
Vocalization Pattern and Echolocation Signal Characteristics of Yangtze Finless Porpoise (Neophocaena asiaeorientalis asiaeorientalis) in Captivity

Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Experimental Procedure
2.3. Data Acquisition and Analytical Processing
3. Results
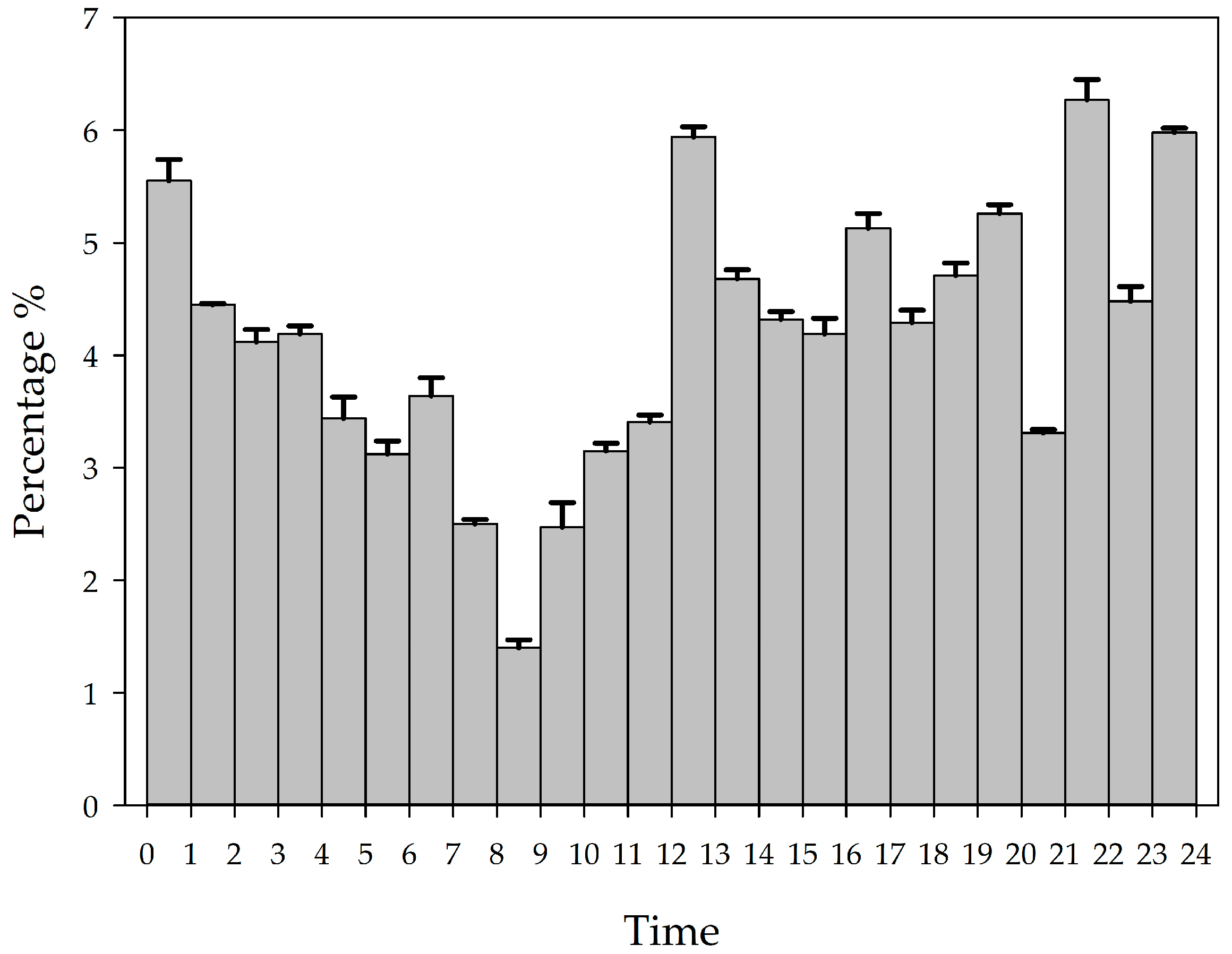
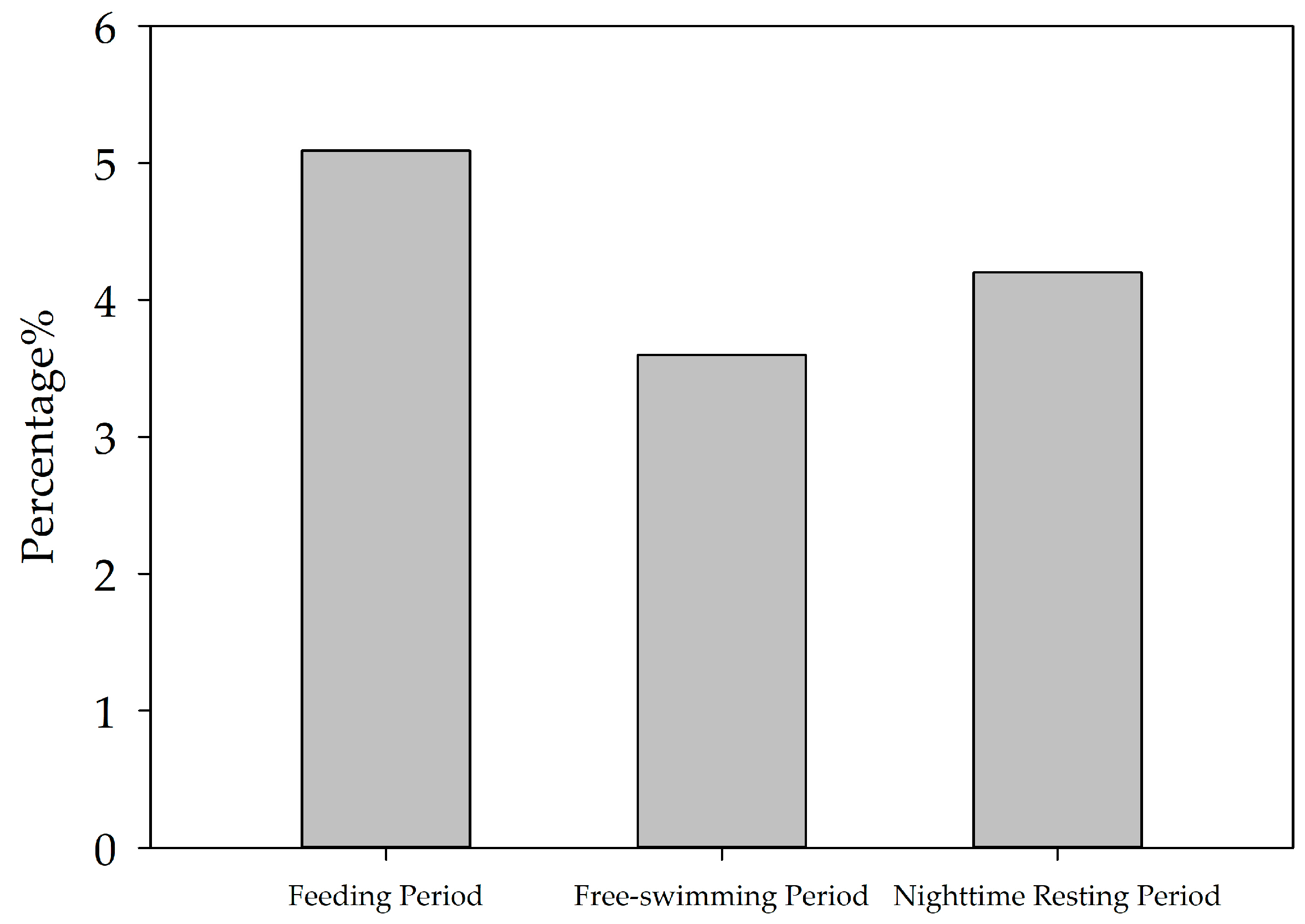
3.1. Comparison of Vocalization Number Characteristics of the YFPs in Three Behavioral States
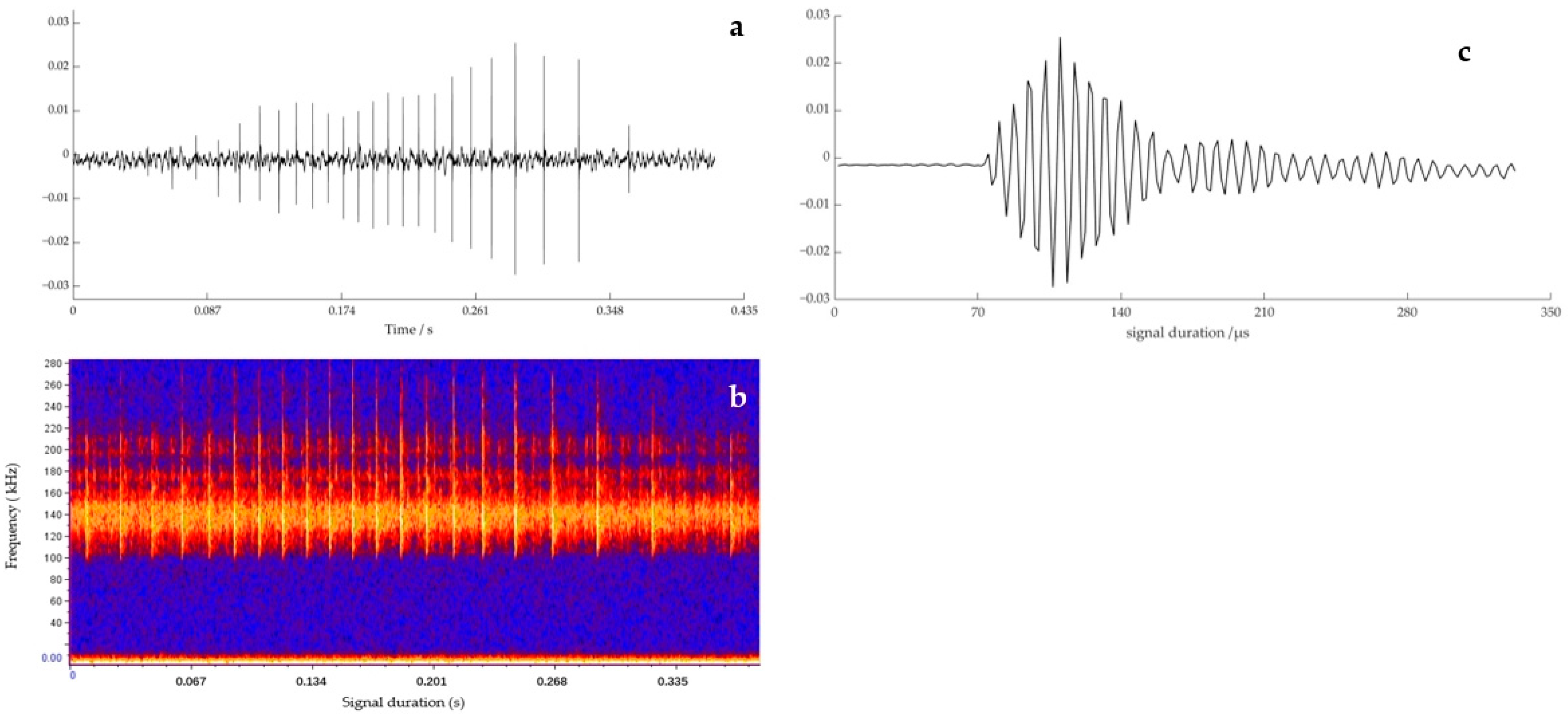
3.2. Comparison of Echolocation Signal Parameters of the YFPs under Three Behavioral States
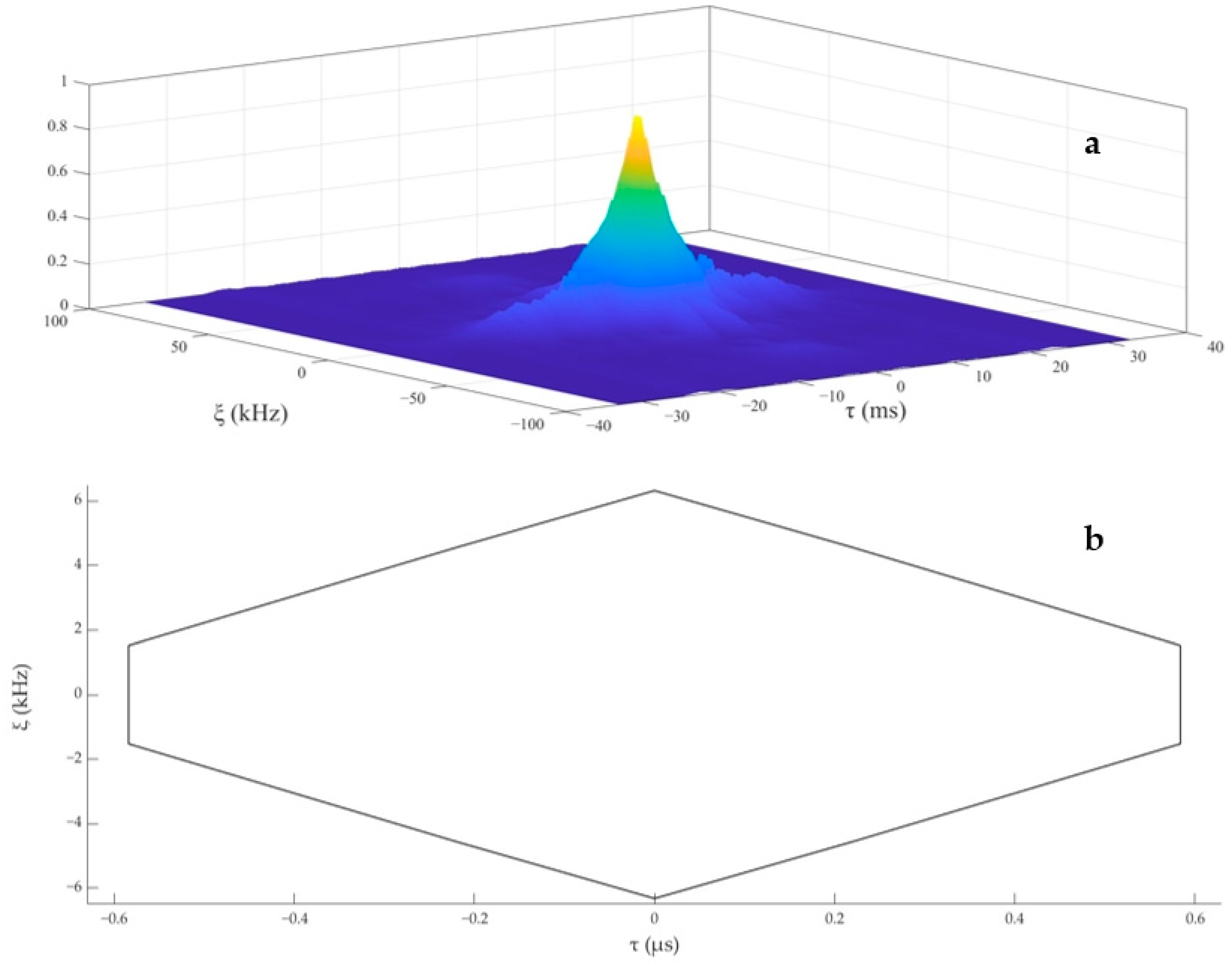
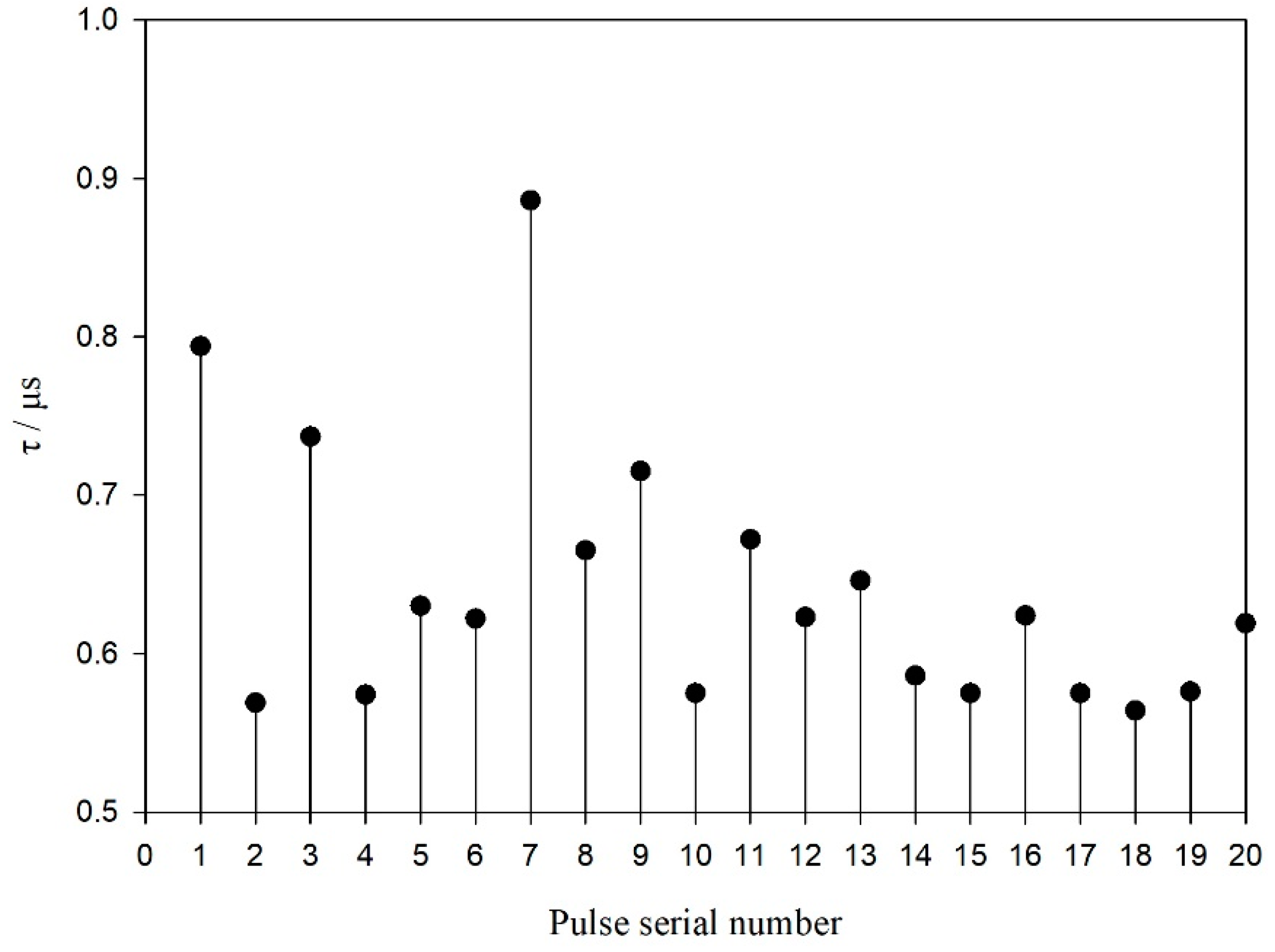
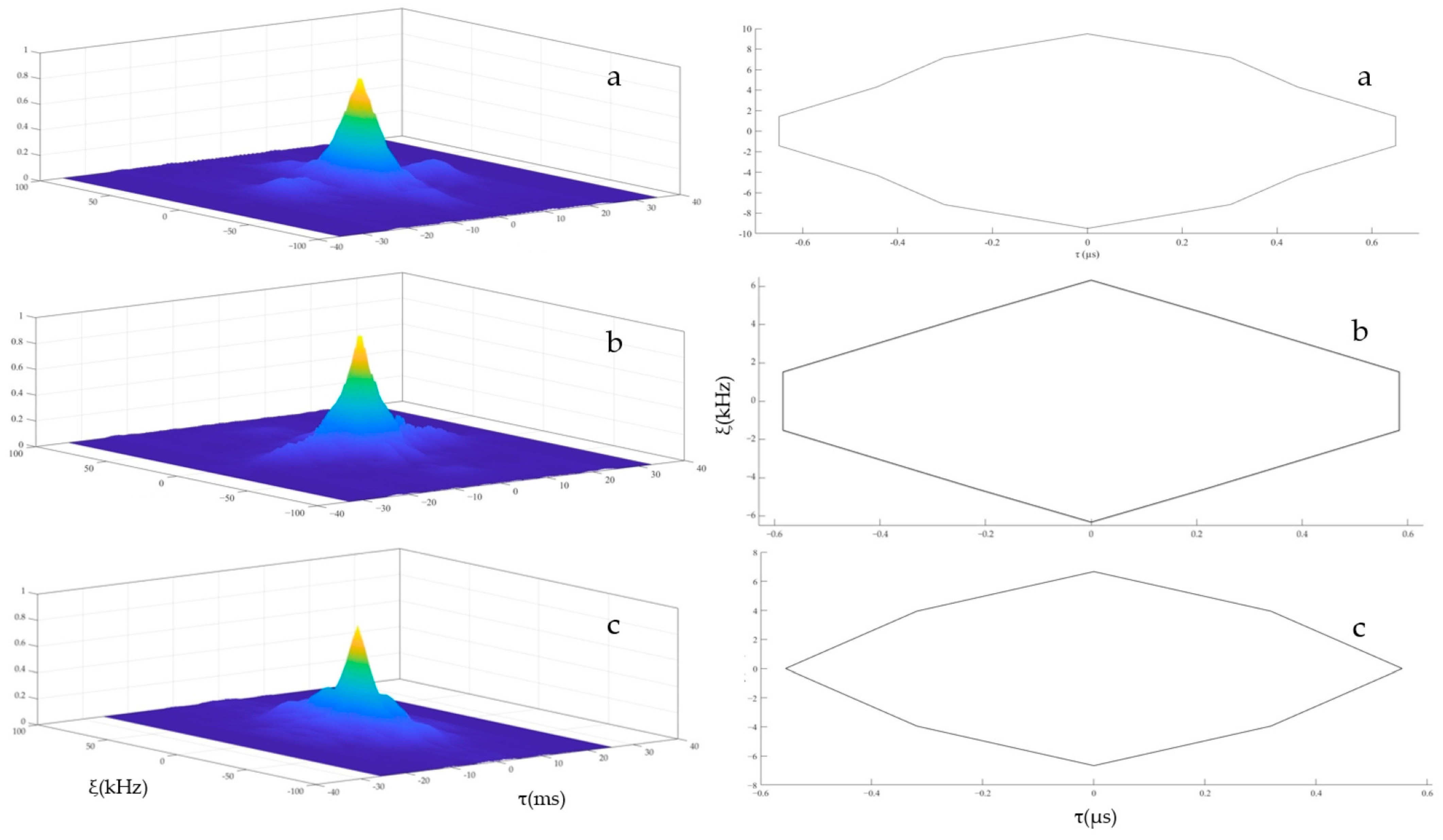
3.3. Echolocation Signal Resolution of the YFPs
4. Discussion
4.1. Analysis of the Diurnal Variation Pattern of the Number of Vocalization Events of the YFPs
4.2. Analysis of Echolocation Signals Characterization of the YFPs in Three Behavioral States
4.3. Analysis of Echolocation Signal Resolution of the YFPs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, X.; Guang, X.; Sun, D.; Xu, S.; Yang, G. Population genomics of finless porpoises reveal an incipient cetacean species adapted to freshwater. Nat. Commun. 2018, 9, 1276. [Google Scholar] [CrossRef]
- Wang, D.; Turvey, S.; Zhao, X.; Mei, Z. Neophocaena asiaeorientalis ssp. asiaeorientalis. The IUCN Red List of Threatened Species; Version 3.1; IUCN: Gland, Switzerland; Cambridge, UK, 2013. [Google Scholar]
- Yan, F.J.; Yu, W.J. The Yangtze finless porpoise population has rebounded to 1249. Xinhua Daily Telegraph, 1 March 2023; p. 006. [Google Scholar]
- Wang, K.X.; Wang, D. Characteristics and Functions of Sound of the Yangtze Finless Porpolse (Neophocaena phocaenoides) in Captivity. Tech. Acoust. 1999, 18, 4. [Google Scholar]
- Wang, D. A preliminary study on sound and acoustic behavior of the Yangtze Finless Porpoise, Neophocaena Phocaenoides. Acta Hydrobiol. Sin. 1996, 20, 127–133. [Google Scholar]
- Busnel, R. (Ed.) Animal Sonar Systems; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; Volume 28. [Google Scholar]
- Zhang, T.C.; Ju, T.; Li, S.H.; Xie, Y.; Wang, D.; Wang, Z.T.; Wang, K.X. Navigation noise property of large vessels in Hechangzhou region of the Yangtze River and its potential effects on the Yangtze finless porpoise. Acta Theriol. Sin. 2018, 38, 543–550. [Google Scholar]
- Duan, P.X.; Wang, Z.T.; Akamatsu, T.; Tregenza, N.; Li, G.Y.; Wang, K.X.; Wang, D. Anthropogenic activity, hydrological regime, and light level jointly influence temporal patterns in biosonar activity of the Yangtze finless porpoise at the junction of the Yangtze River and Poyang Lake, China. Zool. Res. 2023, 44, 919–931. [Google Scholar] [CrossRef]
- Michael, L.; Peter, T.M. Context-dependent biosonar adjustments during active target approaches in echolocating harbour porpoises. J. Exp. Biol. 2019, 222, jeb206169. [Google Scholar]
- Fang, L.; Wang, D.; Li, Y.; Cheng, Z.; Pine, M.K.; Wang, K.X.; Li, S.H. The source parameters of echolocation clicks from captive and free-ranging Yangtze finless porpoises (Neophocaena asiaeorientalis asiaeorientalis). PLoS ONE 2015, 10, e0129143. [Google Scholar] [CrossRef]
- Wang, Z.; Akamatsu, T.; Wang, K.; Wang, D. The diel rhythms of biosonar behavior in the Yangtze Finless Porpoise (Neophocaena asiaeorientalis asiaeorientalis) in the port of the Yangtze River: The correlation between prey availability and boat traffic. PLoS ONE 2014, 9, e97907. [Google Scholar] [CrossRef]
- Chen, M.; Yu, D.; Wang, K.; Zhang, K.; Wang, Z. Seasonal and diel activities of the Yangtze finless porpoise in natural and highly disturbed habitats: Implications for conservation planning of freshwater cetaceans. Aquat. Conserv. Mar. Freshw. Ecosyst. 2022, 32, 605–616. [Google Scholar] [CrossRef]
- Serres, A.; Xu, C.; Hao, Y.G.; Wang, D. The click production of captive Yangtze Finless Porpoises (Neophocaena asiaeorientalis asiaorientalis) is influenced by social and environmental factors. Animals 2021, 11, 511. [Google Scholar] [CrossRef]
- Chloe, E.M.; Laia, R.-D.; Peter, T.M. Directional biosonar beams allow echolocating harbour porpoises to actively discriminate and intercept closely-spaced targets. J. Exp. Biol. 2021, 224, jeb242779. [Google Scholar]
- Thorpe, C.W.; Bates, R.H.; Dawson, S.M. Intrinsic echolocation capability of Hector’s dolphin, Cephalorhynchus hectori. J. Acoust. Soc. Am. 1991, 90, 2931–2934. [Google Scholar] [CrossRef]
- Akamatsu, T.; Wang, D.; Wang, K.; Li, S.; Dong, S.; Zhao, X.; Barlow, J.; Stewart, B.S.; Richlen, M. Estimation of the detection probability for Yangtze finless porpoises (Neophocaena phocaenoides asiaeorientalis) with a passive acoustic method. J. Acoust. Soc. Am. 2008, 123, 4403–4411. [Google Scholar] [CrossRef]
- Li, S.H.; Wang, K.X.; Wang, D.; Akamatsu, T. Echolocation signals of the free-ranging Yangtze finless porpoise (Neophocaena phocaenoides asiaeorientialis). J. Acoust. Soc. Am. 2005, 117, 3288–3296. [Google Scholar] [CrossRef]
- Li, S.H.; Wang, D.; Wang, K.X.; Taylor, E.A.; Cros, E.; Shi, W.J.; Wang, Z.T.; Fang, L.; Chen, Y.F.; Kong, F.M. Evoked-potential audiogram of an Indo-Pacific humpback dolphin (Sousa chinensis). J. Exp. Biol. 2012, 215 Pt 17, 3055–3063. [Google Scholar] [CrossRef]
- Li, S.; Akamatsu, T.; Dong, L.; Wang, K.; Wang, D.; Kimura, S. Widespread passive acoustic detection of Yangtze finless porpoise using miniature stereo acoustic data-loggers: A review. J. Acoust. Soc. Am. 2010, 128, 1476–1482. [Google Scholar] [CrossRef]
- Wang, Z.; Akamatsu, T.; Mei, Z.; Dong, L.; Imaizumi, T.; Wang, K.; Wang, D. Frequent and prolonged nocturnal occupation of port areas by Yangtze finless porpoises (Neophocaena asiaeorientalis): Forced choice for feeding? Integr. Zool. 2015, 10, 122–132. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, P.; Xia, D.J.; Li, Z.H.; Wu, Z.; Yu, D.P.; Chen, M.M. Study on the utilization pattern of island-type habitat patches by Yangtze finless porpoise and analysis of potential factors. Acta Hydrobiol. Sin. 2023, 48, 1–10. [Google Scholar]
- Sasaki-Yamamoto, Y.; Akamatsu, T.; Ura, T.; Sugimatsu, H.; Kojima, J.; Bahl, R.; Behera, S.; Kohshima, S. Diel changes in the movement patterns of Ganges River dolphins monitored using stationed stereo acoustic data loggers. Mar. Mammal Sci. 2013, 29, 589–605. [Google Scholar] [CrossRef]
- Akamatsu, T.; Wang, D.; Wang, K.X.; Naito, Y. Biosonar behaviour of free-ranging porpoises. Proc. R. Soc. B 2005, 272, 797–801. [Google Scholar] [CrossRef]
- Akamatsu, T.; Teilmann, J.; Miller, L.A.; Tougaard, J.; Dietz, R.; Wang, D.; Siebert, U.; Naito, Y. Comparison of echolocation behaviour between coastal and riverine porpoises. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2007, 54, 290–297. [Google Scholar] [CrossRef]
- Au, W.W.; Würsig, B. Echolocation signals of dusky dolphins (Lagenorhynchus obscurus) in Kaikoura, New Zealand. J. Acoust. Soc. Am. 2004, 115, 2307–2313. [Google Scholar] [CrossRef]
- Fang, L.; Li, S.; Wang, K.; Wang, Z.; Shi, W.; Wang, D. Echolocation signals of free-ranging Indo-Pacific humpback dolphins (Sousa chinensis) in Sanniang Bay, China. J. Acoust. Soc. Am. 2015, 138, 1346–1352. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Wang, X.Y.; XU, X.H.; Song, Z.Z.; Yang, W.Y.; Zhang, Y. Analysis of echolocation from Indo-Pacific humpback dolphin (Sousa chinensis) and finless porpoise (Neophocaena asiaeorientalis sunmeri). Acta Acust. 2021, 46, 423–430. [Google Scholar]
- Madsen, P.T.; Carder, D.A.; Bedholm, K.; Ridgway, S.H. Porpoise clicks from a sperm whale nose—Convergent evolution of 130 kHz pulses in toothed whale sonars? Bioacoustics 2005, 15, 195–206. [Google Scholar] [CrossRef]
- Madsen, P.T.; Wahlberg, M. Recording and quantification of ultrasonic echolocation clicks from free-ranging toothed whales. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2007, 54, 1421–1444. [Google Scholar] [CrossRef]
- Li, H.; Gao, D.Z.; Lin, J.H.; Chi, J. Analyses of click signal characteristics of captive bottlenose dolphins, beluga whales and false killer whales. Tech. Acoust. 2019, 38, 24–31. [Google Scholar]
- Chen, S.; Niu, F.Q.; Lin, C.L.; Liu, W. Click signal of bottlenose dolphin and its simulation analysis under different environments. Tech. Acoust. 2019, 38, 7. [Google Scholar]
- Tian, T. Sonar Technology, 2nd ed.; Harbin Engineering University Press: Harbin, China, 2010; pp. 22–30. [Google Scholar]
- Fish, M.P.; Mowbray, W.H. Production of underwater sound by the white whale or beluga, Delphinapterus leucas (Pallas). J. Mar. Res. 1962, 20, 149–162. [Google Scholar]
- Niu, F.Q.; Yang, Y.M.; Wen, H.T.; Liu, Z.W. Vocalization and signal characteristic of bottlenose dolphin. Tech. Acoust. 2011, 30, 148–152. [Google Scholar]
- Lyamin, O.I.; Manger, P.R.; Ridgway, S.H.; Mukhametov, L.M.; Siegel, J.M. Cetacean sleep: An unusual form of mammalian sleep. Neurosci. Biobehav. Rev. 2008, 32, 1451–1484. [Google Scholar] [CrossRef]
- Kremers, D.; Jaramillo, M.B.; Böye, M.; Lemasson, A.; Hausberger, M. Nocturnal vocal activity in captive bottlenose dolphins (Tursiops truncatus): Could dolphins have presleep choruses? Anim. Behav. Cogn. 2014, 1, 464–469. [Google Scholar] [CrossRef][Green Version]
- Therrien, S.C.; Thomas, J.A.; Therrien, R.E.; Stacey, R. Time of Day and Social Change Affect Underwater Sound Production by Bottlenose Dolphins (Tursiops truncatus) at the Brookfield Zoo. Aquat. Mamm. 2012, 38, 65–75. [Google Scholar] [CrossRef]
- Serres, A.; Hao, Y.G.; Wang, D. Swimming features in captive odontocetes: Indicative of animals’ emotional state? Behav. Process. 2020, 170, 103998. [Google Scholar] [CrossRef]
- Wei, Z.; Wang, D.; Zhang, X.F.; Wang, K.X.; Chen, D.Q.; Zhao, Q.Z.; Wang, X. Observation on some sexual behavior of the Yangtze finless porpoise (Neophocaena phocaenoides asiaerientals) in captivity. Acta Theriol. Sin. 2004, 24, 98–102. [Google Scholar]
- Morisaka, T.; Karczmarski, L.; Akamatsu, T.; Sakai, M.; Dawson, S.; Thornton, M. Echolocation signals of Heaviside’s dolphins (Cephalorhynchus heavisidii). J. Acoust. Soc. Am. 2011, 129, 449–457. [Google Scholar] [CrossRef]
- Baumann-Pickering, S.; Roch, M.A.; Wiggins, S.M.; Schnitzler, H.U.; Hildebrand, J.A. Acoustic behavior of melon-headed whales varies on a diel cycle. Behav. Ecol. Sociobiol. 2015, 69, 1553–1563. [Google Scholar] [CrossRef]
- Wei, C.; Xu, X.M.; Zhang, Y.; Niu, F.Q. Comparison study on whistle characteristics of captive bottlenose dolphins (Tursiops turncatus) during swimming and under training. Acta Acust. 2014, 39, 452–458. (In Chinese) [Google Scholar]
- Qing, X. Research on the Mechanisms of Biosonar Operation in the Representative Hunting Behaviour of Cetaceans; Harbin Engineering University: Harbin, China, 2021. [Google Scholar]
- Read, A.J.; Drinker, P.; Northridge, S. Bycatch of marine mammals in U.S. and global fisheries. Conserv. Biol. 2006, 20, 163–169. [Google Scholar] [CrossRef]
- Kastelein, R.A.; Au, W.W.L.; de Haan, D. Detection distances of bottom-set gillnets by harbour porpoises (Phocoena phocoena) and bottlenose dolphins (Tursiops truncatus). Mar. Environ. Res. 2000, 49, 359–375. [Google Scholar] [CrossRef]
- Dukas, R.; Kamil, A.C. The cost of limited attention in blue jays. Behav. Ecol. 2000, 11, 502–506. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Period | |
|---|---|
| Free-swimming State | 6:00–7:00; 7:00–8:00; 9:00–10:00; 11:00–12:00; 13:00–14:00; 15:00–16:00; 17:00–18:00 |
| Feeding State | 10:00–11:00; 12:00–13:00; 14:00–15:00; 6:00–17:00; 19:00–20:00; 21:00–22:00; 0:00–1:00 |
| Nighttime Resting State | 1:00–2:00; 2:00–3:00; 3:00–4:00; 4:00–5:00; 5:00–6:00; 18:00–19:00; 20:00–21:00; 22:00–23:00; 23:00–24:00 |
| p Value | |
|---|---|
| Peak frequency (kHz) | 0.171 |
| −3 dB_BW (kHz) | <0.001 *** |
| −10 dB_BW (kHz) | <0.001 *** |
| −10 dB duration (µs) | <0.001 *** |
| Centroid frequency (kHz) | 0.059 |
| RMS_BW (kHz) | 0.017 * |
| State (I) | State (J) | Mean ± S.D. (I) | Mean ± S.D. (J) | p Value | |
|---|---|---|---|---|---|
| Peak frequency (kHz) | Free-swimming | Feeding | 130.59 ± 5.25 | 129.77 ± 5.66 | 0.064 |
| Free-swimming | Nighttime Resting | 130.59 ± 5.25 | 130.05 ± 5.54 | 0.249 | |
| Feeding | Nighttime Resting | 129.77 ± 5.66 | 130.05 ± 5.54 | 0.540 | |
| −3 dB_BW (kHz) | Free-swimming | Feeding | 19.78 ± 5.60 | 19.14 ± 5.57 | 0.011 * |
| Free-swimming | Nighttime Resting | 19.78 ± 5.60 | 22.04 ± 6.02 | <0.001 *** | |
| Feeding | Nighttime Resting | 19.14 ± 5.57 | 22.04 ± 6.02 | <0.001 *** | |
| −10 dB_BW (kHz) | Free-swimming | Feeding | 34.57 ± 8.30 | 33.79 ± 7.90 | 0.204 |
| Free-swimming | Nighttime Resting | 34.57 ± 8.30 | 37.80 ± 10.12 | <0.001 *** | |
| Feeding | Nighttime Resting | 33.79 ± 7.90 | 37.80 ± 10.12 | <0.001 *** | |
| −10 dB duration (µs) | Free-swimming | Feeding | 54.52 ± 18.88 | 59.20 ± 18.70 | 0.001 ** |
| Free-swimming | Nighttime Resting | 54.52 ± 18.88 | 49.11 ± 15.94 | <0.001 *** | |
| Feeding | Nighttime Resting | 59.20 ± 18.70 | 49.11 ± 15.94 | <0.001 *** | |
| Centroid frequency (kHz) | Free-swimming | Feeding | 122.56 ± 10.75 | 124.14 ± 10.12 | 0.094 |
| Free-swimming | Nighttime Resting | 122.56 ± 10.75 | 124.24 ± 9.64 | 0.132 | |
| Feeding | Nighttime Resting | 124.14 ± 10.12 | 124.24 ± 9.64 | 0.820 | |
| RMS_BW (kHz) | Free-swimming | Feeding | 28.90 ± 15.35 | 26.14 ± 14.00 | 0.035 * |
| Free-swimming | Nighttime Resting | 28.90 ± 15.35 | 25.89 ± 13.42 | 0.037 * | |
| Feeding | Nighttime Resting | 26.14 ± 14.00 | 25.89 ± 13.42 | 0.920 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Liang, H.; Lin, D.; Zhang, J.; Li, D.; Ye, K.; Lu, W.; Liu, K. Vocalization Pattern and Echolocation Signal Characteristics of Yangtze Finless Porpoise (Neophocaena asiaeorientalis asiaeorientalis) in Captivity. Fishes 2024, 9, 119. https://doi.org/10.3390/fishes9040119
Chen J, Liang H, Lin D, Zhang J, Li D, Ye K, Lu W, Liu K. Vocalization Pattern and Echolocation Signal Characteristics of Yangtze Finless Porpoise (Neophocaena asiaeorientalis asiaeorientalis) in Captivity. Fishes. 2024; 9(4):119. https://doi.org/10.3390/fishes9040119
Chicago/Turabian StyleChen, Jia, Haiying Liang, Danqing Lin, Jialu Zhang, Dong Li, Kun Ye, Wenfei Lu, and Kai Liu. 2024. "Vocalization Pattern and Echolocation Signal Characteristics of Yangtze Finless Porpoise (Neophocaena asiaeorientalis asiaeorientalis) in Captivity" Fishes 9, no. 4: 119. https://doi.org/10.3390/fishes9040119
APA StyleChen, J., Liang, H., Lin, D., Zhang, J., Li, D., Ye, K., Lu, W., & Liu, K. (2024). Vocalization Pattern and Echolocation Signal Characteristics of Yangtze Finless Porpoise (Neophocaena asiaeorientalis asiaeorientalis) in Captivity. Fishes, 9(4), 119. https://doi.org/10.3390/fishes9040119