
Inferences about Fishing Impacts on the Reproductive Pattern of Torroto Grunt—Genyatremus luteus (BLOCH, 1790)—From the Amazon Continental Shelf


Abstract
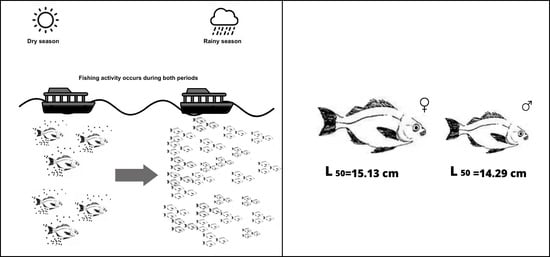
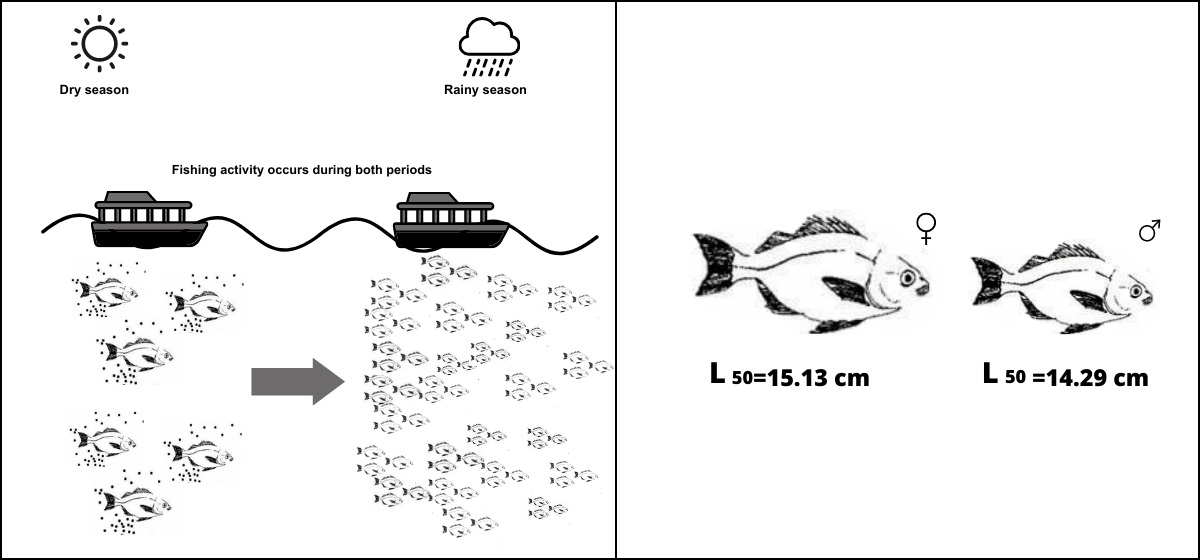
1. Introduction
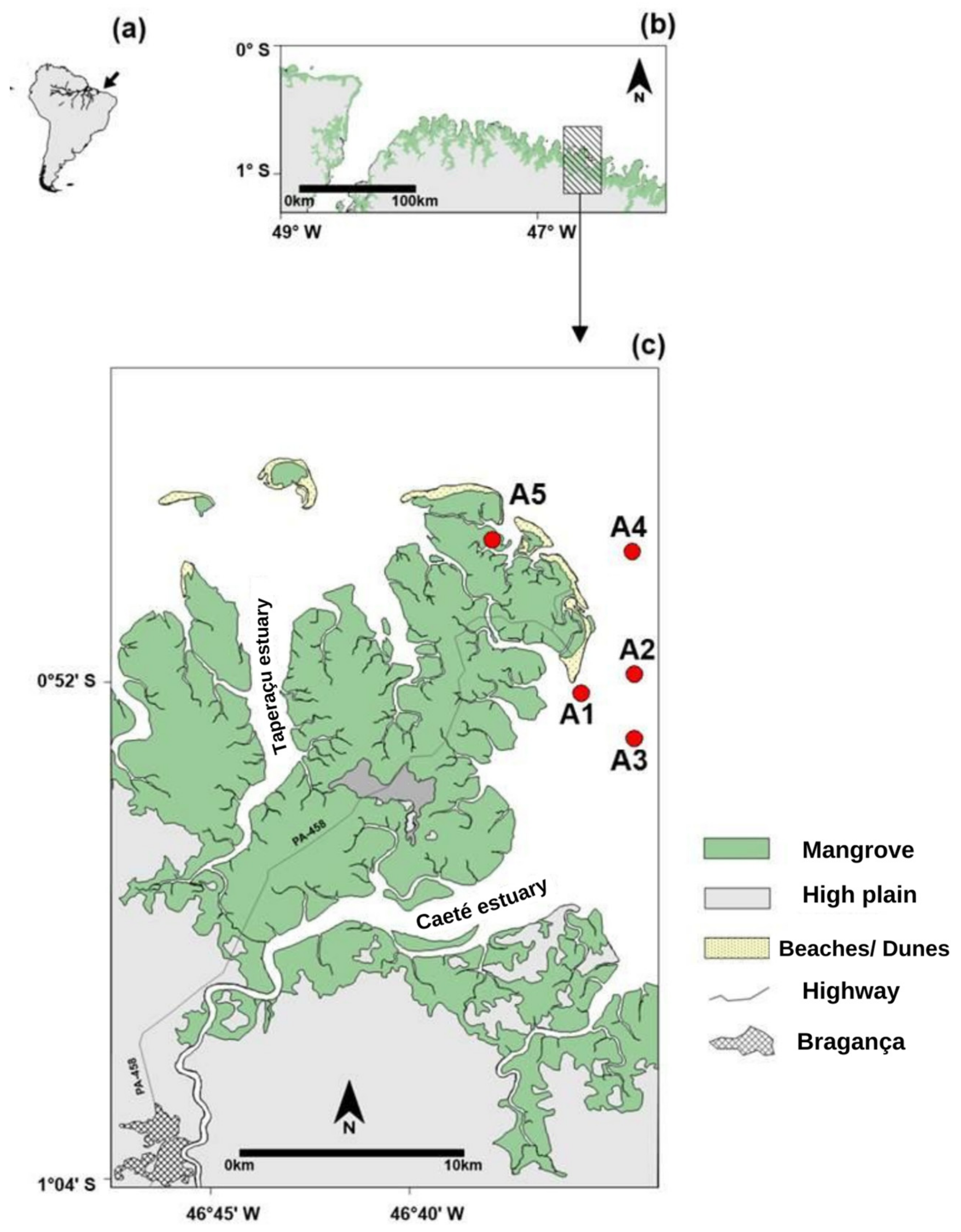
2. Materials and Methods
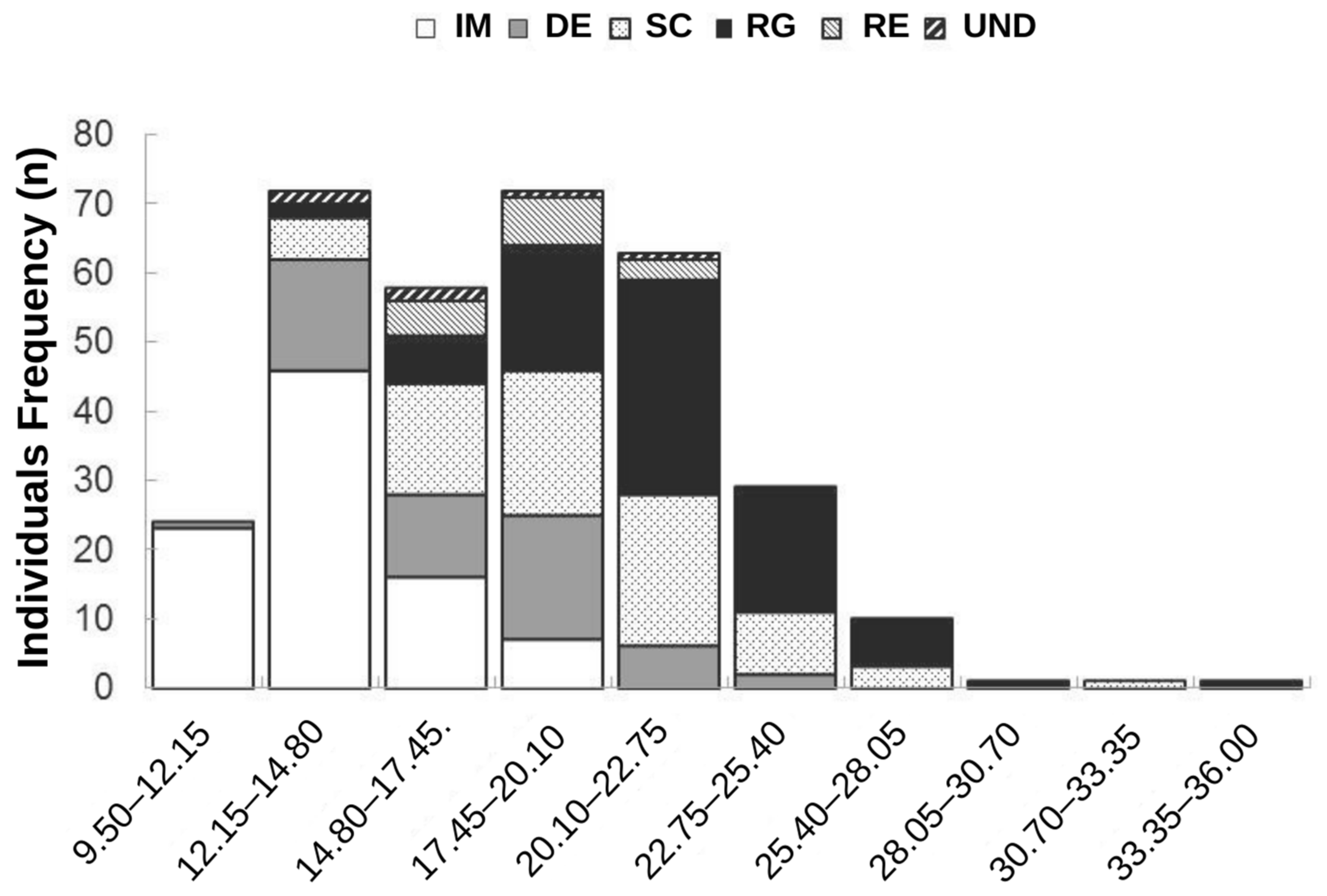
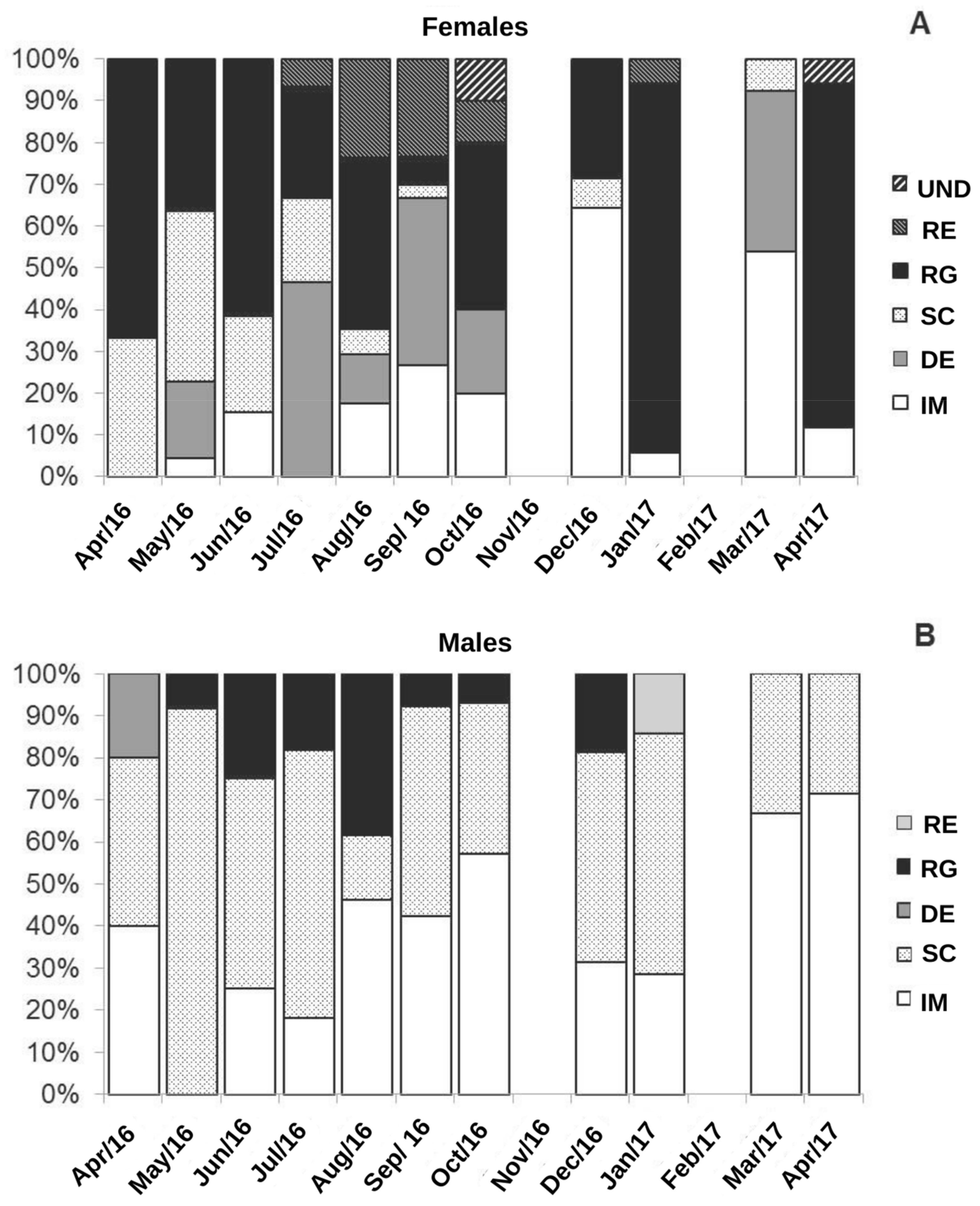
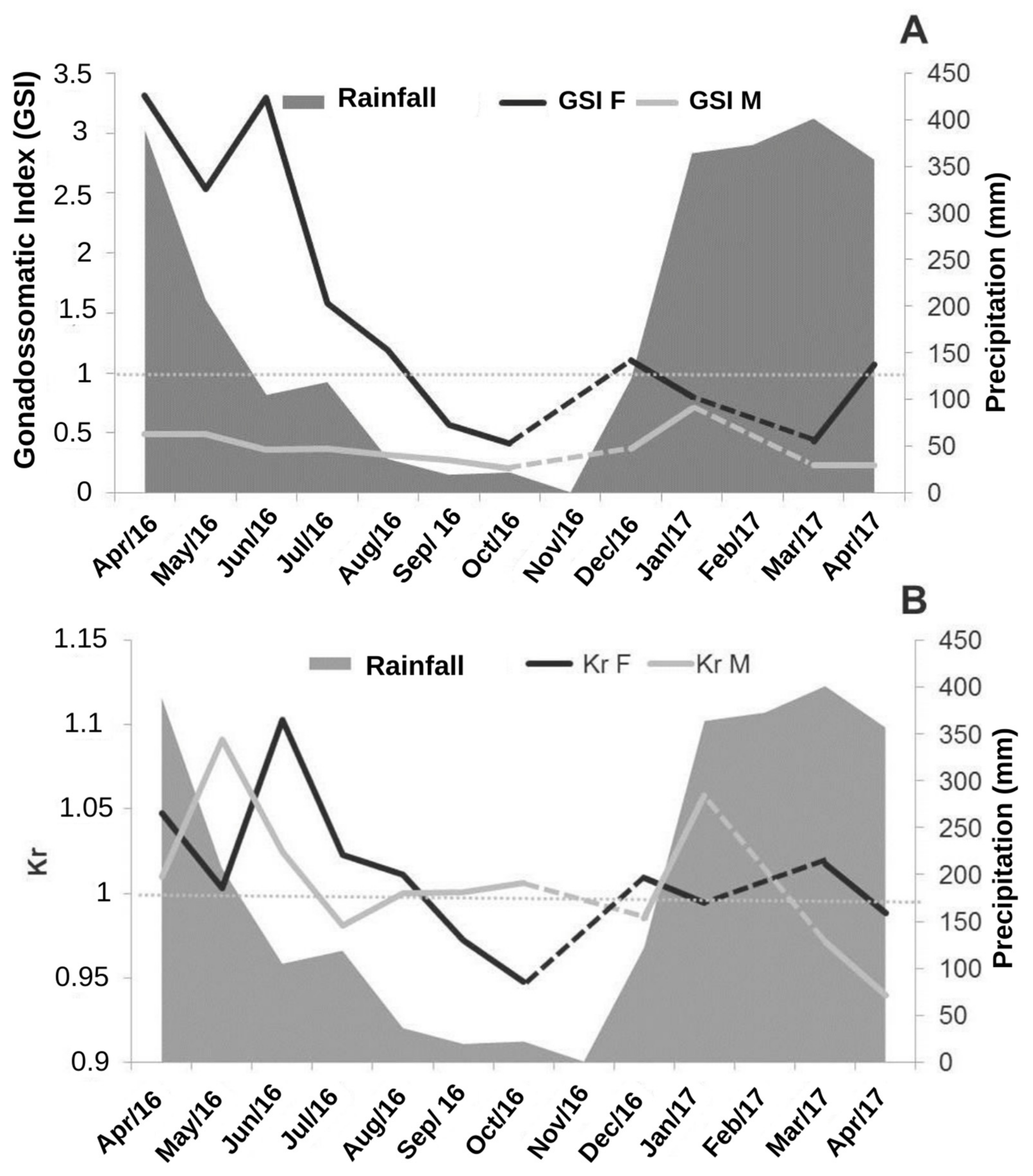
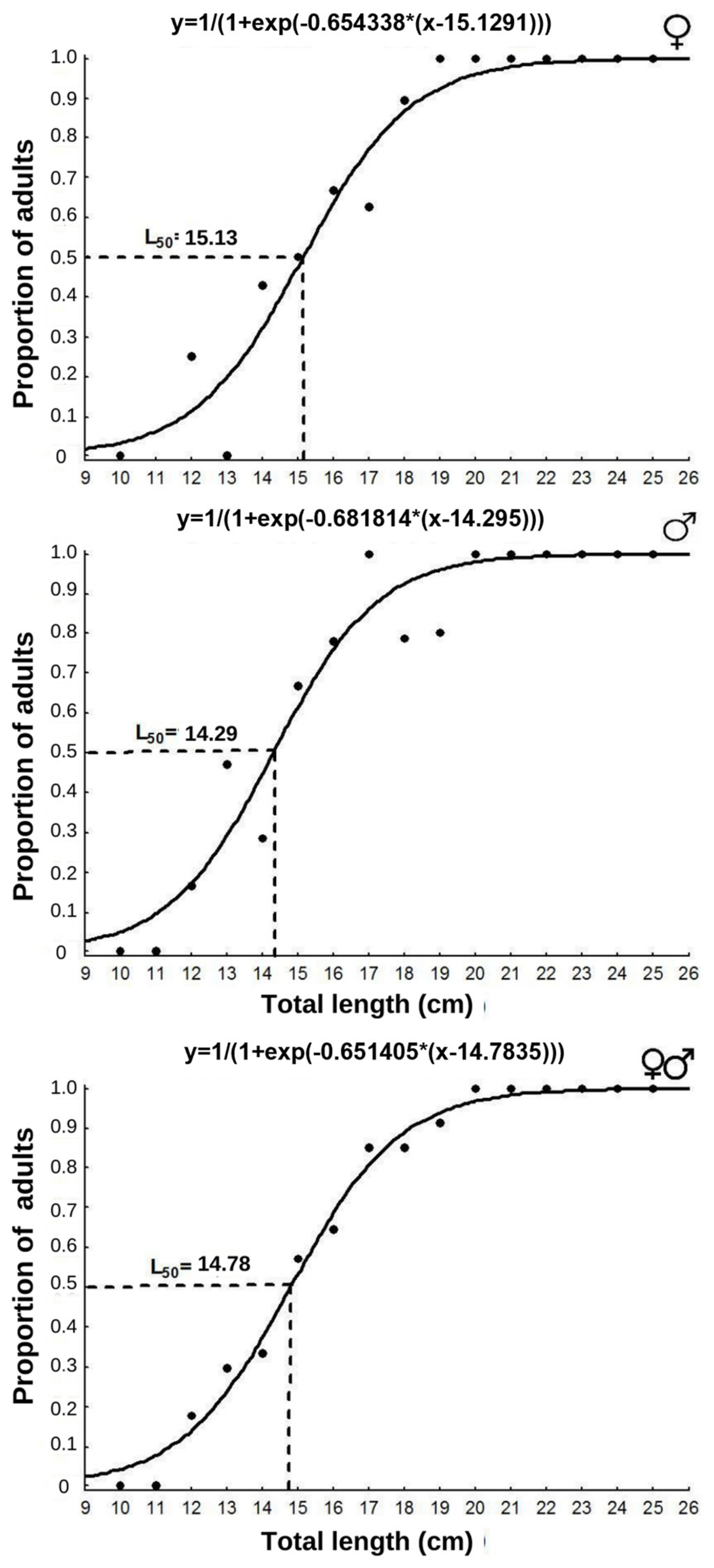
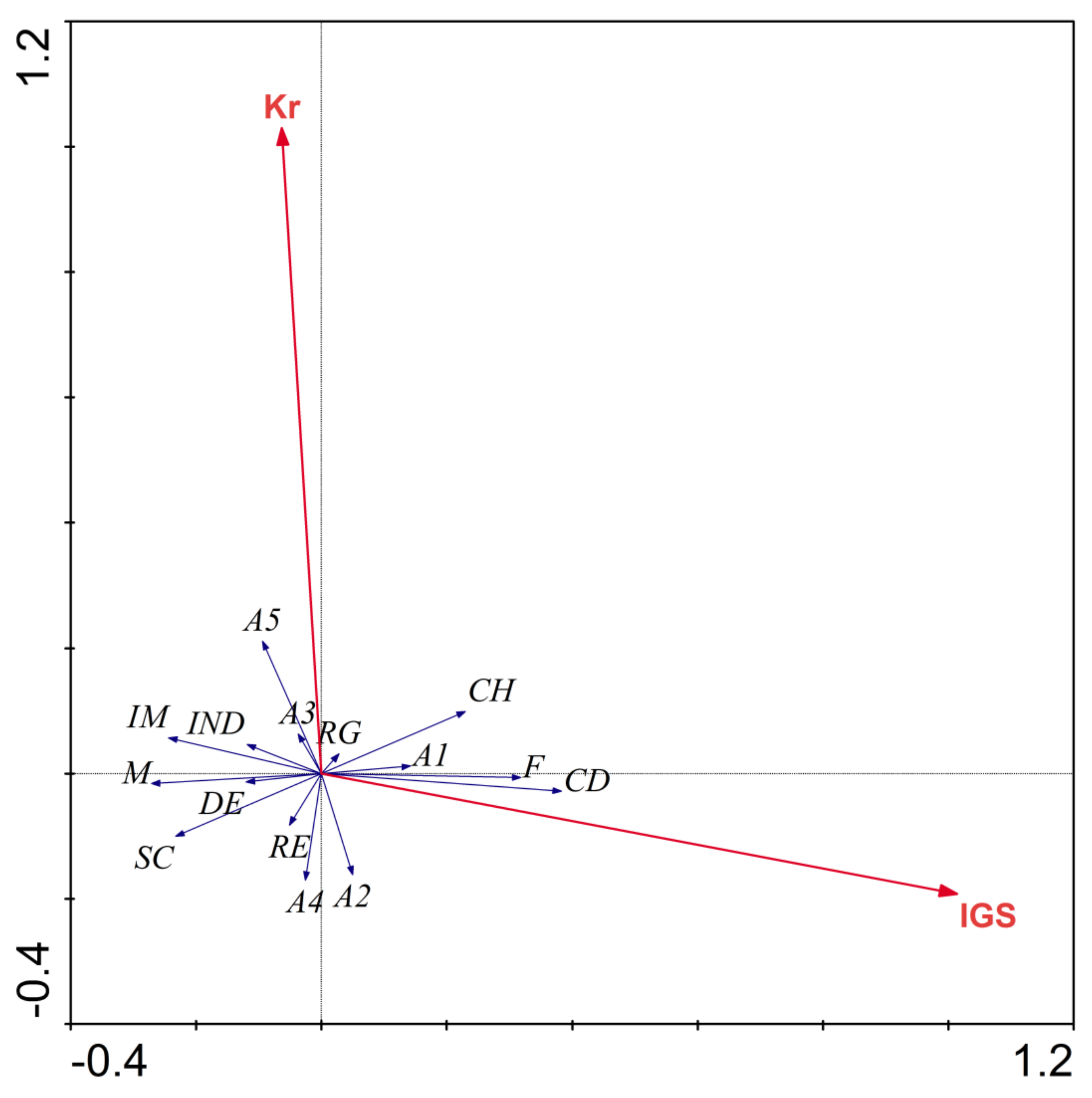
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Winemiller, K.O. Life history strategies, population regulation, and implications for fisheries management. Can. J. Fish. Aquat. Sci. 2005, 62, 872–885. [Google Scholar] [CrossRef]
- Schreck, C.B. Stress and fish reproduction: The roles of allostasis and hormesis. Gen. Comp. Endocrinol. 2010, 165, 549–556. [Google Scholar] [CrossRef]
- McEwen, B.S.; Wingfield, J.C. The concept of allostasis in biology and biomedicine. Horm. Behav. 2003, 43, 2–15. [Google Scholar] [CrossRef]
- Fonteles Filho, A.A. Oceanografia, Biologia e Dinâmica Populacional de Recursos Pesqueiros; Expressão Gráfica e Editora: Fortaleza, Brazil, 2011. [Google Scholar]
- Barletta-Bergan, A.; Barletta, M.; Saint-Paul, U. Structure and Seasonal Dynamics of Larval Fish in the Caeté River Estuary in North Brazil; Estuarine, Coastal and Shelf Science: Bragança, PA, Brazil, 2002; Volume 54, pp. 193–206. [Google Scholar]
- Barletta-Bergan, A. An Evaluation of the Larval Fish Assemblage in a North Brazilian Mangrove Area. In Mangrove Dynamics and Management in North Brazil; Saint-Paul, U., Schneider, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 209–220. [Google Scholar]
- Giarrizzo, T.; Krumme, U. Spatial differences and seasonal cyclicity in the intertidal fish fauna from four mangrove creeks in a salinity zone of the Curuçá estuary, north Brazil. Bull. Mar. Sci. 2007, 80, 739–754. [Google Scholar]
- Barletta, M.; Saint-Paul, U. Distribution pattern of fish in a mangrove estuary. In Mangrove Dynamics and Management in North Brazil; Saint-Paul, U., Schneider, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 171–208. [Google Scholar]
- Fernandes, S.C.P.; Bentes, A.B.; Pereira, L.D.J.G.; Nascimento, M.S.; Bentes, B. Variação temporal da captura comercial do peixe pedra, Genyatremus luteus, desembarcado em um pólo pesqueiro da costa norte do Brasil–Península de Ajuruteua–Bragança-PA. Bol. Inst. Pesca 2015, 41, 173–182. [Google Scholar]
- Fernandes, S.C.P.; Pereira, L.D.J.G.; Nascimento, M.S.; Andrade, C.E.R.; Bentes, B. Biologia Populacional do peixe pedra, Genyatremus luteus (BLOCH, 1790), costa amazônica brasileira. Bol. Inst. Pesca 2017, 43, 527–541. [Google Scholar] [CrossRef]
- Lindeman, K.C.; Richards, W.J. Haemulidae: Grunts. In Early Stages of Atlantic Fishes: An Identification Guide for Western Central North Atlantic; Richards, W.J., Ed.; CRC Press: Boca Raton, FL, USA, 2006; pp. 1597–1646. [Google Scholar]
- Maneschy, M.C. Pescadores curralistas no litoral do estado do Pará: Evolução e continuidade de uma pesca tradicional. Rev. Soc. Bras. Hist. Ciênc. 1993, 10, 53–73. [Google Scholar]
- Brown-Peterson, N.J.; Wyanski, D.M.; Saborido-Rey, F.; Macewicz, B.J.; Lowerre-Barbieri, S.K. A Standardized terminology for describing reproductive development in fishes. Mar. Coast. Fish. Dyn. Manag. Ecosyst. Sci. 2011, 3, 52–70. [Google Scholar] [CrossRef]
- Moraes, B.C.; Costa, J.M.N.; Costa, A.C.L.; Costa, M.H. Variação espacial e temporal da precipitação no estado do Pará. Acta Amaz. 2005, 35, 207–217. [Google Scholar] [CrossRef]
- King, M. Fisheries Biology, Assessment and Management; Fishing News Books: Oxford, UK, 1995. [Google Scholar]
- Le Cren, E.D. The length-weight relationship and seasonal cycle in gonad weight and condition in the perch (Perca fluviatilis). J. Anim. Ecol. 1951, 20, 201–219. [Google Scholar] [CrossRef]
- Vazzoler, A.E.A.M. Biologia da Reprodução de Peixes Teleósteos: Teoria e Prática; Eduem: Maringá, Brazil, 1996. [Google Scholar]
- Froese, R.; Tsikliras, A.C.; Stergiou, K.I. 2011: Editorial note on weight–length relations of fishes. Acta Ichthyol. Piscat. 2011, 41, 261–263. [Google Scholar] [CrossRef]
- Haimovici, M.; Canziani, G.V. Length-weight relationship of marine fishes from Southern Brazil. Environ. Sci. 2000, 23, 19–23. [Google Scholar]
- Froese, R. Cube law, condition factor and weight–length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Elliott, M.; Whitfield, A.K.; Potter, I.C.; Blaber, S.J.; Cyrus, D.P.; Nordlie, F.G.; Harrison, T.D. The guild approach to categorizing estuarine fish assemblages: A global review. Fish Fish. 2007, 8, 241–268. [Google Scholar] [CrossRef]
- Andrade-Tubino, M.F.; Ribeiro, A.L.R.; Vianna, M. Organização espaço-temporal das ictiocenoses demersais nos ecossistemas estuarinos brasileiros: Uma síntese. Oecol. Aust. 2009, 12, 640–661. [Google Scholar] [CrossRef]
- Auth, T.D.; Daly, E.A.; Brodeur, R.D.; Fisher, J.L. Phenological and distributional shifts in ichthyoplankton associated with recent warming in the northeast Pacific Ocean. Glob. Chang. Biol. 2018, 24, 259–272. [Google Scholar] [CrossRef]
- Mavruk, S.; Bengil, F.; Yüksek, A.; Özyurt, C.E.; Kiyağa, V.B.; Avşar, D. Intra-annual patterns of coastal larval fish assemblages along environmental gradients in the northeastern Mediterranean. Fish. Oceanogr. 2017, 27, 232–245. [Google Scholar] [CrossRef]
- Jalabert, B. Particularities of reproduction and oogenesis in teleost fish compared to mammals. Reprod. Nutr. Dev. 2005, 45, 261–279. [Google Scholar] [CrossRef]
- Noleto-Filho, E.M.; Yauri, W.L.M.; Luvizotto-Santos, R. Captura de reprodutores de peixe-pedra Genyatremus luteus (Bloch, 1797) (Pisces: Haemulidae) e manutenção em sistema fechado. Bol. Lab. Hidrobiol. 2012, 1, 55–60. [Google Scholar]
- Nikolskii, G.V. The Ecology of Fishes; Academic Press: New York, NY, USA, 1963. [Google Scholar]
- Johannes, R.E. Reproductive strategies of coastal marine fishes in the tropics. Environ. Biol. Fishes 1978, 3, 65–84. [Google Scholar] [CrossRef]
- Gómez, G.; Guzman, R.; Chacon, R. Algunos aspectos de La biologia reproductiva y poblacional del torroto, Genyatremus luteus, (Block, 1797) (Pisces: Haemulidae) em el golfo de Parla, Venezuela. Zootec. Trop. 2002, 20, 223–234. [Google Scholar]
- Marques, N.E.F.; Sampaio, J.A.A.; Bezerra, S.F.B.; Yauri, W.M.; Santos, R.L. Indução hormonal à desova do peixe pedra Genyatremus luteus (Bloch, 1790)(Teleostei: Haemulidae). Bol. Lab. Hidrobiol. 2016, 26, 32–35. [Google Scholar]
- Moura, H.T.G.S.; Nunes, Z.M.P. Caracterização sazonal das águas do sistema estuarino do Caeté (Bragança-PA). Bol. Inst. Pesca 2016, 42, 844–854. [Google Scholar] [CrossRef]
- Oliveira, S.M.O.; Pereira, L.C.C.; Vila-Concejo, A. Processos morfodinâmicos em uma praia de macromaré no litoral amazônico. Quat. Environ. Geosci. 2014, 5, 125–136. [Google Scholar] [CrossRef]
- Silva, A.M.; Oliveira, M.R.; Chellappa, S. Biologia reprodutiva do coró, Pomadasys corvinaeformis Steindachner (Osteichthyes: Haemulidae) das águas costeiras do Rio Grande do Norte, Brasil. Biota Amaz. 2012, 2, 15–24. [Google Scholar] [CrossRef]
- Shatunovskiy, M.I. Some indices of reproductive ability of fishes. J. Ichthyol. 1988, 28, 119–123. [Google Scholar]
- Araújo, B.R. Desova e fecundidade em peixes de água doce e marinhos. Rev. Biol. Ciênc. Terra 2009, 9, 24–31. [Google Scholar]
- Wooton, R.J. Ecology of Teleost Fishes; Chapman and Hall: London, UK, 1990. [Google Scholar]
- Domínguez-Petit, R.; Korta, M.; Saborido-Rey, F.; Murua, H.; Sainza, M.; Piñeiro, C. Changes in size at maturity of European hake Atlantic populations in relation with stock structure and environmental regimes. J. Mar. Syst. 2008, 71, 260–278. [Google Scholar] [CrossRef]
- Lojo, D.; Cousido-Rocha, M.; Cerviño, S.; Dominguez-Petit, R.; Sainza, M.; Pennino, M.G. Assessing changes in size at maturity for the European hake (Merluccius merluccius) in Atlantic Iberian waters. Sci. Mar. 2022, 86, e046. [Google Scholar] [CrossRef]
- Lima, W.M.G.D.; Pinaya, W.H.D.; Lutz ÍAD, F.; Nascimento, M.S.D.; Silva, L.E.O.D.; Cintra, I.H.A.; Bentes, B. Effects of the trawling industrial fishery system on fish community structure on the inner Amazon shelf. Ocean. Coast. Res. 2021, 69, 9–10. [Google Scholar] [CrossRef]
- De Souza-Araujo, J.; Hussey, N.E.; Hauser-Davis, R.A.; Rosa, A.H.; de Oliveira Lima, M.; Giarrizzo, T. Human risk assessment of toxic elements (As, Cd, Hg, Pb) in marine fish from the Amazon. Chemosphere 2022, 301, 134575. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month/Year | Sex | N | Total Length (cm) | Weight (g) | ||
|---|---|---|---|---|---|---|
| Min–Max | Average ± SD | Min–Max | Average ± SD | |||
| Apr/16 | F | 6 | 18.20–35.40 | 23.38 ± 5.66 | 114.00–771.00 | 277.67 ± 224.40 |
| M | 4 | 13.90–22.90 | 17.75 ± 3.70 | 47.00–198.00 | 112.75 ± 64.01 | |
| May/16 | F | 22 | 17.60–31.90 | 23.23 ± 3.21 | 88.00–549.00 | 235.73 ± 100.53 |
| M | 14 | 14.80–23.20 | 19.16 ± 1.90 | 74.00–233.00 | 138.43 ± 43.84 | |
| Jun/16 | F | 13 | 10.40–24.60 | 20.78 ± 4.20 | 22.00–302.00 | 194.46 ± 79.08 |
| M | 5 | 9.50–21.90 | 17.18 ± 4.39 | 16.00–188.00 | 109.80 ± 65.36 | |
| UND | 1 | 11.2 | - | 23 | - | |
| Jul/16 | F | 15 | 16.50–25.70 | 20.33 ± 2.31 | 78.00–315.00 | 159.60 ± 62.14 |
| M | 11 | 14.50–20.10 | 17.28 ± 1.86 | 51.00–127.00 | 91.73 ± 24.51 | |
| UND | 1 | 13.6 | - | 46 | - | |
| Aug/16 | F | 17 | 11.80–27.40 | 18.62 ± 4.29 | 28.00–371.00 | 135.71 ± 93.58 |
| M | 22 | 10.80–20.10 | 15.49 ± 2.90 | 22.00–147.00 | 72.95 ± 39.77 | |
| UND | 1 | 10.1 | - | 19 | - | |
| Sep/16 | F | 30 | 11.60–22.50 | 16.55 ± 2.73 | 26.00–206.00 | 84.40 ± 41.81 |
| M | 30 | 11.10–21.20 | 16.00 ± 3.21 | 25.00–180.00 | 81.73 ± 47.59 | |
| UND | 6 | 10.40–18.60 | 13.27 ± 3.11 | 18.00–114.00 | 43.50 ± 33.95 | |
| Oct/16 | F | 10 | 13.20–25.80 | 20.15 ± 3.97 | 41.00–289.00 | 152.20 ± 79.11 |
| M | 17 | 11.30–22.60 | 16.26 ± 4.44 | 25.00–190.00 | 91.59 ± 65.32 | |
| UND | 6 | 10.10–12.90 | 11.44 ± 1.00 | 20.00–45.00 | 31.40 ± 9.26 | |
| Nov/16 | F | - | - | - | - | - |
| M | - | - | - | - | - | |
| Dec/16 | F | 14 | 12.60–23.70 | 16.55 ± 3.50 | 36.00–211.00 | 90.50 ± 59.78 |
| M | 17 | 12.00–24.60 | 17.69 ± 3.86 | 32.00–252.00 | 109.53 ± 69.12 | |
| UND | 1 | 16.2 | - | 65 | - | |
| Jan/17 | F | 17 | 13.60–28.60 | 21.55 ± 3.64 | 42.00–385.00 | 190.29 ± 86.59 |
| M | 8 | 11.50–21.40 | 16.28 ± 3.37 | 28.00–151.00 | 85.38 ± 40.27 | |
| Feb/17 | F | - | - | - | - | - |
| M | - | - | - | - | - | |
| Mar/17 | F | 13 | 12.40–19.10 | 15.37 ± 2.16 | 32.00–120.00 | 68.92 ± 28.11 |
| M | 5 | 12.50–17.90 | 15.54 ± 1.92 | 35.00–98.00 | 68.00 ± 25.18 | |
| UND | 2 | 13.40–14.00 | 13.70 ± 0.30 | 45.00–49.00 | 47.00 ± 2.00 | |
| Apr/17 | F | 17 | 14.10–25.40 | 18.56 ± 3.27 | 47.00–305.00 | 122.41 ± 69.03 |
| M | 7 | 13.00–17.80 | 15.37 ± 1.67 | 37.00–106.00 | 65.29 ± 22.79 | |
| Total | F | 174 | 10.40–35.40 | 19.28 ± 4.28 | 22.00–771.00 | 147.21 ± 101.50 |
| M | 140 | 9.50–24.60 | 16.63 ± 3.41 | 16.00–252.00 | 92.14 ± 53.82 | |
| UND | 17 | 10.10–18.60 | 12.66 ± 2.39 | 18.00–114.00 | 39.12 ± 23.35 | |
| Total individuals | 331 | 9.50–35.40 | 17.82 ± 4.23 | 16.00–771.00 | 118.37 ± 87.86 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, S.; Pinheiro, L.; Bentes, B. Inferences about Fishing Impacts on the Reproductive Pattern of Torroto Grunt—Genyatremus luteus (BLOCH, 1790)—From the Amazon Continental Shelf. Fishes 2023, 8, 181. https://doi.org/10.3390/fishes8040181
Fernandes S, Pinheiro L, Bentes B. Inferences about Fishing Impacts on the Reproductive Pattern of Torroto Grunt—Genyatremus luteus (BLOCH, 1790)—From the Amazon Continental Shelf. Fishes. 2023; 8(4):181. https://doi.org/10.3390/fishes8040181
Chicago/Turabian StyleFernandes, Suelly, Larissa Pinheiro, and Bianca Bentes. 2023. "Inferences about Fishing Impacts on the Reproductive Pattern of Torroto Grunt—Genyatremus luteus (BLOCH, 1790)—From the Amazon Continental Shelf" Fishes 8, no. 4: 181. https://doi.org/10.3390/fishes8040181
APA StyleFernandes, S., Pinheiro, L., & Bentes, B. (2023). Inferences about Fishing Impacts on the Reproductive Pattern of Torroto Grunt—Genyatremus luteus (BLOCH, 1790)—From the Amazon Continental Shelf. Fishes, 8(4), 181. https://doi.org/10.3390/fishes8040181