
Amino Acid Variation at the Mitochondrial Binding Site of Antimycin A is Proposed to Reflect Sensitivity and Toxicity Differences among Fish Species


Abstract
1. Introduction

2. Materials and Methods
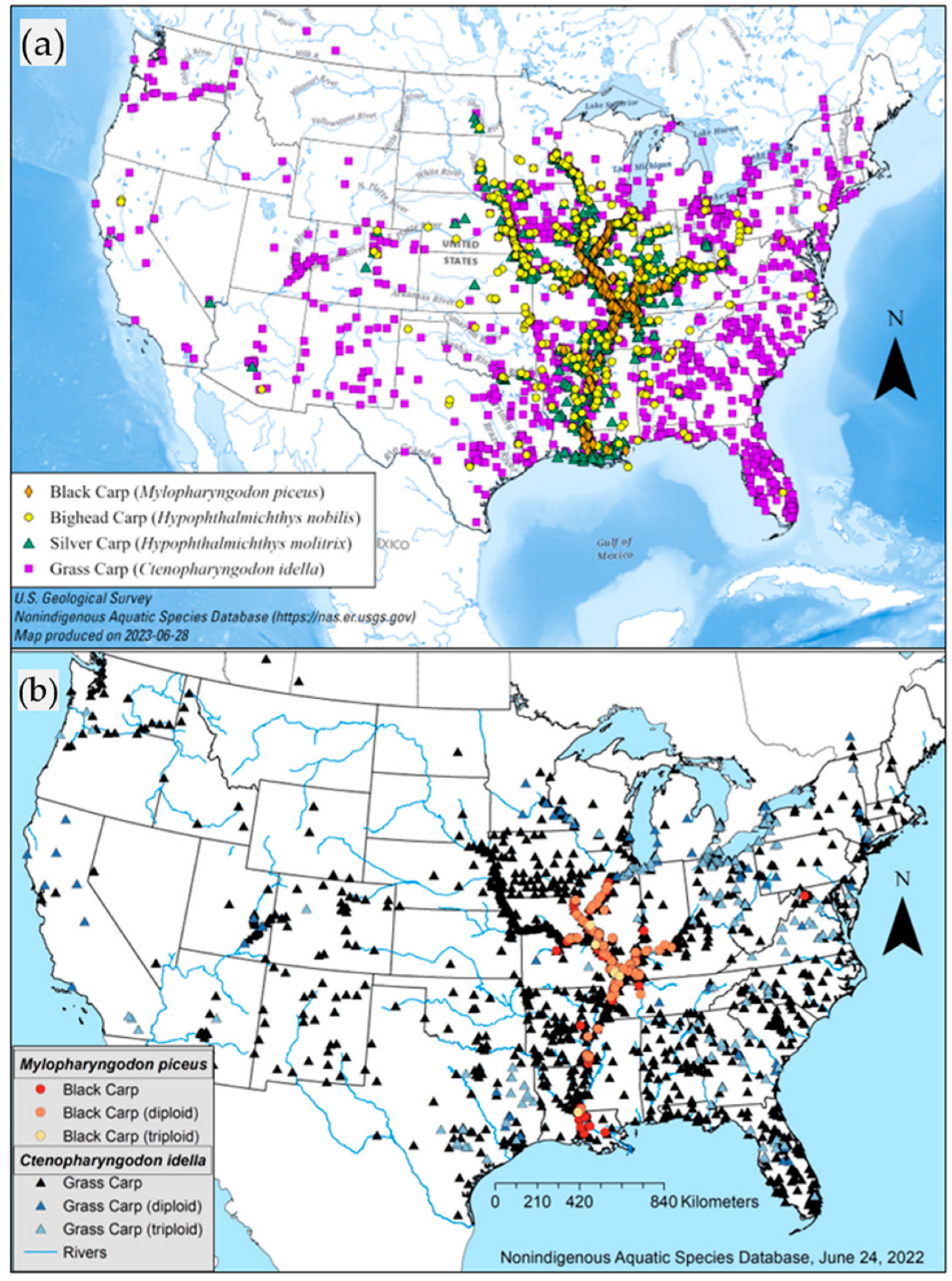
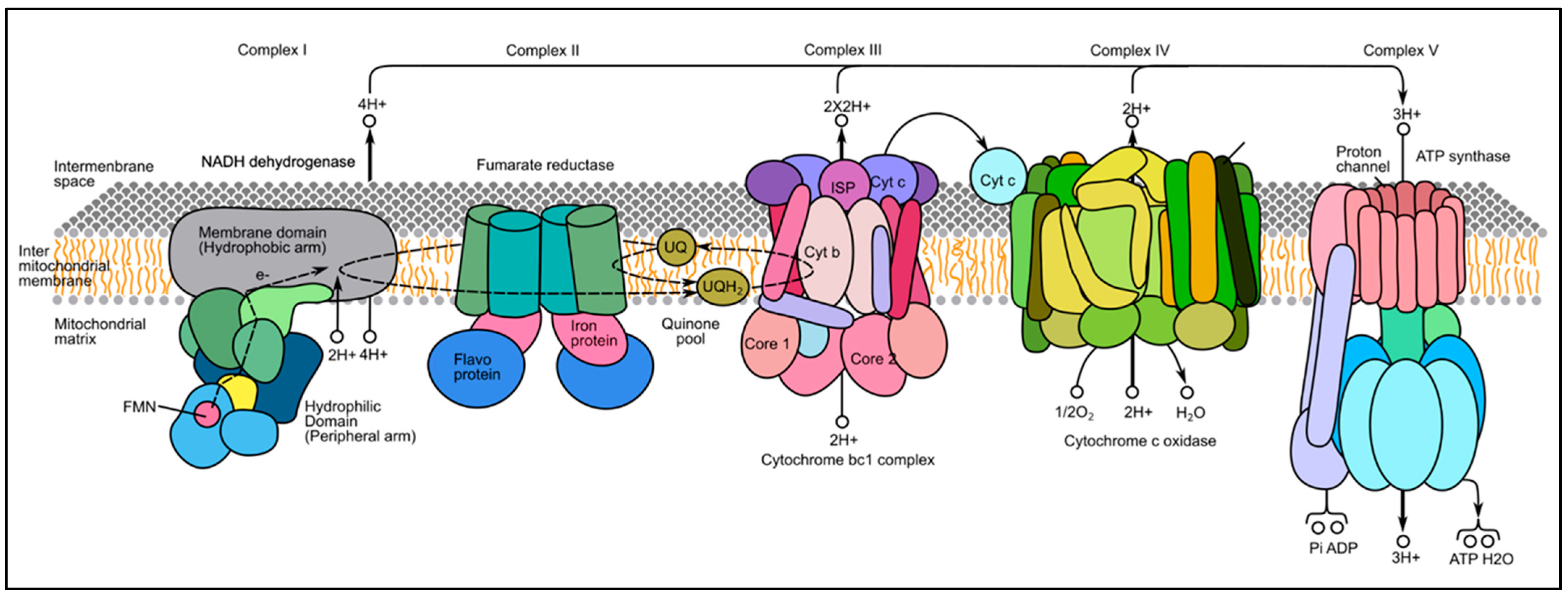
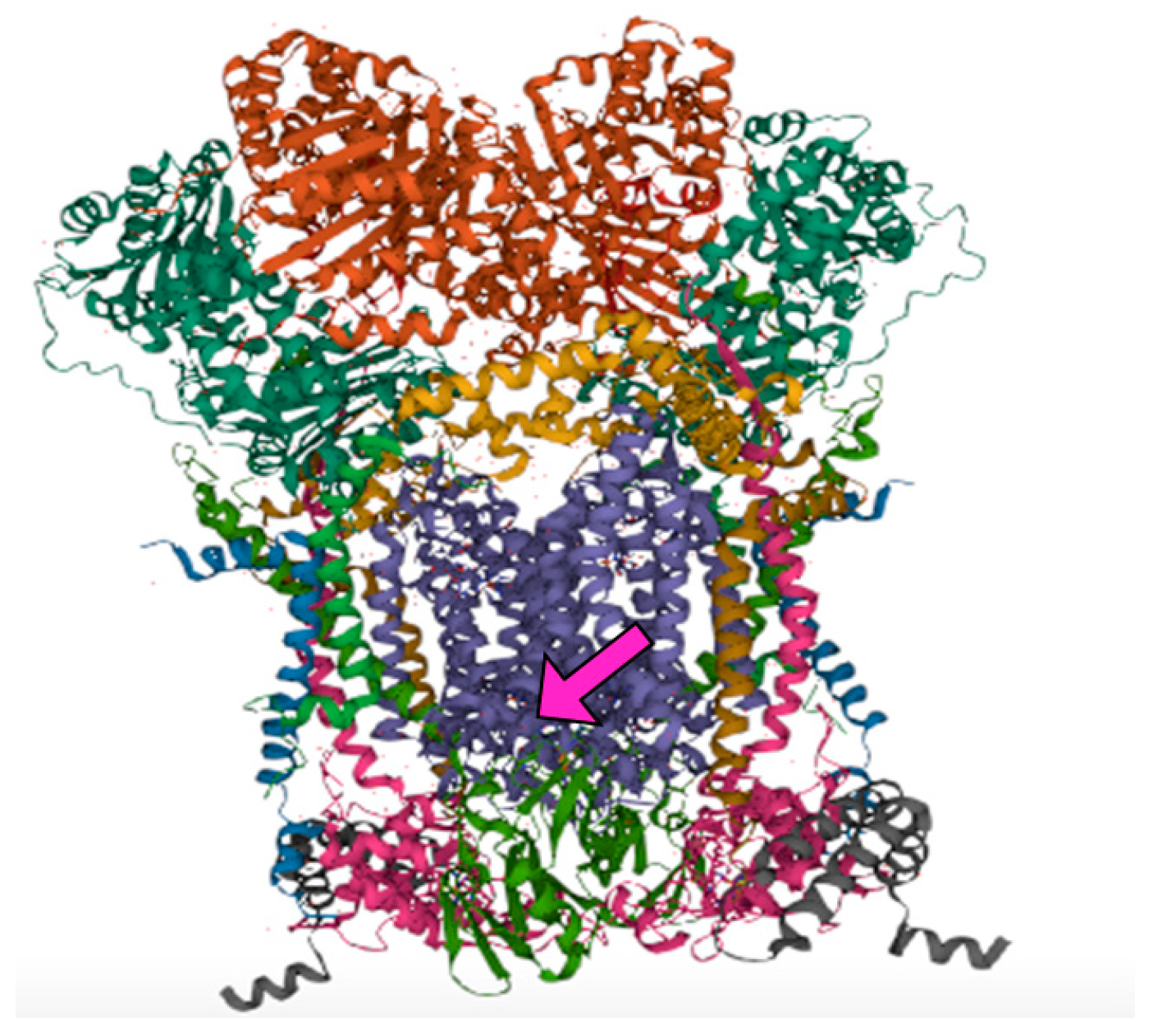
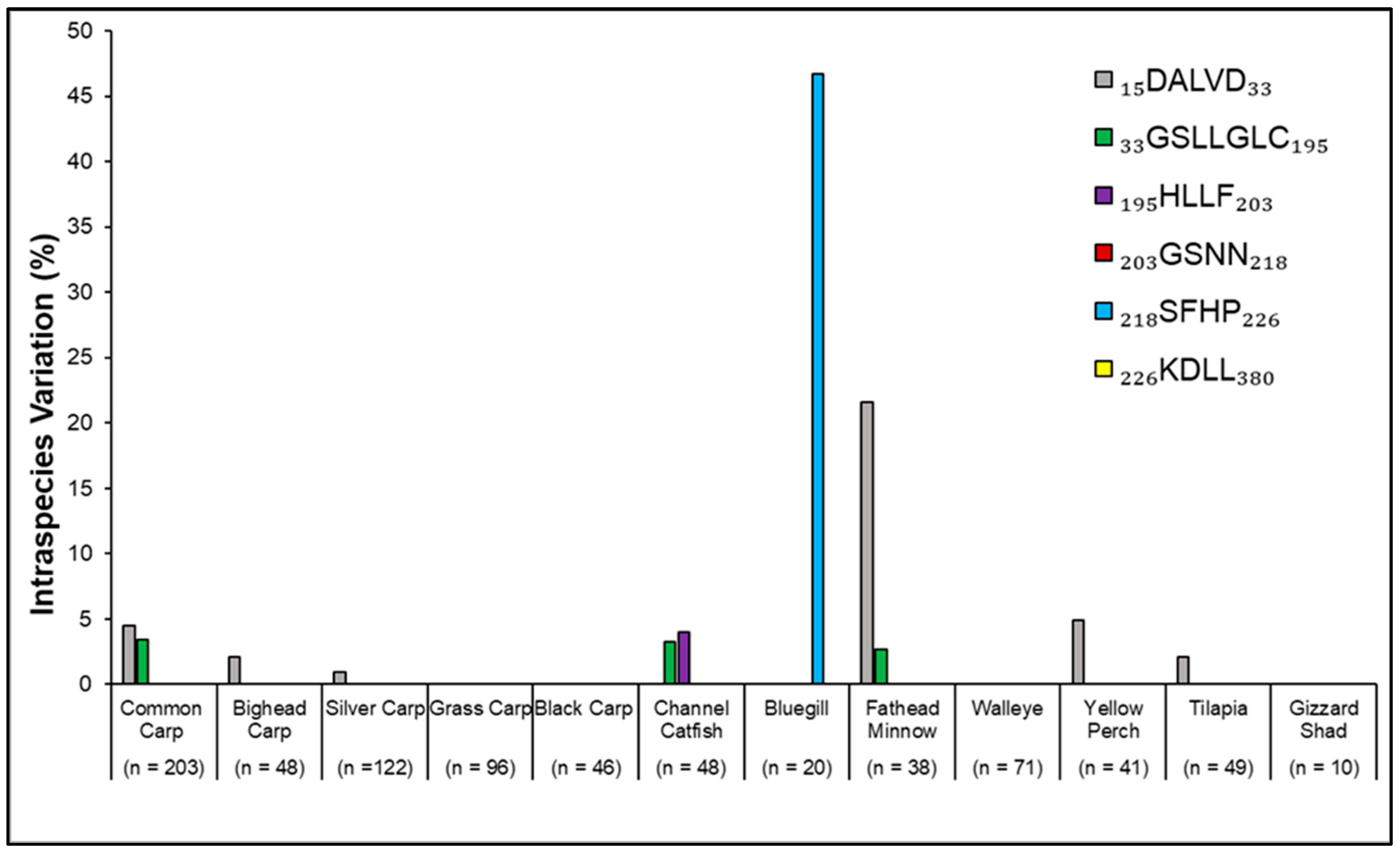
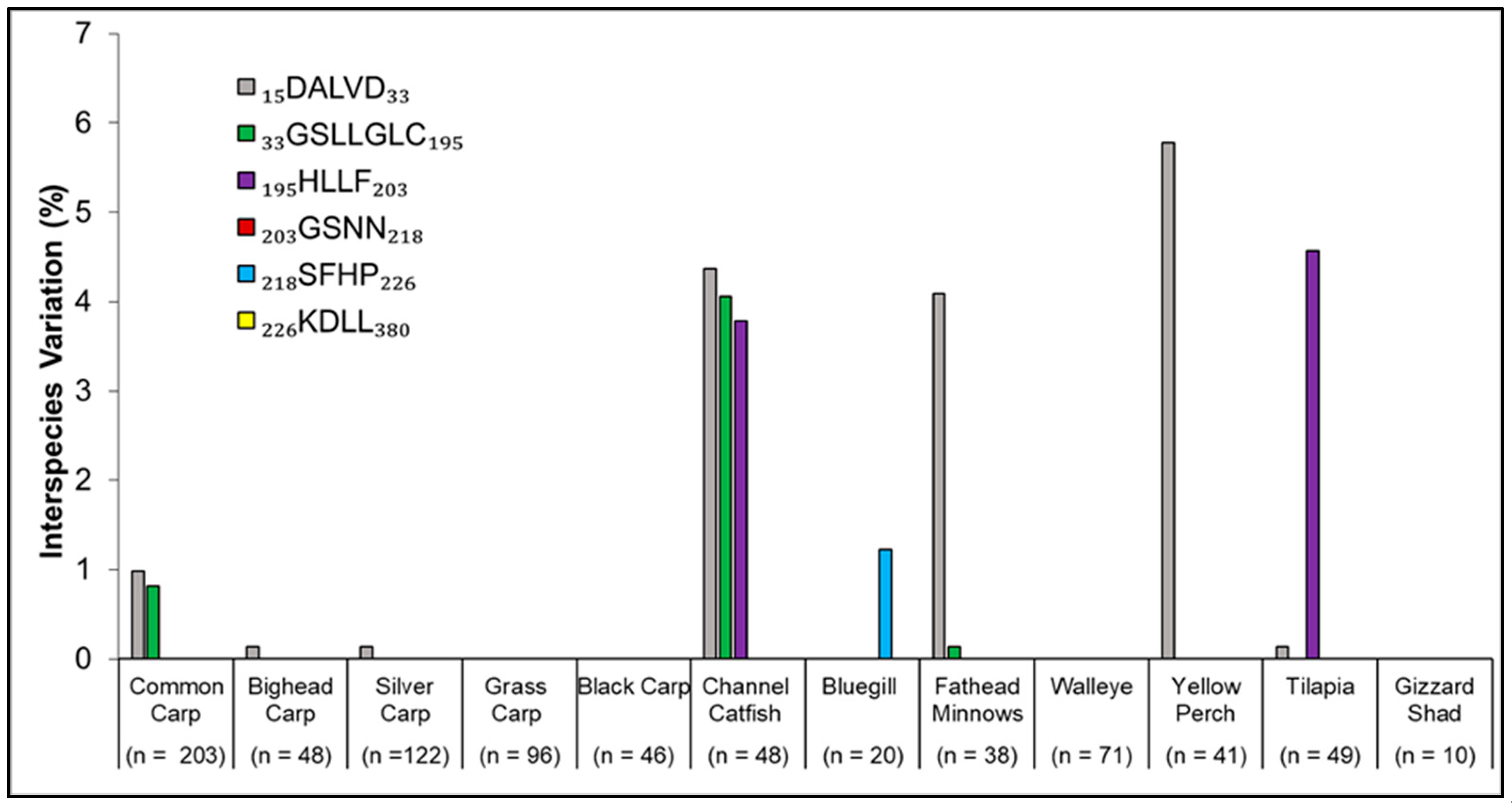
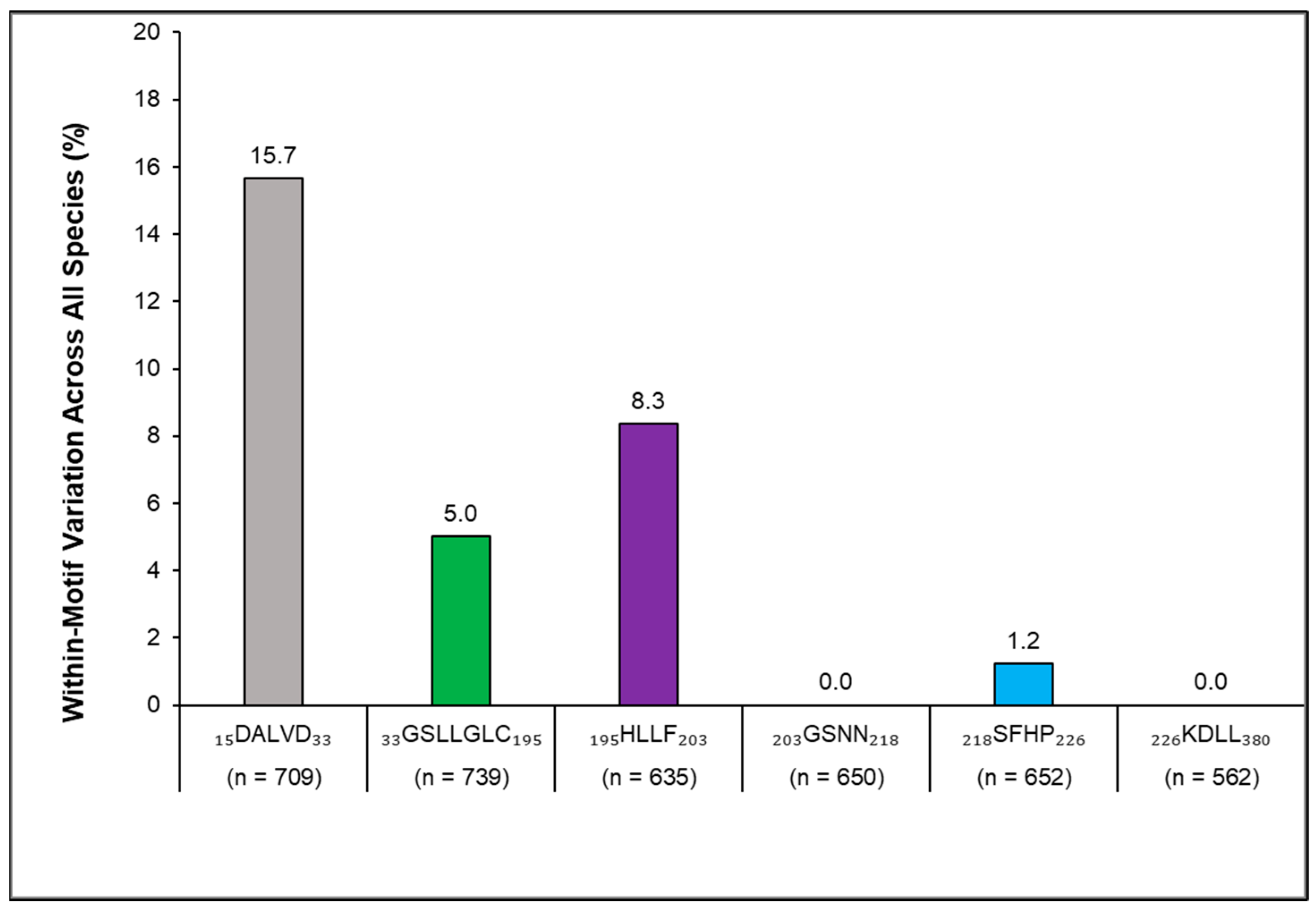
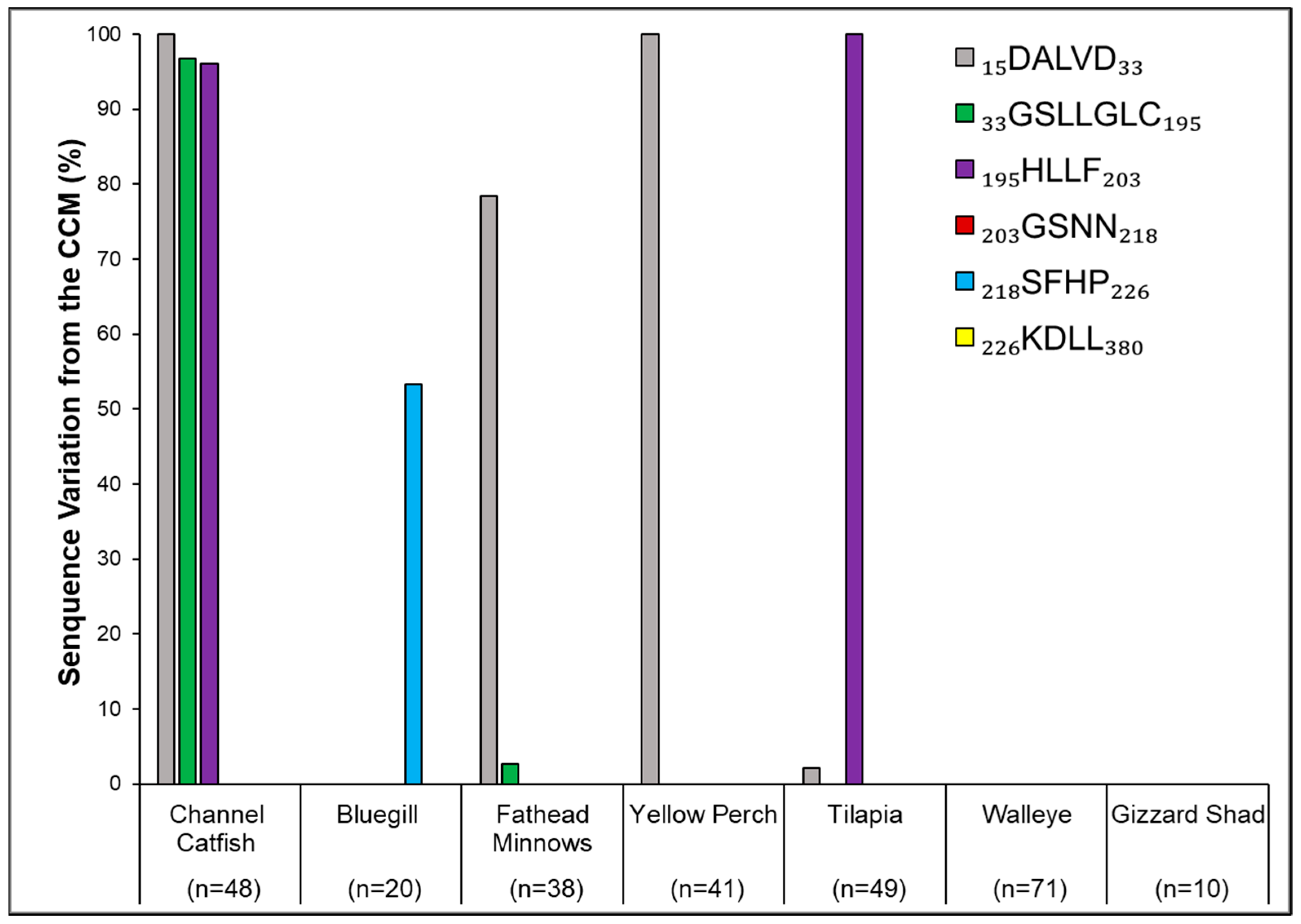
3. Results



4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fish | Amino Acid Position | ||||||
|---|---|---|---|---|---|---|---|
| Common Name | Scientific Name | 16 | 19 | 38 | 197 | 198 | 219 |
| Fathead Minnow | Pimephales promelas | Gly | Val | Gly | Leu | Leu | Ser |
| Bluegill | Lepomis macrochirus | Asp | Val | Gly | Leu | Leu | Leu |
| Yellow Perch | Perca flavescens | Asn | Val | Gly | Leu | Leu | Ser |
| Nile Tilapia | Oreochromis niloticus | Asp | Val | Gly | Leu | Ile | Ser |
| Channel Catfish | Ictalurus punctatus | Asn | Ile | Leu | Ala | Leu | Ser |
References
- Wittmann, M.E.; Jerde, C.L.; Howeth, J.G.; Maher, S.P.; Deines, A.M.; Jenkins, J.A.; Whitledge, G.W.; Burbank, S.R.; Chadderton, W.L.; Mahon, A.R. Grass carp in the Great Lakes region: Establishment potential, expert perceptions, and re-evaluation of experimental evidence of ecological impact. Can. J. Fish. Aquat. Sci. 2014, 71, 992–999. [Google Scholar] [CrossRef]
- Poole, J.R.; Sauey, B.W.; Amberg, J.J.; Bajer, P.G. Assessing the efficacy of corn-based bait containing antimycin-a to control common carp populations using laboratory and pond experiments. Biol. Invasions 2018, 20, 1809–1820. [Google Scholar] [CrossRef]
- Kolar, C.S.; Chapman, D.C.; Courtenay, W.R.; Housel, C.M.; Williams, J.D.; Jennings, D.P. Bigheaded Carps: A biological Synopsis and Environmental Risk Assessment; American Fisheries Society: Bethesda, MD, USA, 2007; 204p. [Google Scholar]
- Conover, G.; Simmonds, R.; Whalen, M. (Eds.) Management and Control Plan for Bighead, Black, Grass, and Silver Carps in the United States; Asian Carp Working Group, Aquatic Nuisance Species Task Force: Washington, DC, USA, 2007; 223p. [Google Scholar]
- Cudmore, B.; Jones, L.A.; Mandrak, N.E.; Dettmers, J.M.; Chapman, D.C.; Kolar, C.S.; Conover, G. Ecological Risk Assessment of Grass Carp (Ctenopharyngodon idella) for the Great Lakes Basin. Fisheries and Oceans Canada, Canadian Science Advisory Secretariat: Ottawa, ON, Canada, 2019; 115p. [Google Scholar]
- Conner, J.V.; Gallagher, R.P.; Chatry, M.F. Larval evidence for natural reproduction of the grass carp (Ctenopharngodon idella) in the lower Mississippi River. In Proceedings of the Fourth Annual Larval Fish Conference, University of Mississippi, Oxford, MS, USA, 27–28 February 1980; Fuiman, L.A., Ed.; FWS/OBS-80/43; Fish and Wildlife Service: Washington, DC, USA, 1980; pp. 1–19. Available online: http://pubs.er.usgs.gov/publication/fwsobs80_43 (accessed on 28 June 2023).
- Chapman, D.C.; Davis, J.J.; Jenkins, J.A.; Kocovsky, P.M.; Miner, J.G.; Farver, J.; Jackson, P.R. First evidence of grass carp recruitment in the Great Lakes Basin. J. Great Lakes Res. 2013, 39, 547–554. [Google Scholar] [CrossRef]
- Jenkins, J.A.; Chauvin, M.D.; Johnson, D.; Brown, B.L.; Bailey, J.; Kelly, A.M.; Kinter, B.T. Defensible standardized ploidy assessments for Grass Carp (Ctenopharyngodon idella, Cyprinidae) intercepted from the commercial supply chain. J. Great Lakes Res. 2019, 45, 371–383. [Google Scholar] [CrossRef]
- DeBoer, J.A.; Anderson, A.M.; Casper, A.F. Multi-trophic response to invasive silver carp (Hypophthalmichthys molitrix) in a large floodplain river. Freshw. Biol. 2018, 63, 597–611. [Google Scholar] [CrossRef]
- Zhang, H.; Rutherfor, E.S.; Mason, D.M.; Breck, J.T.; Wittmann, M.E.; Cooke, R.M.; Lodge, D.M.; Rothlisberger, J.D.; Zhu, X.; Johnson, T.B.; et al. Forecasting the impacts of silver and bighead carp on the Lake Erie food web. Trans. Am. Fish. Soc. 2016, 145, 136–162. [Google Scholar] [CrossRef]
- Kolar, C.S.; Courtenay, W.R.; Nico, L.G. Managing undesired and invading fishes. In Inland Fisheries Management in North America, 3rd ed.; Hubert, W.A., Quist, M.C., Eds.; American Fisheries Society: Bethesda, MD, USA, 2010; pp. 213–259. [Google Scholar]
- Chick, J.H.; Maher, R.J.; Burr, B.M.; Thomas, M.R. First black carp captured in the U.S. Science 2003, 300, 1875–1876. [Google Scholar] [CrossRef]
- Nico, L.G.; Jelks, H.L. The black carp in North America: An update. In Invasive Asian Carps in North America; Chapman, D.C., Hoff, M.F., Eds.; American Fisheries Society: Bethesda, MD, USA, 2011; Volume 74, pp. 89–104. [Google Scholar]
- Whitledge, G.W.; Kroboth, P.T.; Chapman, D.C.; Phelps, Q.E.; Sleeper, W.; Bailey, J.; Jenkins, J.A. Estabishment of invasive Black Carp (Mylopharyngodon piceus) in the Mississippi River Basin: Identifying sources and year classes contributing to recruitment. Biol. Invasions 2022, 24, 3885–3904. [Google Scholar] [CrossRef]
- Office of the Federal Register. Title 50—Wildlife and Fisheries.Chapter 1—United States Fish and Wildlife Service, Department of the Interior.Part 16—Injurious Wildlife; 50CFR16.13; U.S. Government Publishing Office: Washington, DC, USA, 1996; pp. 1–7.
- Hubert, T.D. Environmental fate and effects of the lampricide TFM: A review. J. Great Lakes Res. 2003, 29, 456–474. [Google Scholar] [CrossRef]
- Willis, D.J.; Hoyer, M.V.; Canfield, D.E.; Lindberg, W.J. Training grass carp to respond to sound for potential lake management uses. N. Am. J. Aquac. 2002, 22, 208–212. [Google Scholar] [CrossRef]
- Kroboth, P.T.; Chapman, D.C.; Steevens, J.A.; Byrd, C.G. Ingested toxicity of antimycin A to grass carp Ctenopharyngodon idella and black carp Mylopharyngodon piceus in two carriers. Manag. Biol. Invasions 2022, 13, 737–749. [Google Scholar] [CrossRef]
- Rach, J.J.; Boogaard, M.; Kolar, C. Toxicity of rotenone and antimycin to silver carp and bighead carp. N. Am. J. Fish. Manag. 2009, 29, 388–395. [Google Scholar] [CrossRef]
- Zielinski, D.P.; Sorensen, P.W. Numeric simulation demonstrates that the upstream movement of invasive bigheaded carp can be blocked at sets of Mississippi River locks-and-dams using a combination of optimized spillway gate operations, lock deterrents, and carp removal. Fishes 2021, 6, 10. [Google Scholar] [CrossRef]
- Department of Prevention, Pesticides and Toxic Substances. Reregistration Eligibility Decision for Antimycin A.; U.S. Environmental Protection: Washington, DC, USA, 2007; 34p.
- Zhang, Z.; Huang, L.; Shulmeister, V.M.; Chi, Y.; Kim, K.K.; Hung, L.W.; Crofts, A.R.; Berry, E.A.; Kim, S.H. Electron transfer by domain movement in cytochrome bc1. Nature 1998, 392, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Pinho, B.R.; Santos, M.M.; Fonseca-Silva, A.; Valentao, P.; Andrade, P.B.; Oliveira, J.M.A. How mitochondrial dysfunction affects zebrafish development and cardiovascular function: An in vivo model for testing mitochondria-targeted drugs. Br. J. Pharmacol. 2013, 169, 1072–1090. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Lee, H.J.; Kim, W.J.; Lee, J.H.; Min, K.S. Mitochondrial cytochrome b sequence variation in Korean salmonids. J. Fish Biol. 2020, 56. [Google Scholar] [CrossRef]
- Khalifa, M.A.; Younes, M.I.; Ghazy, A. Cytochrome b shows signs of adaptive protein evolution in Gerbillus species from Egypt. J. Basic Appl. Zool. 2018, 79, 1. [Google Scholar] [CrossRef]
- Smith, M.J.; Simco, B.A.; Warren, C.O. Comparative effects of antimycin A on isolated mitochondria of channel catfish (Ictalurus punctatus) and rainbow trout (Salmo gairdneri). Comp. Biochem. Physiol. 1975, 52, 113–117. [Google Scholar] [CrossRef]
- U.S. Geological Survey. Nonindigenous Aquatic Species Database. Available online: https://nas.er.usgs.gov (accessed on 22 June 2022 and 28 June 2023).
- Walker, C.R.; Lennon, R.E.; Berger, B.L. Preliminary Observations on the Toxicity of Antimycin A to Fish and Other Aquatic Animals; Investigations in Fish Control; U.S. Department of the Interior, Bureau of Sport Fisheries and Wildlife: Washington, DC, USA, 1964.
- Berger, B.L.; Lennon, R.E.; Hogan, J.W. Ecotox: Laboratory Studies on Antimycin A as a Fish Toxicant; Investigation in Fish Control No.26, Fish Wildl. Serv., Bur. Sport Fish. Wildl.; USDI: Washington, DC, USA, 1969; 21p.
- Marking, L.L.; Bills, T.D. Sensitivity of four species of carp to selected fish toxicants. N. Am. Jounal Fish. Manag. 1981, 1, 51–54. [Google Scholar] [CrossRef]
- Gilderhus, P.A. Exposure times necessary for antimycin and rotenone to eliminate certain freshwater fish. J. Fish. Res. Board Can 1972, 29, 199–202. [Google Scholar] [CrossRef]
- Mayer, F.L.; Ellersieck, M.R. Manual of Acute Toxicity: Interpretation and Data Base for 410 Chemicals and 66 Species of Freshwater Animals; Technical Report PB86239878; U.S. Department of the Interior Fish and Wildlife Service: Columbia, MO, USA, 1986; USDI Fish and Wildlife Service, Publication No.160, Washington, DC; 588p. Available online: https://ntrl.ntis.gov/NTRL/dashboard/searchResults/titleDetail/PB86239878.xhtml# (accessed on 28 June 2023).
- Chapman, D.; Fairchild, J.; Carollo, B.; Deters, J.; Feltz, K.; Witte, C. An Examination of the Sensitivity of Bighead Carp and Silver Carp to Antimycin A and Rotenone; US Geological Survey: Columbia, MO, USA, 2003; pp. 1–22.
- Henderson, S. Preliminary studies on the tolerance of the White Amur, Ctenopharyngodon idella, to Rotenone and other commonly used pond treatment chemicals. In Proceedings of the Twenty-Seventh Annual Conference of the Southeastern Association of Game and Fish Commissioners, Hot Springs, AR, USA, 14–17 October 1973; United States Environmental Protection Agency: Washington, DC, USA, 1974; Volume 27, pp. 435–447. [Google Scholar]
- Finlayson, B.J.; Schnick, R.A.; Cailteux, R.L.; DeMong, L.; Horton, W.D.; McClay, W.; Thompson, C.W. Assessment of antimycin a use in fisheries and its potential for reregistration. Fisheries 2002, 27, 10–18. [Google Scholar] [CrossRef]
- Marking, L.L.; Dawson, V.K. The half-life of biological activity of antimycin determined by fish bioassay. Trans. Am. Fish. Soc. 1972, 101, 100–105. [Google Scholar] [CrossRef]
- Ott, K.C. Antimycin. A brief review of it’s chemistry, environmental fate, and toxicology. Biochem. Et Biophys. Acta 1994, 1185, 1–9. [Google Scholar]
- Clearwater, S.J.; Hickey, C.W.; Martin, M.L. Overview of Potential Piscicides and Molluscicides for Controlling Aquatic Pest Species in New Zealand; Department of Conservation: Wellington, New Zealand, 2008; p. 74.
- Kim, H.; Esser, L.; Hossain, M.B.; Xia, D.; Yu, C.; Rizo, J.; van der Helm, D.; Deisenhofer, J. Structure of antimycin A1, a specific electron transfer inhibitor of Ubiquinol−Cytochrome c Oxidoreductase. J. Am. Chem. Soc. 1999, 121, 4902–4903. [Google Scholar] [CrossRef]
- Ridgway, J.L.; Lawson, K.M.; Shier, S.A.; Calfee, R.D.; Chapman, D.C. An assessment of fish herding techniques: Management implications for mass removal and control of silver carp. N. Am. J. Fish. Manag. 2023, 43, 176–188. [Google Scholar] [CrossRef]
- Chapman, D.C. "Modified Unified Method" of carp capture; U.S. Geological Survey Fact Sheet 2020–3005; US Geological Survey: Columbia, MO, USA, 2020; 2p. [CrossRef]
- Sun, H.; Zhang, A.; Yan, G.; Zhang, Y.; Meng, X.; Liu, L.; Xie, N.; Cheng, W.; Wang, X. Acupuncture targeting and regulating multiple signaling pathways related to Zusanli acupoint using iTRAQ-based quantitative proteomic analysis. Acupunct. Relat. Ther. 2014, 2, 51–56. [Google Scholar] [CrossRef]
- Huang, L.; Cobessi, D.; Tung, E.Y.; Berry, E.A. Binding of the respiratory chain inhibitor antimycin to the mitochondrial bc1 complex: A new crystal structure reveals an altered intramolecular hydrogen-bonding pattern. J. Mol. Biol. 2005, 351, 573–597. [Google Scholar] [CrossRef] [PubMed]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Sunghwan, K.; et al. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef] [PubMed]
- Cupp, A.R.; Woiak, Z.; Erickson, R.A.; Amberg, J.J.; Gaikowski, M.P. Carbon dioxide as an under-ice lethal control for invasive fishes. Biol. Invasions 2017, 19, 2543–2552. [Google Scholar] [CrossRef]
- Vetter, B.J.; Murchy, K.A.; Cupp, A.R.; Amberg, J.J.; Gaikowski, M.P.; Mensinger, A.F. Acoustic deterrence of bighead carp (Hypophthalmichthys nobilis) to a broadband sound stimulus. J. Great Lakes Res. 2017, 43, 163–171. [Google Scholar] [CrossRef]
- Lennon, R.E.; Berger, B.L. A resume on field applications of antimycin A to control fish. In Investigations in Fish Control; U.S. Bureau of Sport Fisheries and Wildlife: Washington, DC, USA, 1970; pp. 1–23. [Google Scholar]
- Rach, J.J.; Luoma, J.A.; Marking, L.L. Development of an antimycin-impregnated bait for controlling common carp. N. Am. J. Fish. Manag. 1994, 14, 442–446. [Google Scholar] [CrossRef]
- Miyoshi, H.; Tokutake, N.; Imaeda, Y.; Akagi, T.; Iwamura, H. A model of antimycin A binding based on structure-activity studies of synthetic antimycin A analogues. Biochim. Et Biophys. Acta 1995, 1229, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Kuhlman, B.; Bradley, P. Advances in protein structure prediction and design. Nat. Rev. Mol. Cell Biol. 2019, 20, 681–697. [Google Scholar] [CrossRef]
- Zhao, B.; Katuwawala, A.; Oldfield, C.J.; Dunker, A.K.; Faraggi, E.; Gsponer, J.; Kloczkowski, A.; Malhis, N.; Mirdita, M.; Obradovic, Z.; et al. DescribePROT: Database of amino acid-level protein structure and function predictions. Nucleic Acids Res. 2021, 49, D298–D308. [Google Scholar] [CrossRef] [PubMed]



| Genus Species | Common Name | Family | Concentration Range | Effect | Author |
|---|---|---|---|---|---|
| Dorosoma cepedianum | American Gizzard Shad | Clupeidae | 0.06–0.8 μg/L | Mortality | [28] |
| Cyprinus carpio | Common Carp | Cyprinidae | 0.04–100 μg/L | Mortality | [29] |
| C. carpio | Common Carp | Cyprinidae | 4.0 mg/kg body weight | LD50 (gavage) | [2] |
| C. carpio | Common Carp | Cyprinidae | 0.57 µg/L | LC50 | [30] |
| Pimephales promelas | Fathead Minnow | Cyprinidae | 0.025–20.5 μg/L | Mortality | [31,32] |
| P. promelas | Fathead Minnow | Cyprinidae | 0.08–0.11 µg/L | LC50 | [18] |
| Hypophthalmichthys nobilis | Bighead Carp | Cyprinidae | 0.60 μg/L | LC50 | [30] |
| H. nobilis | Bighead Carp | Cyprinidae | 7.50 μg/L | Mortality at 60% | [33] |
| H. molitrix | Silver Carp | Cyprinidae | 0.83 μg/L | LC50 | [30] |
| H. molitrix | Silver Carp | Cyprinidae | 7.50 μg/L | Mortality at 20% | [33] |
| H. nobilis & H. molitrix | Bighead & Silver Carp | Cyprinidae | 2.50 μg/L | No mortality | [33] |
| H. nobilis & H. molitrix | Bighead & Silver Carp | Cyprinidae | 5.00 μg/L | No mortality | [33] |
| Ctenopharyngodon idella | Grass Carp | Cyprinidae | 1.00 µg/L | LC50 | [30] |
| C. idella | Grass Carp | Cyprinidae | 1 μg/L | Mortality at 100% | [34] |
| C. idella | Grass Carp | Cyprinidae | 4–8 mg/L b | Mortality at 100% | [18] |
| Mylopharyngodon piceus | Black Carp | Cyprinidae | 4 to >16 mg/L b | Mortality at 100% | [18] |
| Ictalurus punctatus | Channel Catfish | Ictaluridae | 1.36–22 μg/L | Mortality | [29,32] |
| I. punctatus | Channel Catfish | Ictaluridae | 9.00 μg/L | LC50 | [29] |
| Oreochromis mossambicus | Tilapia Mossambica | Cichlidae | 5 μg/L | Mortality | [29] |
| Lepomis macrochirus | Bluegill | Centrarchidae | 0.0339–100 μg/L | Mortality | [29] |
| L. marochirus | Bluegill | Centrarchidae | 0.14 μg/L | LC50 | [29] |
| Perca flavescens | Yellow Perch | Percidae | 0.06–0.66 μg/L | Mortality | [28] |
| Oncorhyncus mykiss | Rainbow Trout | Salmonidae | 0.04 μg/L | LC50 | [35] |
| O. mykiss | Rainbow Trout | Salmonidae | 0.05–20.40 μg/L | LC50 | [36] |
| O. mykiss | Rainbow Trout | Salmonidae | 0.00003 μg/L | Mortality | [36] |
| Salvelinus fontinalis | Brook Trout | Salmonidae | 1 μg/L | Mortality | [29] |
| S. fontinalis | Brook Trout | Salmonidae | 0.035 μg/L | EC50 | [29] |
| Sander vitreus | Walleye | Percidae | 0.10 μg/L–0.60 μg/L | Mortality | [28] |
| Fish | Number of Accessions | Motifs | Similarity within Species (%) | Variation from the CCM (%) |
|---|---|---|---|---|
| Common Carp Cyprinus carpio (Cyprinidae) | 203 | 1 15DALVD33 | 95.54 | 4.46 |
| 2 33GSLLGLC195 | 96.57 | 3.43 | ||
| 3 195HLLF203 | 100 | 0 | ||
| 4 203GSNN218 | 100 | 0 | ||
| 5 218SFHP226 | 100 | 0 | ||
| 6 226KDLL380 | 100 | 0 | ||
| Bighead Carp Hypophthalmichthys nobilis (Cyprinidae) | 48 | 1 15DALVD33 | 97.87 | 2.13 |
| 2 33GSLLGLC195 | 100 | 0 | ||
| 3 195HLLF203 | 100 | 0 | ||
| 4 203GSNN218 | 100 | 0 | ||
| 5 218SFHP226 | 100 | 0 | ||
| 6 226KDLL380 | 100 | 0 | ||
| Silver Carp H. molitrix (Cyprinidae) | 122 | 1 15DALVD33 | 99.10 | 0.90 |
| 2 33GSLLGLC195 | 100 | 0 | ||
| 3 195HLLF203 | 100 | 0 | ||
| 4 203GSNN218 | 100 | 0 | ||
| 5 218SFHP226 | 100 | 0 | ||
| 6 226KDLL380 | 100 | 0 | ||
| Grass Carp Ctenopharyngodon idella (Cyprinidae) | 96 | 1 15DALVD33 | 100 | 0 |
| 2 33GSLLGLC195 | 100 | 0 | ||
| 3 195HLLF203 | 100 | 0 | ||
| 4 203GSNN218 | 100 | 0 | ||
| 5 218SFHP226 | 100 | 0 | ||
| 6 226KDLL380 | 100 | 0 | ||
| Black Carp Mylopharyngodon piceus (Cyprinidae) | 46 | 1 15DALVD33 | 100 | 0 |
| 2 33GSLLGLC195 | 100 | 0 | ||
| 3 195HLLF203 | 100 | 0 | ||
| 4 203GSNN218 | 100 | 0 | ||
| 5 218SFHP226 | 100 | 0 | ||
| 6 226KDLL380 | 100 | 0 | ||
| Channel Catfish Ictalurus punctatus (Ictaluridae) | 48 | 1 14NALID32 | 100 | 100 |
| 2 32GSLLLLC194 | 96.77 | 96.77 | ||
| 3 194HALF202 | 96.00 | 96.00 | ||
| 4 202GSNN217 | 100 | 0 | ||
| 5 217SFHP225 | 100 | 0 | ||
| 6 225KDLL380 | 100 | 0 | ||
| Bluegill Lepomis macrochirus (Centrachidae) | 20 | 1 15DALVD33 | 100 | 0 |
| 2 33GSLLGLC195 | 100 | 0 | ||
| 3 195HLLF203 | 100 | 0 | ||
| 4 203GSNN218 | 100 | 0 | ||
| 5 218LFHP226 | 53.33 | 53.33 | ||
| 6 226KDLL380 | 100 | 0 | ||
| Fathead Minnow Pimephales promelas (Cyprinidae) | 38 | 1 15GALVD33 | 78.38 | 78.38 |
| 2 33GSLLGLC195 | 97.37 | 2.63 | ||
| 3 195HLLF203 | 100 | 0 | ||
| 4 203GSNN218 | 100 | 0 | ||
| 5 218SFHP226 | 100 | 0 | ||
| 6 226KDLL380 | 100 | 0 | ||
| Walleye Sander vitreus (Percidae) | 71 | 1 14DALVD32 | 100 | 0 |
| 2 32GSLLGLC194 | 100 | 0 | ||
| 3 194HLLF202 | 100 | 0 | ||
| 4 202GSNN217 | 100 | 0 | ||
| 5 217SFHP225 | 100 | 0 | ||
| 6 226KDLL380 | 100 | 0 | ||
| Yellow Perch Perca flavescens (Percidae) | 41 | 1 14NALVD32 | 95.12 | 100 |
| 2 32GSLLGLC194 | 100 | 0 | ||
| 3 194HLLF202 | 100 | 0 | ||
| 4 202GSNN217 | 100 | 0 | ||
| 5 217SFHP225 | 100 | 0 | ||
| 6 225KDLL398 | 100 | 0 | ||
| Nile Tilapia Oreochromis niloticus (Cichlidae) | 49 | 1 15DALVD33 | 97.87 | 2.13 |
| 2 33GSLLGLC195 | 100 | 0 | ||
| 3 195HLIF203 | 100 | 100 | ||
| 4 203GSNN218 | 100 | 0 | ||
| 5 218SFHP226 | 100 | 0 | ||
| 6 226KDLL380 | 100 | 0 | ||
| American Gizzard Shad Dorosoma cepedianum (Clupeidae) | 10 | 1 15DALVD33 | 100 | 0 |
| 2 33GSLLGLC195 | 100 | 0 | ||
| 3 195HLLF203 | 100 | 0 | ||
| 4 203GSNN218 | 100 | 0 | ||
| 5 218SFHP226 | 100 | 0 | ||
| 6 226KDLL380 | 100 | 0 |
| CCM Amino Acid | Variant Amino Acid | |||||
|---|---|---|---|---|---|---|
| Common Name | Scientific Name | Name | R Group | Name | R Group | Charge Characteristics |
| Fathead Minnow | Pimephales promelas | D: Aspartic Acid | CH2COOH | G: Glycine | H | Non-polar, was polar and negatively charged |
| Bluegill | Lepomis macrochirus | S: Serine | CH2OH | L: Leucine | CH2CH(CH3)2 | Non-polar, was polar |
| Yellow Perch | Perca flavescens | D: Aspartic Acid | CH2COOH | N: Asparagine | CH2CONH2 | Polar uncharged, was polar and negatively charged |
| Nile Tilapia | Oreochromis niloticus | L: Leucine | CH2CH(CH3)2 | I: Isoleucine | CH(CH3)CH2CH3 | Both are nonpolar |
| Channel Catfish | Ictalurus punctatus | D: Aspartic Acid | CH2COOH | N: Asparagine | CH2CONH2 | Polar uncharged, was polar and negatively charged |
| V: Valine | CH(CH3)2 | I: Isoleucine | CH(CH3)CH2CH3 | Both are nonpolar | ||
| G: Glycine | H | L: Leucine | CH2CH(CH3)2 | Both are nonpolar | ||
| L: Leucine | CH2CH(CH3)2 | A: Alanine | CH3 | Both are nonpolar | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baudoin, B.A.; Brown, B.L.; Calfee, R.D.; Jenkins, J.A. Amino Acid Variation at the Mitochondrial Binding Site of Antimycin A is Proposed to Reflect Sensitivity and Toxicity Differences among Fish Species. Fishes 2023, 8, 381. https://doi.org/10.3390/fishes8070381
Baudoin BA, Brown BL, Calfee RD, Jenkins JA. Amino Acid Variation at the Mitochondrial Binding Site of Antimycin A is Proposed to Reflect Sensitivity and Toxicity Differences among Fish Species. Fishes. 2023; 8(7):381. https://doi.org/10.3390/fishes8070381
Chicago/Turabian StyleBaudoin, Brooke A., Bonnie L. Brown, Robin D. Calfee, and Jill A. Jenkins. 2023. "Amino Acid Variation at the Mitochondrial Binding Site of Antimycin A is Proposed to Reflect Sensitivity and Toxicity Differences among Fish Species" Fishes 8, no. 7: 381. https://doi.org/10.3390/fishes8070381
APA StyleBaudoin, B. A., Brown, B. L., Calfee, R. D., & Jenkins, J. A. (2023). Amino Acid Variation at the Mitochondrial Binding Site of Antimycin A is Proposed to Reflect Sensitivity and Toxicity Differences among Fish Species. Fishes, 8(7), 381. https://doi.org/10.3390/fishes8070381