Fetzer, L.O.; Teixeira, C.P.; Eiras, Á.E. Meteorological Variables and Mosquito Monitoring Are Good Predictors for Infestation Trends of Aedes aegypti, the Vector of Dengue, Chikungunya and Zika. Parasites Vectors 2017, 10, 78. [Google Scholar] [CrossRef]
- Guzzetta, G.; Marques-Toledo, C.A.; Rosà, R.; Teixeira, M.; Merler, S. Quantifying the Spatial Spread of Dengue in a Non-Endemic Brazilian Metropolis via Transmission Chain Reconstruction. Nat. Commun. 2018, 9, 2837. [Google Scholar] [CrossRef] [PubMed]
- Tumioto, G.L.; Gregianini, T.S.; Dambros, B.P.; Cestari, B.C.; Alves Nunes, Z.M.; Veiga, A.B.G. Laboratory Surveillance of Dengue in Rio Grande Do Sul, Brazil, from 2007 to 2013. PLoS ONE 2014, 9, e104394. [Google Scholar] [CrossRef]
- Bellegarde Fernandes, M.A.; Natal, D.; Domingos, M.D.F. Aspectos Epidemiológicos Da Transmissão De Dengue Em Santos, São Paulo, No Período De 1997 A 2012. J. Health Biol. Sci. 2014, 2, 5. [Google Scholar] [CrossRef]
- de Figueiredo, M.L.; Gomes, A.d.C.; Amarilla, A.A.; Leandro, A.d.S.; Orrico, A.d.S.; de Araujo, R.F. Mosquitoes Infected with Dengue Viruses in Brazil. Virol. J. 2010, 7, 152. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Name | City | State | Species | Stage | Number of Mosquitoes | Generation | Year |
|---|---|---|---|---|---|---|---|
| ARA | Araraquara | São Paulo | Aedes aegypti | eggs | 100 females + 100 males | F2 | 2022 |
| JAB | Jaboticatubas | Minas Gerais | Aedes aegypti | eggs | 100 females + 100 males | F2 | 2022 |
| PET | Petrolina | Pernambuco | Aedes aegypti | eggs | 100 females + 100 males | F2 | 2022 |
| POA | Porto Alegre | Rio Grande do Sul | Aedes aegypti | eggs | 100 females + 100 males | F2 | 2022 |
| SAN | Santos | São Paulo | Aedes aegypti | eggs | 100 females + 100 males | F2 | 2022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Freitas, A.C.; Santos, E.; Baldon, L.; de Mendonça, S.; Rezende, F.O.; Moreira, R.; Sousa, V.; Lima, M.; Silva, E.; Ferreira, F.; et al. Infectivity and Dissemination of Dengue Virus-1 in Different Aedes aegypti Populations Throughout Brazil. Trop. Med. Infect. Dis. 2025, 10, 112. https://doi.org/10.3390/tropicalmed10040112
de Freitas AC, Santos E, Baldon L, de Mendonça S, Rezende FO, Moreira R, Sousa V, Lima M, Silva E, Ferreira F, et al. Infectivity and Dissemination of Dengue Virus-1 in Different Aedes aegypti Populations Throughout Brazil. Tropical Medicine and Infectious Disease. 2025; 10(4):112. https://doi.org/10.3390/tropicalmed10040112
Chicago/Turabian Stylede Freitas, Amanda Cupertino, Ellen Santos, Lívia Baldon, Silvana de Mendonça, Fernanda Oliveira Rezende, Rafaela Moreira, Viviane Sousa, Mariana Lima, Emanuele Silva, Flávia Ferreira, and et al. 2025. "Infectivity and Dissemination of Dengue Virus-1 in Different Aedes aegypti Populations Throughout Brazil" Tropical Medicine and Infectious Disease 10, no. 4: 112. https://doi.org/10.3390/tropicalmed10040112
APA Stylede Freitas, A. C., Santos, E., Baldon, L., de Mendonça, S., Rezende, F. O., Moreira, R., Sousa, V., Lima, M., Silva, E., Ferreira, F., de Almeida, J. P. P., Amadou, S., Marçal, B., Comini, S., Rocha, M., Todjro, Y., Leite, T. J., Santos, V., de Faria, I. J. d. S., ... Ferreira, A. (2025). Infectivity and Dissemination of Dengue Virus-1 in Different Aedes aegypti Populations Throughout Brazil. Tropical Medicine and Infectious Disease, 10(4), 112. https://doi.org/10.3390/tropicalmed10040112