
Molecular Cloning and Characterization of Scavenger Receptor Class B Type 1 in Grass Carp (Ctenopharyngodon idellus) and Its Expression Profile following Grass Carp Reovirus Challenge


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Fish and Sampling
2.2. Gene Cloning and Sequence Analysis of SRB1 in C. idellus
2.3. GCRV Challenge and Sampling
2.4. Quantification of Gene Expression
2.5. Statistical Analysis
3. Results
3.1. Molecular Features of CiSRB1
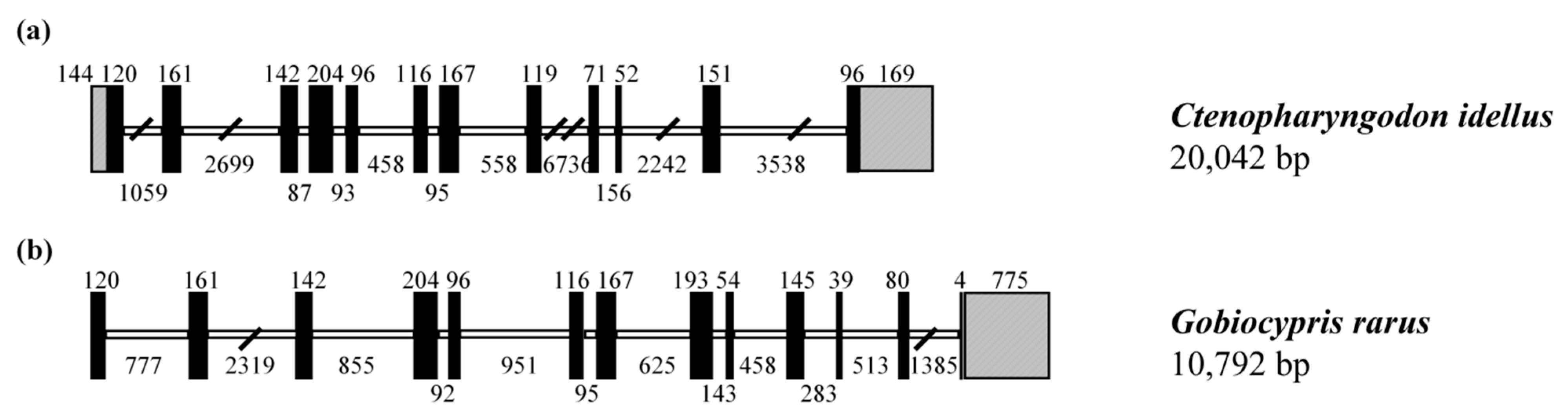
3.2. Homology Analysis of CiSRB1
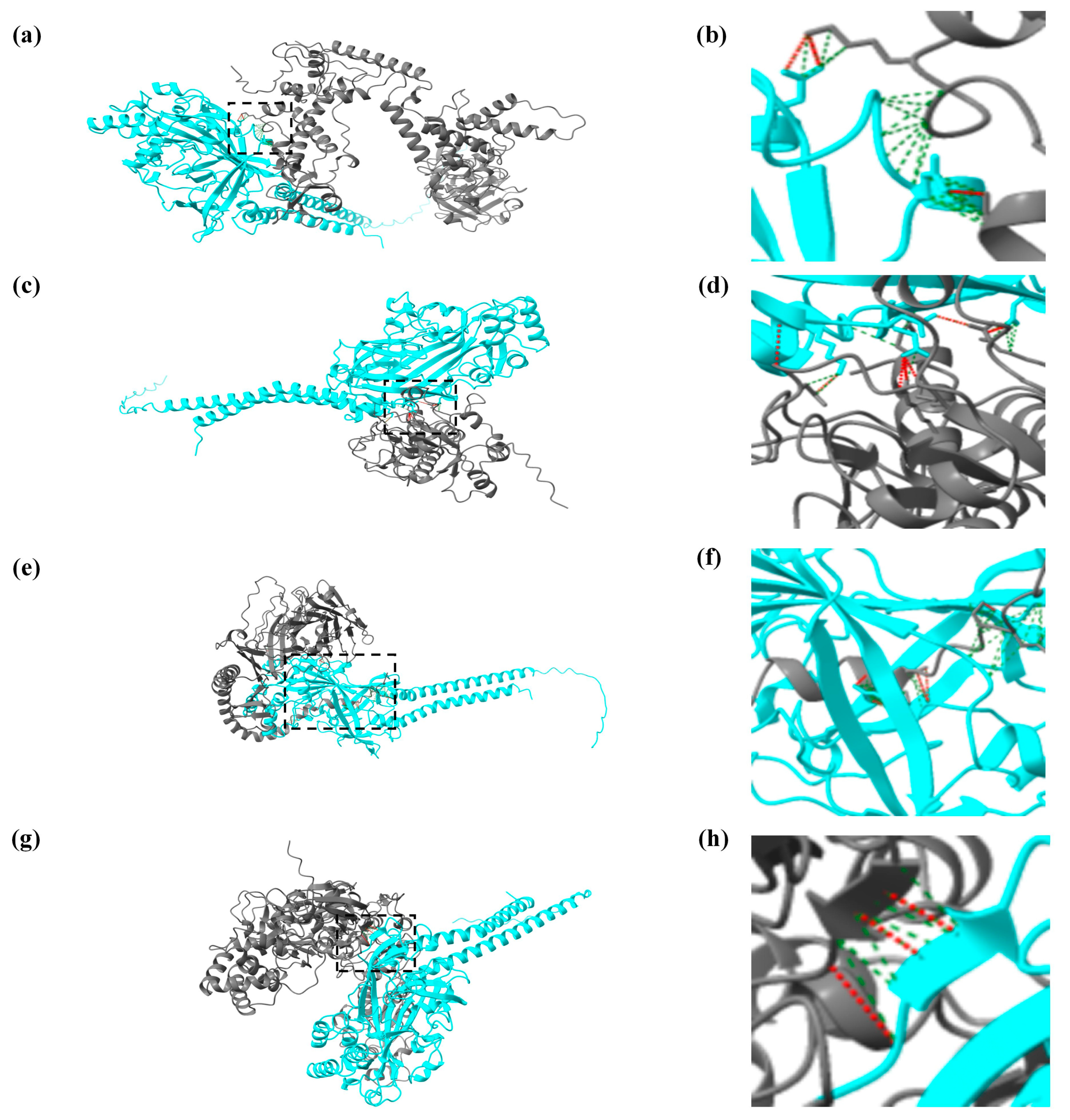
3.3. Predictive Analysis of Protein Interactions between CiSRB1 and GCRV Particles
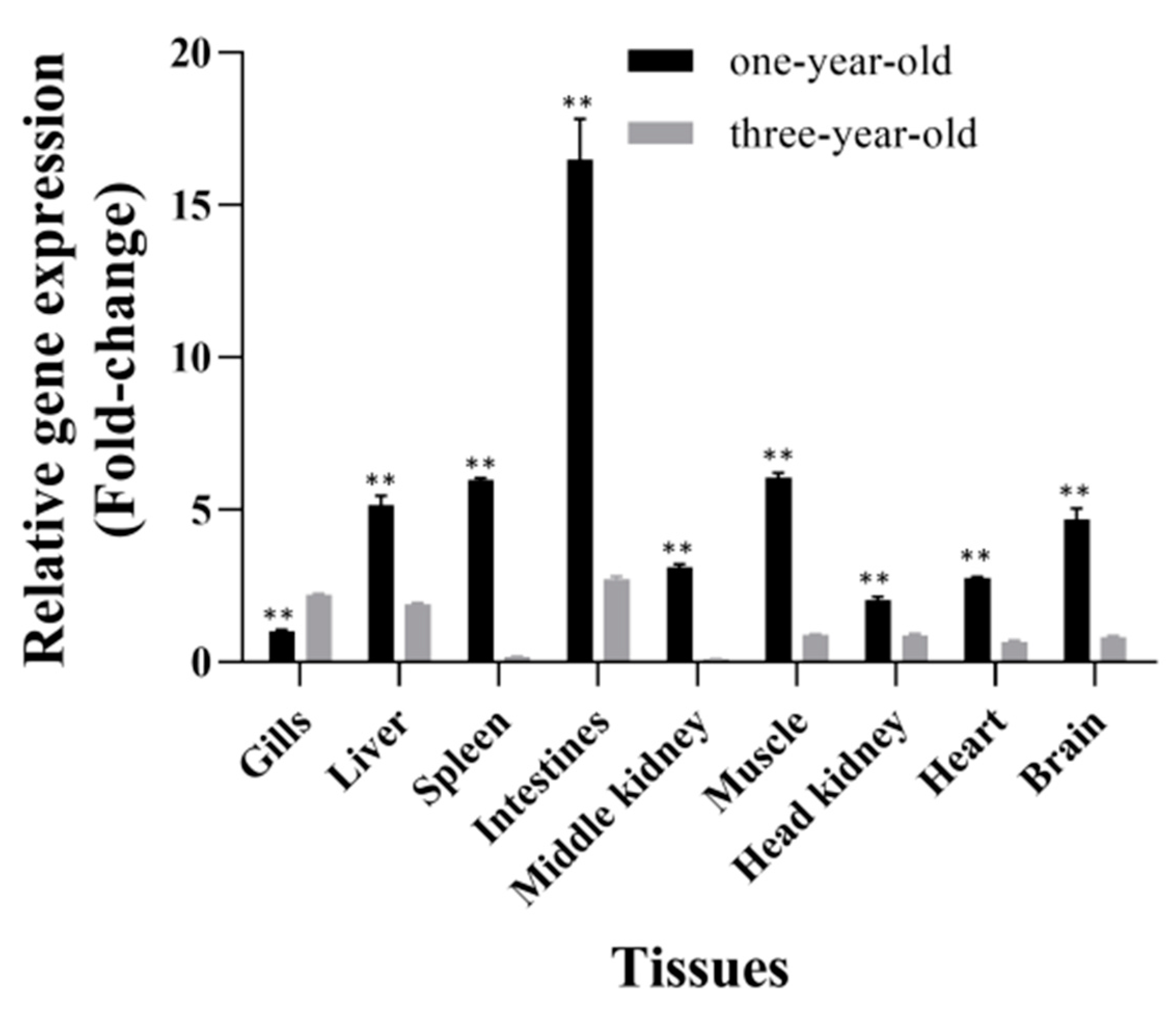
3.4. Tissue Distribution of CiSRB1 in Healthy Grass Carp
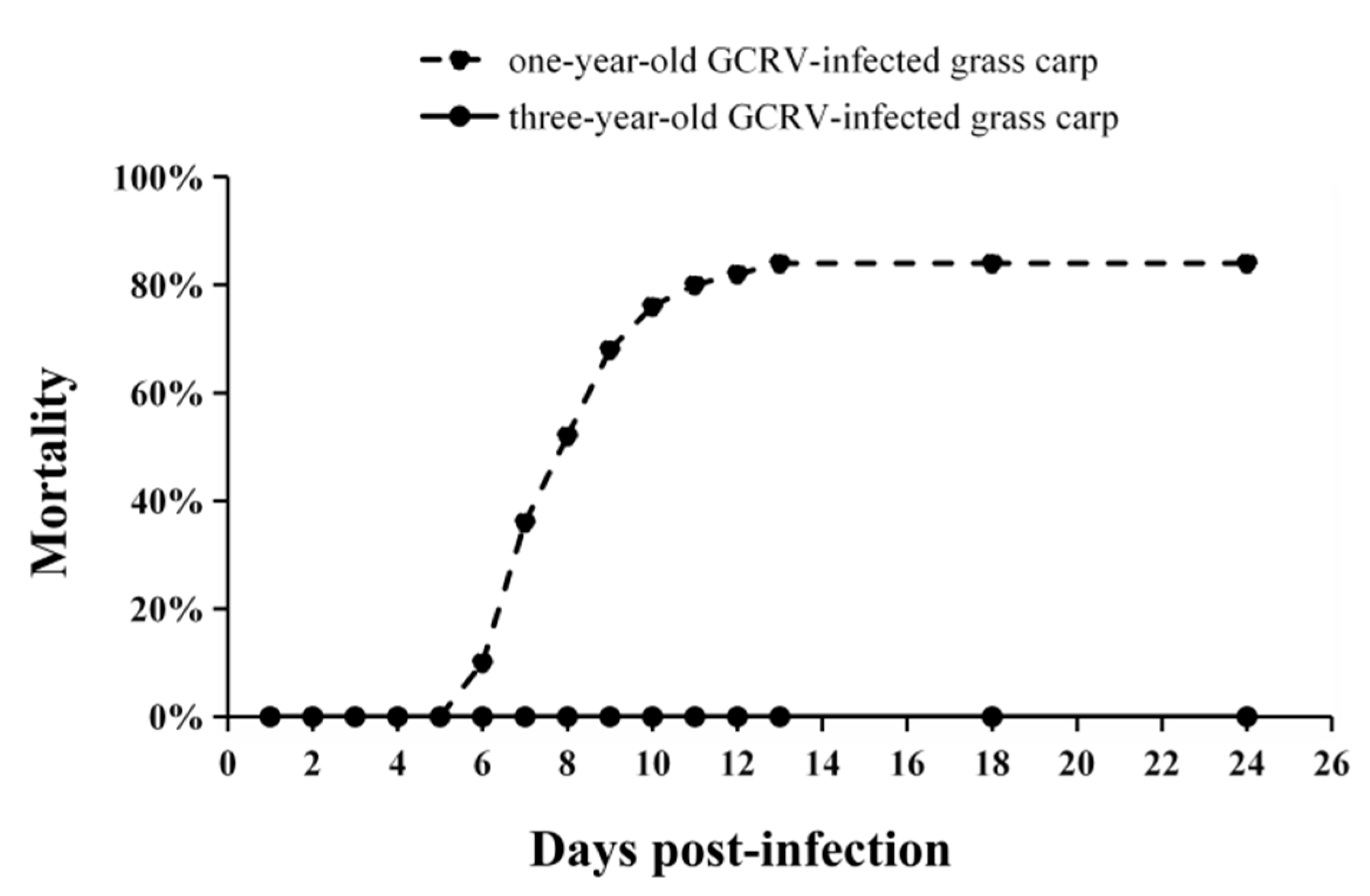
3.5. Mortality of Grass Carp after GCRV Infection
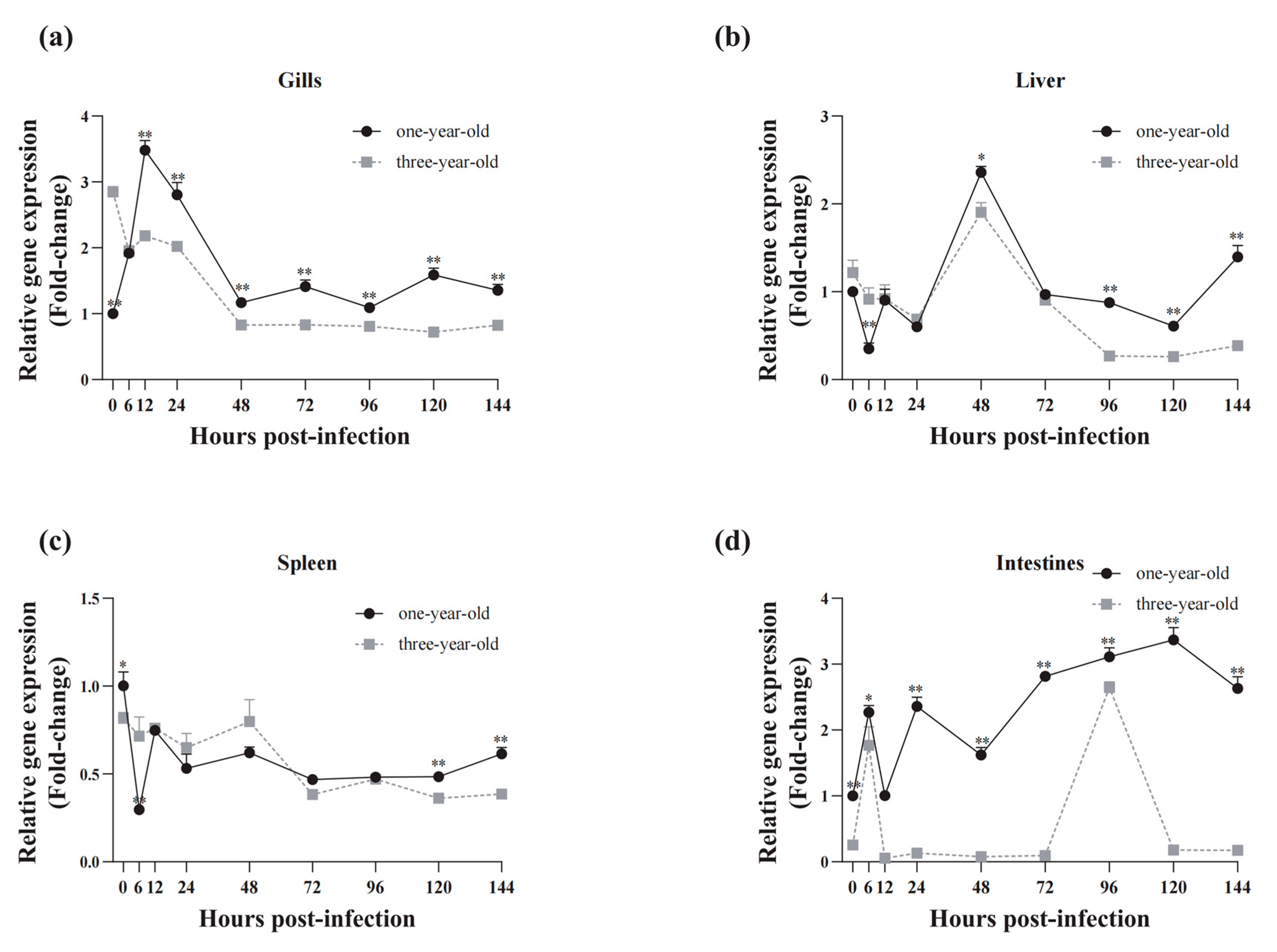
3.6. Expression Profiles of CiSRB1 following GCRV Infection
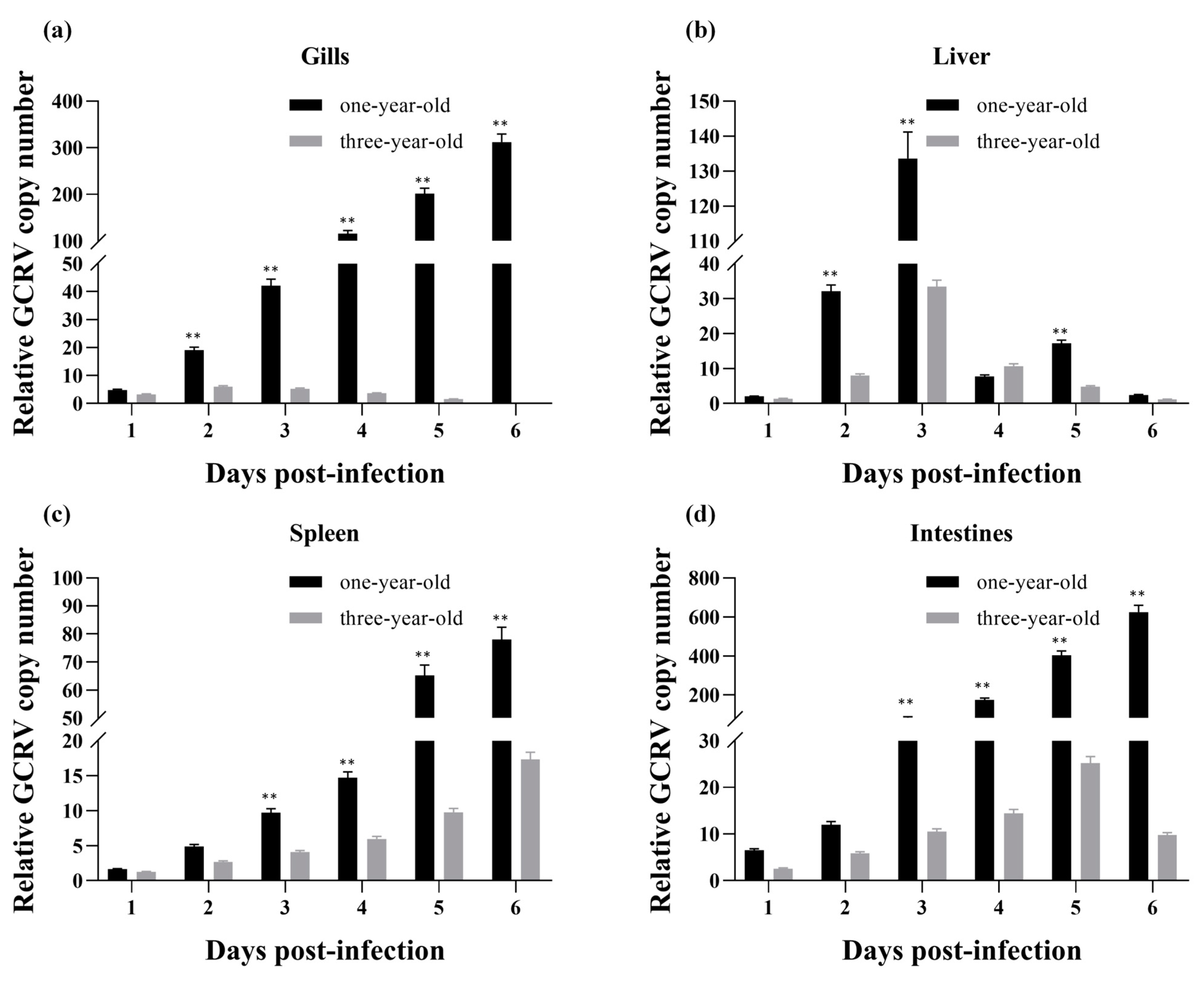
3.7. Relative Copy Numbers of GCRV RNA in Infected Grass Carp
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, L.; Wang, C.; Huang, Y.; Xu, B.; Liu, Y.; Yu, J.; Xiong, L.; Xiao, T.; Liu, Q. Identification of the C1qDC gene family in grass carp (Ctenopharyngodon idellus) and the response of C1qA, C1qB, and C1qC to GCRV infection in vivo and in vitro. Fish Shellfish Immunol. 2024, 148, 109477. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Su, J. Type II grass carp reovirus infects leukocytes but not erythrocytes and thrombocytes in grass carp (Ctenopharyngodon idella). Viruses 2021, 13, 870. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhao, W.; Ji, N.; Xu, D.; Li, Y.; Xiao, T.; Wang, J.; Zou, J. Analysis of tissue tropism of GCRV-II infection in grass carp using a VP35 monoclonal antibody. Dev. Comp. Immunol. 2024, 157, 105189. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Liao, L.; Xu, Q.; He, Z.; Xiao, T.; Wang, J.; Huang, J.; Yu, Y.; Wu, B.; Yan, Q. Host–Microbiota Interactions and Responses to Grass Carp Reovirus Infection in Ctenopharyngodon idellus. Environ. Microbiol. 2021, 23, 431–447. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, G.; Yuan, L.; Xia, X.-Q.; Zhang, W.; Shi, M. Transcriptomes of zebrafish in early stages of multiple viral invasions reveal the role of sterols in innate immune switch-on. Int. J. Mol. Sci. 2023, 24, 4427. [Google Scholar] [CrossRef]
- Ravi, S.; Martin, L.C.; Krishnan, M.; Kumaresan, M.; Manikandan, B.; Ramar, M. Interactions between macrophage membrane and lipid mediators during cardiovascular diseases with the implications of scavenger receptors. Chem. Phys. Lipids 2024, 258, 105362. [Google Scholar] [CrossRef]
- Vogel, A.; Brunner, J.S.; Hajto, A.; Sharif, O.; Schabbauer, G. Lipid scavenging macrophages and inflammation. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2022, 1867, 159066. [Google Scholar] [CrossRef]
- Huby, T.; Le Goff, W. Macrophage SR-B1 in atherosclerotic cardiovascular disease. Curr. Opin. Lipidol. 2022, 33, 167–174. [Google Scholar] [CrossRef]
- Barth, H.; Schnober, E.K.; Neumann-Haefelin, C.; Thumann, C.; Zeisel, M.B.; Diepolder, H.M.; Hu, Z.; Liang, T.J.; Blum, H.E.; Thimme, R.; et al. Scavenger receptor class B is required for hepatitis C virus uptake and cross-presentation by human dendritic cells. J. Virol. 2008, 82, 3466–3479. [Google Scholar] [CrossRef]
- Colpitts, C.C.; Baumert, T.F. SCARB1 variants and HCV infection: Host susceptibility is lost in translation. J. Hepatol. 2017, 67, 211–213. [Google Scholar] [CrossRef]
- Jennelle, L.T.; Magoro, T.; Angelucci, A.R.; Dandekar, A.; Hahn, Y.S. Hepatitis C virus alters macrophage cholesterol metabolism through interaction with scavenger receptors. Viral Immunol. 2022, 35, 223–235. [Google Scholar] [CrossRef]
- Li, Y.; Kakinami, C.; Li, Q.; Yang, B.; Li, H. Human apolipoprotein A-I is associated with dengue virus and enhances virus infection through SR-BI. PLoS ONE 2013, 8, e70390. [Google Scholar] [CrossRef]
- Langlois, A.-C.; Marinach, C.; Manzoni, G.; Silvie, O. Plasmodium sporozoites can invade hepatocytic cells independently of the ephrin receptor A2. PLoS ONE 2018, 13, e0200032. [Google Scholar] [CrossRef]
- Tang, Y.; Hu, L.; Liu, Y.; Zhou, B.; Qin, X.; Ye, J.; Shen, M.; Wu, Z.; Zhang, P. Possible mechanisms of cholesterol elevation aggravating COVID-19. Int. J. Med. Sci. 2021, 18, 3533–3543. [Google Scholar] [CrossRef]
- Chen, G.; He, L.; Luo, L.; Huang, R.; Liao, L.; Li, Y.; Zhu, Z.; Wang, Y. Transcriptomics sequencing provides insights into understanding the mechanism of grass carp reovirus infection. Int. J. Mol. Sci. 2018, 19, 488. [Google Scholar] [CrossRef] [PubMed]
- Ou, M.; Huang, R.; Luo, Q.; Xiong, L.; Chen, K.; Wang, Y. Characterisation of scavenger receptor class B type 1 in rare minnow (Gobiocypris rarus). Fish Shellfish Immunol. 2019, 89, 614–622. [Google Scholar] [CrossRef]
- Shen, W.-J.; Asthana, S.; Kraemer, F.B.; Azhar, S. Thematic review series: Lipid transfer proteins scavenger receptor B type 1: Expression, molecular regulation, and cholesterol transport function. J. Lipid Res. 2018, 59, 1114–1131. [Google Scholar] [CrossRef]
- Cai, L.; Ji, A.; de Beer, F.C.; Tannock, L.R.; van der Westhuyzen, D.R. SR-BI protects against endotoxemia in mice through its roles in glucocorticoid production and hepatic clearance. J. Clin. Investig. 2008, 118, 364–375. [Google Scholar] [CrossRef]
- Han, H.; Wang, L.; Xu, S.; Wang, S.; Yang, M.; Qin, Q.; Wei, S. Identification and characterization of scavenger receptor class B type 1 in orange-spotted grouper, Epinephelus coioides. Aquaculture 2022, 546, 737366. [Google Scholar] [CrossRef]
- Li, C.; Ge, X.; Su, B.; Fu, Q.; Wang, B.; Liu, X.; Ren, Y.; Song, L.; Yang, N. Characterization of class B scavenger receptor type 1 (SRB1) in turbot (Scophthalmus maximus L.). Fish Shellfish Immunol. 2020, 100, 358–367. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, Y.; Zhang, Y.; Ning, Z.; Li, Y.; Zhao, Q.; Lu, H.; Huang, R.; Xia, X.; Feng, Q.; et al. The draft genome of the grass carp (Ctenopharyngodon idellus) provides insights into its evolution and vegetarian adaptation. Nat. Genet. 2015, 47, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef]
- Cheng, L.; Fang, Q.; Shah, S.; Atanasov, I.C.; Zhou, Z.H. Subnanometer-resolution structures of the grass carp reovirus core and virion. J. Mol. Biol. 2008, 382, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Guo, H.; Zhang, J.; Yan, L.; Chen, Q.; Yan, S.; Fang, Q. VP5 autocleavage is required for efficient infection by in vitro-recoated aquareovirus particles. J. Gen. Virol. 2015, 96, 1795–1800. [Google Scholar] [CrossRef]
- Meng, E.C.; Goddard, T.D.; Pettersen, E.F.; Couch, G.S.; Pearson, Z.J.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Tools for structure building and analysis. Protein Sci. 2023, 32, e4792. [Google Scholar] [CrossRef]
- Du, F.; Su, J.; Huang, R.; Liao, L.; Zhu, Z.; Wang, Y. Cloning and preliminary functional studies of the JAM-A gene in grass carp (Ctenopharyngodon idellus). Fish Shellfish Immunol. 2013, 34, 1476–1484. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Xiong, L.; Wang, Y.; He, L.; Huang, R.; Liao, L.; Zhu, Z.; Wang, Y. Different responses in one-year-old and three-year-old grass carp reveal the mechanism of age restriction of GCRV infection. Fish Shellfish Immunol. 2019, 86, 702–712. [Google Scholar] [CrossRef]
- Osada, Y.; Sunatani, T.; Kim, I.-S.; Nakanishi, Y.; Shiratsuchi, A. Signalling pathway involving GULP, MAPK and Rac1 for SR-BI-induced phagocytosis of apoptotic cells. J. Biochem. 2009, 145, 387–394. [Google Scholar] [CrossRef]
- Walker, C.G.; Littlejohn, M.D.; Meier, S.; Roche, J.R.; Mitchell, M.D. DNA methylation is correlated with gene expression during early pregnancy in Bos taurus. Physiol. Genom. 2013, 45, 276–286. [Google Scholar] [CrossRef]
- Kleveland, E.J.; Syvertsen, B.L.; Ruyter, B.; Vegusdal, A.; Jørgensen, S.M.; Gjøen, T. Characterization of scavenger receptor class B, type I in Atlantic salmon (Salmo salar L.). Lipids 2006, 41, 1017–1027. [Google Scholar] [CrossRef]
- Sundvold, H.; Helgeland, H.; Baranski, M.; Omholt, S.W.; Våge, D. Characterisation of a novel paralog of scavenger receptor class B member I (SCARB1) in Atlantic salmon (Salmo salar). BMC Genet. 2011, 12, 52. [Google Scholar] [CrossRef] [PubMed]
- Brundert, M.; Ewert, A.; Heeren, J.; Behrendt, B.; Ramakrishnan, R.; Greten, H.; Merkel, M.; Rinninger, F. Scavenger receptor class B type I mediates the selective uptake of high-density lipoprotein–associated cholesteryl ester by the liver in mice. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Verwilligen, R.A.F.; Mulder, L.; Araújo, P.M.; Carneiro, M.; Bussmann, J.; Hoekstra, M.; Van Eck, M. Zebrafish as outgroup model to study evolution of scavenger receptor class B type I functions. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2023, 1868, 159308. [Google Scholar] [CrossRef]
- Duggan, A.E.; Marie, R.S.; Callard, I.P. Expression of SR-BI (scavenger receptor class B type I) in turtle (Chrysemys picta) tissues and other nonmammalian vertebrates. J. Exp. Zool. 2002, 292, 430–434. [Google Scholar] [CrossRef]
- He, L.; Zhu, D.; Liang, X.; Li, Y.; Liao, L.; Yang, C.; Huang, R.; Zhu, Z.; Wang, Y. Multi-Omics sequencing provides insights into age-dependent susceptibility of grass carp (Ctenopharyngodon idellus) to reovirus. Front. Immunol. 2021, 12, 694965. [Google Scholar] [CrossRef] [PubMed]
- Emmenegger, E.J.; Sanders, G.E.; Conway, C.M.; Binkowski, F.P.; Winton, J.R.; Kurath, G. Experimental infection of six North American fish species with the North Carolina strain of spring viremia of carp virus. Aquaculture 2016, 450, 273–282. [Google Scholar] [CrossRef]
- Embregts, C.W.E.; Rigaudeau, D.; Veselý, T.; Pokorová, D.; Lorenzen, N.; Petit, J.; Houel, A.; Dauber, M.; Schütze, H.; Boudinot, P.; et al. Intramuscular DNA vaccination of juvenile carp against spring viremia of carp virus induces full protection and establishes a virus-specific B and T cell response. Front. Immunol. 2017, 8, 1340. [Google Scholar] [CrossRef]
- Jaramillo, D.; Hick, P.; Whittington, R.J. Age dependency of nervous necrosis virus infection in barramundi Lates Calcarifer (Bloch). J. Fish Dis. 2017, 40, 1089–1101. [Google Scholar] [CrossRef] [PubMed]
- García-Álvarez, M.Á.; Arizcun, M.; Chaves-Pozo, E.; Cuesta, A. Profile of innate immunity in gilthead seabream larvae reflects mortality upon betanodavirus reassortant infection and replication. Int. J. Mol. Sci. 2022, 23, 5092. [Google Scholar] [CrossRef]
- Sun, X.; Bao, N.; Rui, C.; Xue, Y.; Fang, Q.; Zheng, T.; Lin, Z.; Liu, X.; Wang, X. Identification of large yellow croakers (Larimichthys crocea) scavenger receptor genes: Involvement in immune response to Pseudomonas plecoglossicida infection and hypoxia-exposure experiments. Fish Shellfish Immunol. 2024, 144, 109307. [Google Scholar] [CrossRef]
- Ning, X.; Sun, L. Gene network analysis reveals a core set of genes involved in the immune response of Japanese flounder (Paralichthys olivaceus) against vibrio anguillarum infection. Fish Shellfish Immunol. 2020, 98, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Eslamloo, K.; Xue, X.; Hall, J.R.; Smith, N.C.; Caballero-Solares, A.; Parrish, C.C.; Taylor, R.G.; Rise, M.L. Transcriptome profiling of antiviral immune and dietary fatty acid dependent responses of Atlantic salmon macrophage-like cells. BMC Genom. 2017, 18, 706. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, R.; Chen, L.; Li, Y.; Li, Y.; Liao, L.; He, L.; Zhu, Z.; Wang, Y. Characterization of SR-B2a and SR-B2b genes and their ability to promote GCRV infection in grass carp (Ctenopharyngodon idellus). Dev. Comp. Immunol. 2021, 124, 104202. [Google Scholar] [CrossRef]
- Wang, H.; Liu, W.; Yu, F.; Lu, L. Disruption of clathrin-dependent trafficking results in the failure of grass carp reovirus cellular entry. Virol. J. 2016, 13, 25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Guo, H.; Zhang, J.; Chen, Q.; Fang, Q. Identification of the caveolae/raft-mediated endocytosis as the primary entry pathway for aquareovirus. Virology 2018, 513, 195–207. [Google Scholar] [CrossRef]
- Sutherland, D.M.; Strebl, M.; Koehler, M.; Welsh, O.L.; Yu, X.; Hu, L.; Dos Santos Natividade, R.; Knowlton, J.J.; Taylor, G.M.; Moreno, R.A.; et al. NgR1 binding to reovirus reveals an unusual bivalent interaction and a new viral attachment protein. Proc. Natl. Acad. Sci. USA 2023, 120, e2219404120. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′→3′) | Application |
|---|---|---|
| CiSRB1-F1 | GGTTTGGCAGTTTTGTTCGG | Partial sequence obtaining |
| CiSRB1-R1 | GAATGGTTGCGAGTCCGAGA | |
| CiSRB1-5′R-out | GTTCGTTCTTCGGGTTTATCTCTACATTC | 5′-Race PCR amplification |
| CiSRB1-5′R-in | CAAACACCACGACAGTCCCGAAC | |
| CiSRB1-3′F-out | ACAGAAACAGGAAAGATAACAGAGGTG | 3′-Race PCR amplification |
| CiSRB1-3′F-in | ATCTTCATCGGTCTCGGACTCGCA | |
| CiSRB1-F2 | GTGTGAAGTAAGGATGGCGGT | ORF qualifying |
| CiSRB1-R2 | GTGGATATTGGCTCTAGCTCGT | |
| CiSRB1-qF | GGGAGATGAATCCGATGTGGTC | qPCR amplification |
| CiSRB1-qR | GACCTTCAACGAGGGACCTTTC | |
| Ciβ-actin-qF | GGATGATGAAATTGCCGCACTGG | qPCR amplification |
| Ciβ-actin-qR | ACCGACCATGACGCCCTGATGT | |
| HZ08F | AGCGCAGCAGGCAATTACTATCT | qPCR amplification |
| HZ08R | ATCTGCTGGTAATGCGGAACG |
| Primer Name | Sequence (5′→3′) | Length (bp) |
|---|---|---|
| CiSRB1-D-F1 | GTGTGAAGTAAGGATGGCGGT | 1310 bp |
| CiSRB1-D-R1 | CTTTTAGGATTTCGTCAGGATTG | |
| CiSRB1-D-F2 | CACTATGTGGAAGGACATCCC | 1040 bp |
| CiSRB1-D-R2 | GTTTCGCTTCAGAAGACCTCTAT | |
| CiSRB1-D-F3 | GAGAGTCAAAAAATACATATACAGGC | 1223 bp |
| CiSRB1-D-R3 | ATACAGCACTGTTACGCTTTGG | |
| CiSRB1-D-F4 | TCAAAAACGGTCCACGAGC | 1424 bp |
| CiSRB1-D-R4 | CCCTCATGTTTGCATAAGTCTAGAT | |
| CiSRB1-D-F5 | CTGTTCCAAACGTAGCTGCCT | 1432 bp |
| CiSRB1-D-R5 | TTCATTTGGGCTCAATCCGT | |
| CiSRB1-D-F6 | CCATCCACACTTTTTTGCGG | 1384 bp |
| CiSRB1-D-R6 | TTGATCGAATAGGACCAGAGGG | |
| CiSRB1-D-F7 | ATGACATTTCTGCCCCACTATGG | 1321 bp |
| CiSRB1-D-R7 | AAACACCGAAACTAACACGACCC | |
| CiSRB1-D-F8 | CTGTCCCTTTTATAGCATTGG | 1187 bp |
| CiSRB1-D-R8 | TAAACATTTGAAATATATCAGTCTG | |
| CiSRB1-D-F9 | GGTGCTGGTCATATAATTAGAAT | 1216 bp |
| CiSRB1-D-R9 | GACGACCATTGACACCTT | |
| CiSRB1-D-F10 | TCCTTCCCTTCGCCTCTC | 1311 bp |
| CiSRB1-D-R10 | TATTCCTGTGATGCAAAGCTG | |
| CiSRB1-D-F11 | GAGCAGCAAATCAGCATATTAG | 1253 bp |
| CiSRB1-D-R11 | CACCACCTCTGTTATCTTTCCT | |
| CiSRB1-D-F12 | CTGTACAATCTCTCATGGACTCAT | 1252 bp |
| CiSRB1-D-R12 | CAAACGTTACTGCGGCTC | |
| CiSRB1-D-F13 | TAGGATTACTCTTGTTGCCTAGC | 1297 bp |
| CiSRB1-D-R13 | TGAGGTATAATATCACGGCTCC | |
| CiSRB1-D-F14 | CCGTTCTCAACACGTTCCG | 1229 bp |
| CiSRB1-D-R14 | ACATTAGAAGGAACATTGCCCAC | |
| CiSRB1-D-F15 | ACTTTATCTCCTTCACTGATCTGT | 1355 bp |
| CiSRB1-D-R15 | AGTGTTTGGCATAGTAATGGAT | |
| CiSRB1-D-F16 | ACGAAGGTCTTTCAGGTTTGGA | 1538 bp |
| CiSRB1-D-R16 | AGGAAGAAACATCCCCACTCAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Shi, J.; Lu, Y.; Luo, Q.; Chu, P.; Huang, R.; Chen, K.; Zhao, J.; Wang, Y.; Ou, M. Molecular Cloning and Characterization of Scavenger Receptor Class B Type 1 in Grass Carp (Ctenopharyngodon idellus) and Its Expression Profile following Grass Carp Reovirus Challenge. Fishes 2024, 9, 276. https://doi.org/10.3390/fishes9070276
Zhang Y, Shi J, Lu Y, Luo Q, Chu P, Huang R, Chen K, Zhao J, Wang Y, Ou M. Molecular Cloning and Characterization of Scavenger Receptor Class B Type 1 in Grass Carp (Ctenopharyngodon idellus) and Its Expression Profile following Grass Carp Reovirus Challenge. Fishes. 2024; 9(7):276. https://doi.org/10.3390/fishes9070276
Chicago/Turabian StyleZhang, Yang, Jiayuan Shi, Yuntao Lu, Qing Luo, Pengfei Chu, Rong Huang, Kunci Chen, Jian Zhao, Yaping Wang, and Mi Ou. 2024. "Molecular Cloning and Characterization of Scavenger Receptor Class B Type 1 in Grass Carp (Ctenopharyngodon idellus) and Its Expression Profile following Grass Carp Reovirus Challenge" Fishes 9, no. 7: 276. https://doi.org/10.3390/fishes9070276
APA StyleZhang, Y., Shi, J., Lu, Y., Luo, Q., Chu, P., Huang, R., Chen, K., Zhao, J., Wang, Y., & Ou, M. (2024). Molecular Cloning and Characterization of Scavenger Receptor Class B Type 1 in Grass Carp (Ctenopharyngodon idellus) and Its Expression Profile following Grass Carp Reovirus Challenge. Fishes, 9(7), 276. https://doi.org/10.3390/fishes9070276