
Semi-Automated Inquiry of Fish Launch Angle and Speed for Hazard Analysis

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
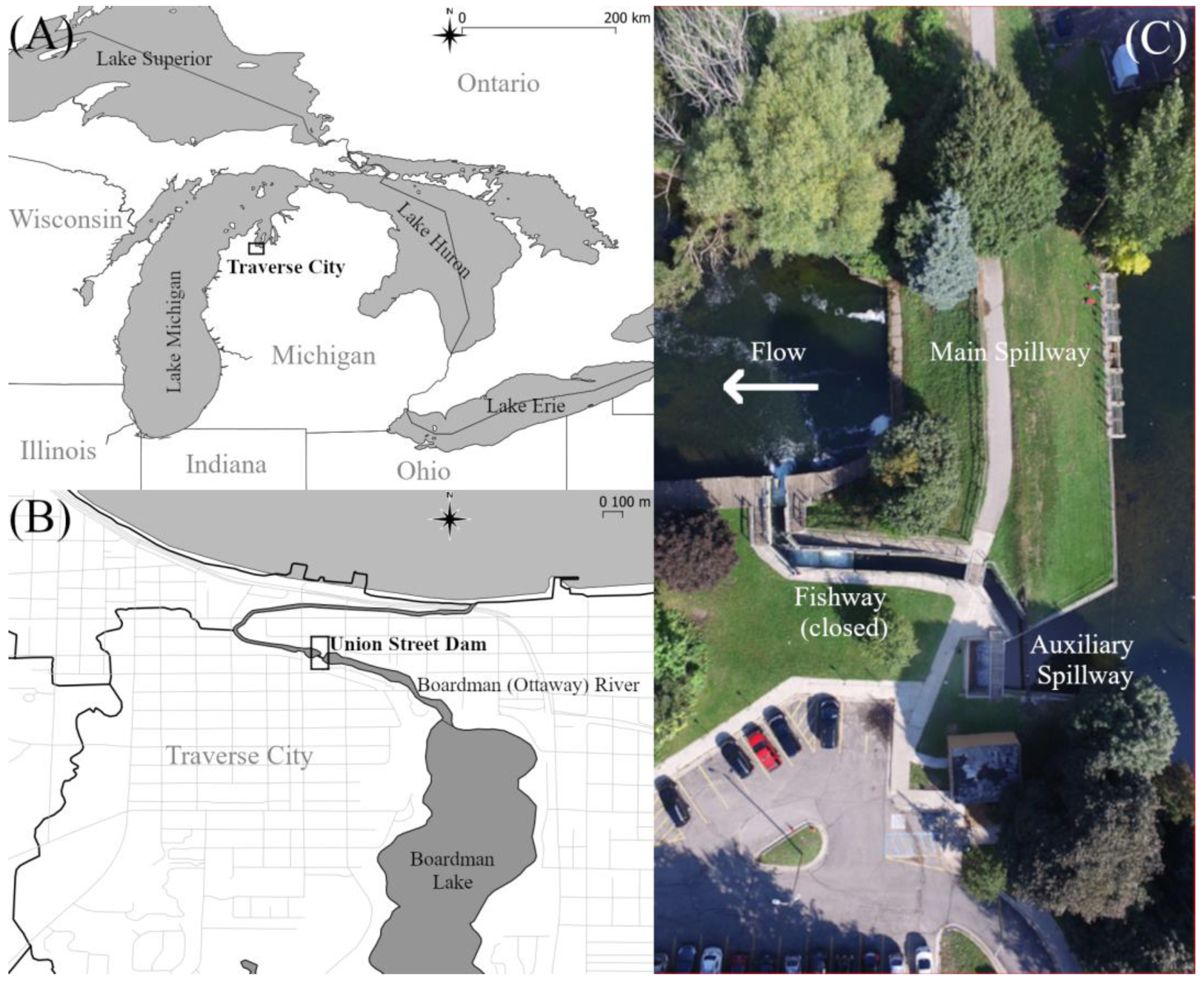
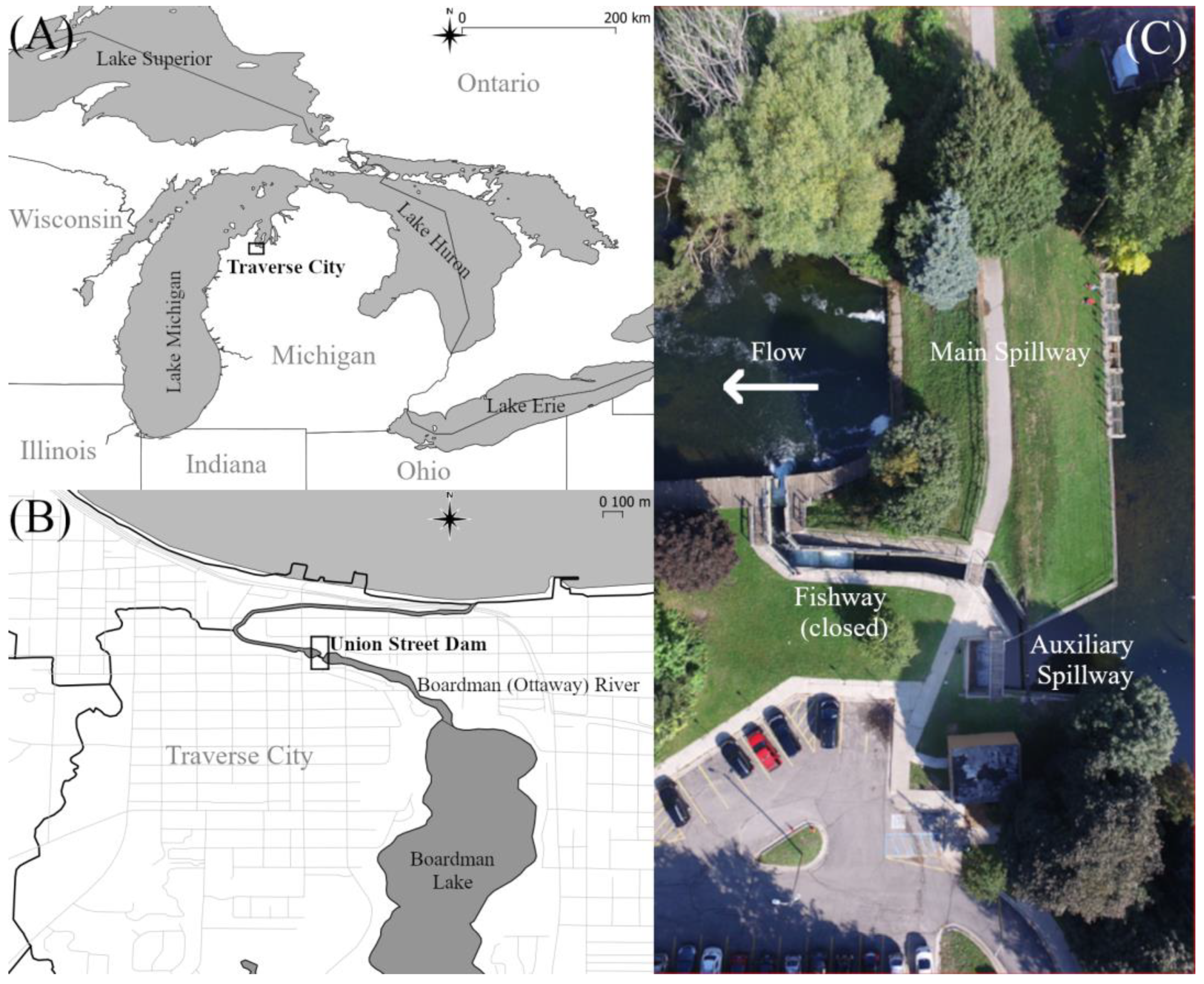
2.1. Field Site
2.2. Data Collection and Preprocessing
2.3. Image Annotation
2.4. Annotation Analysis
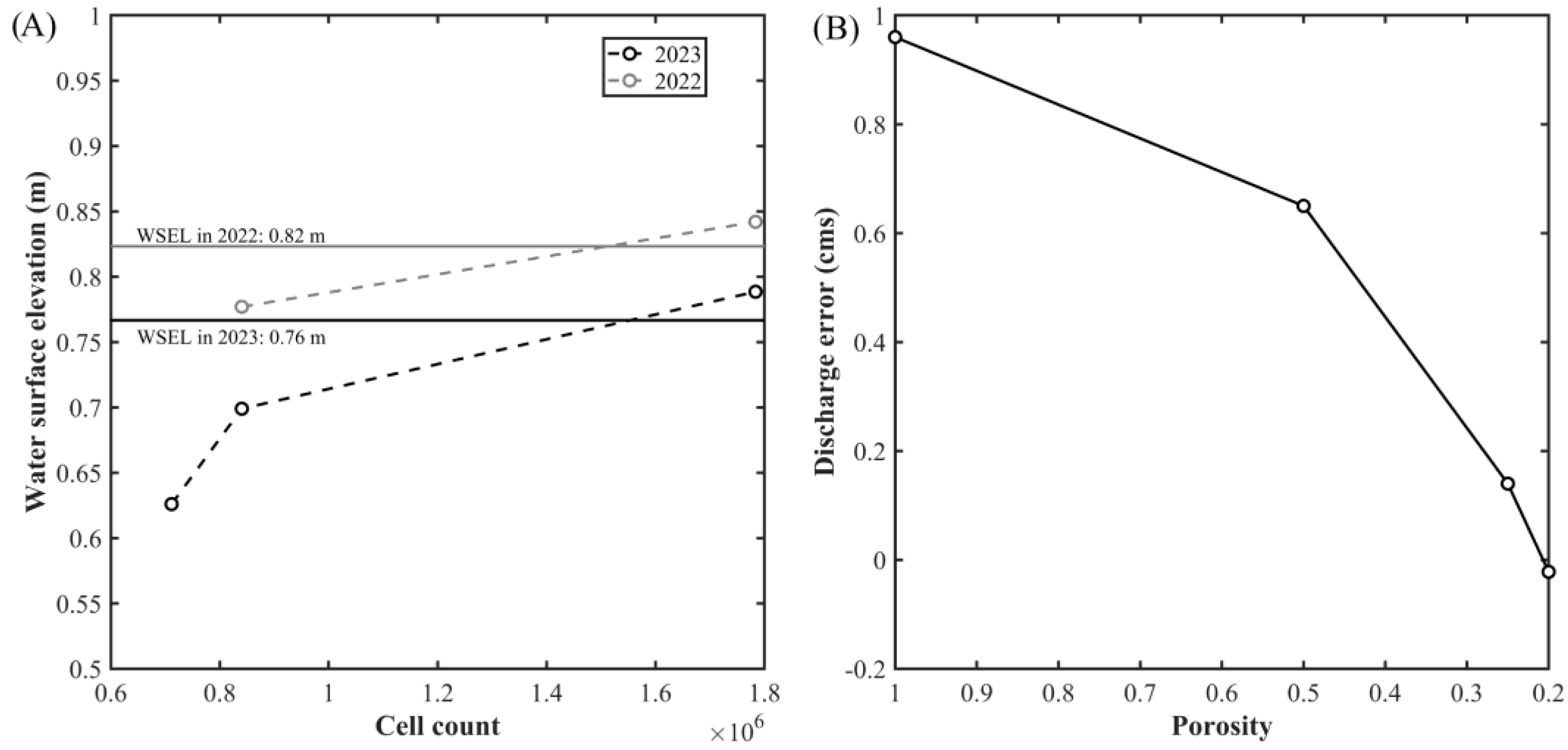
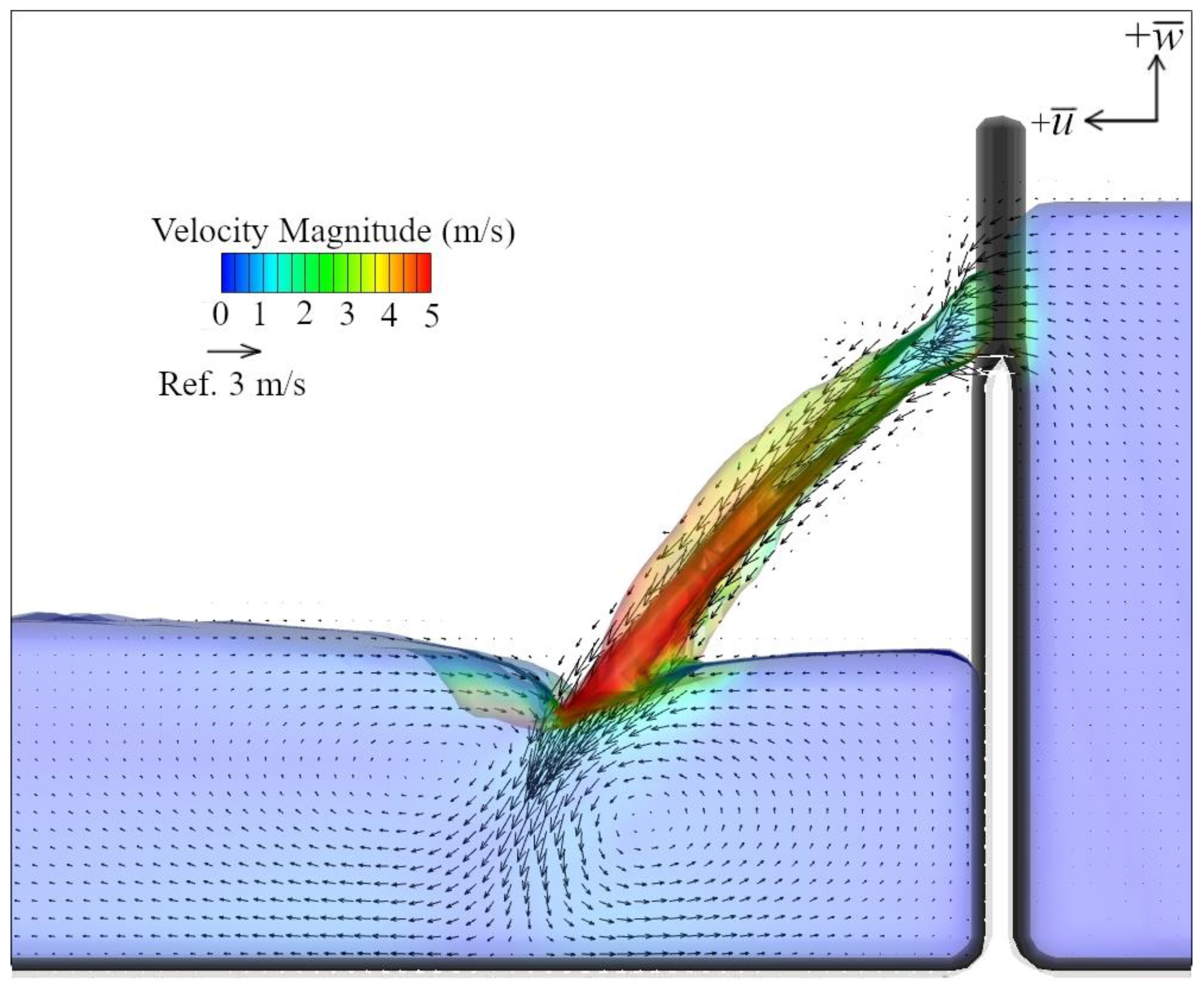
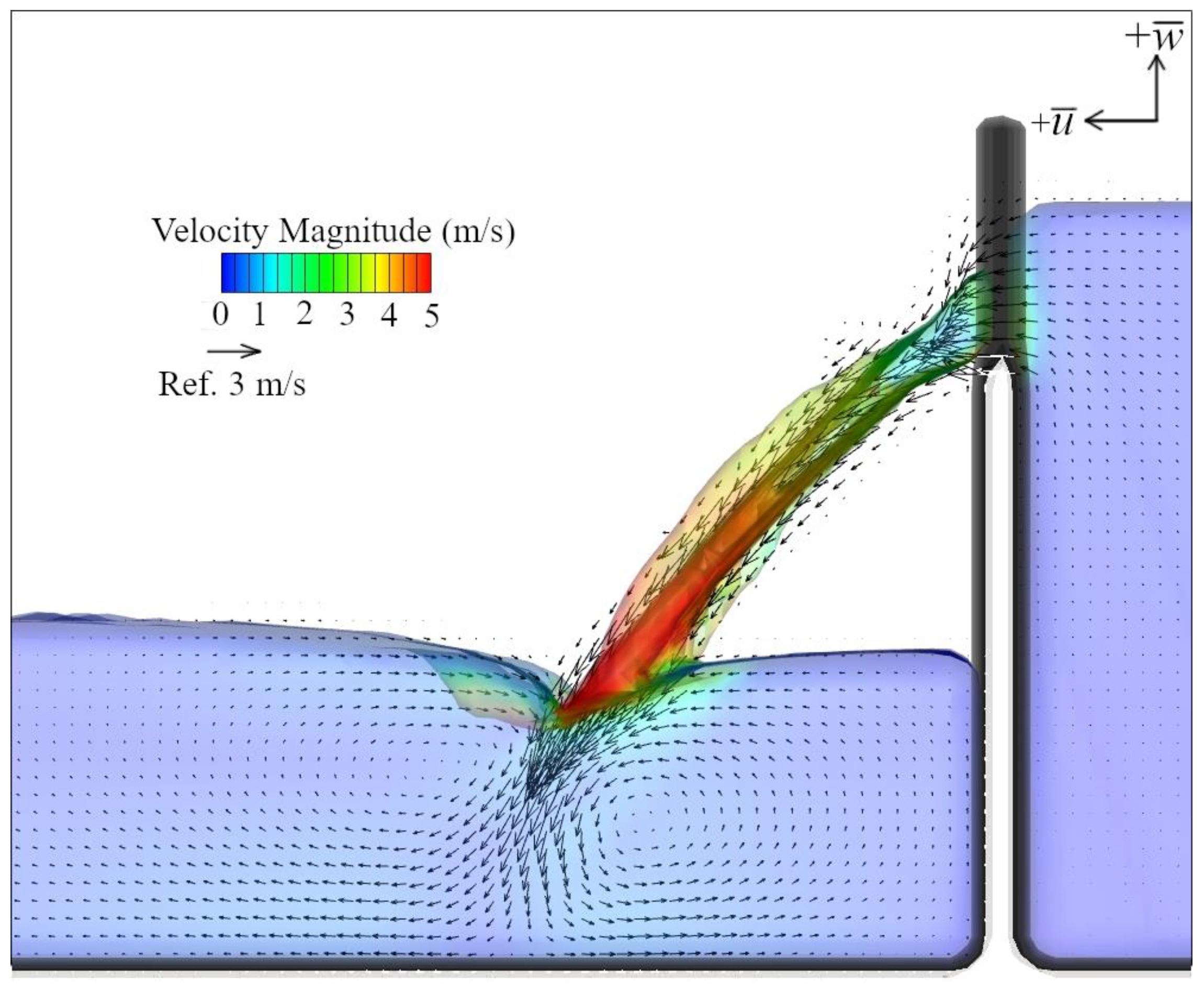
2.5. Hydraulic Modeling
2.6. Statistical Analyses
3. Results
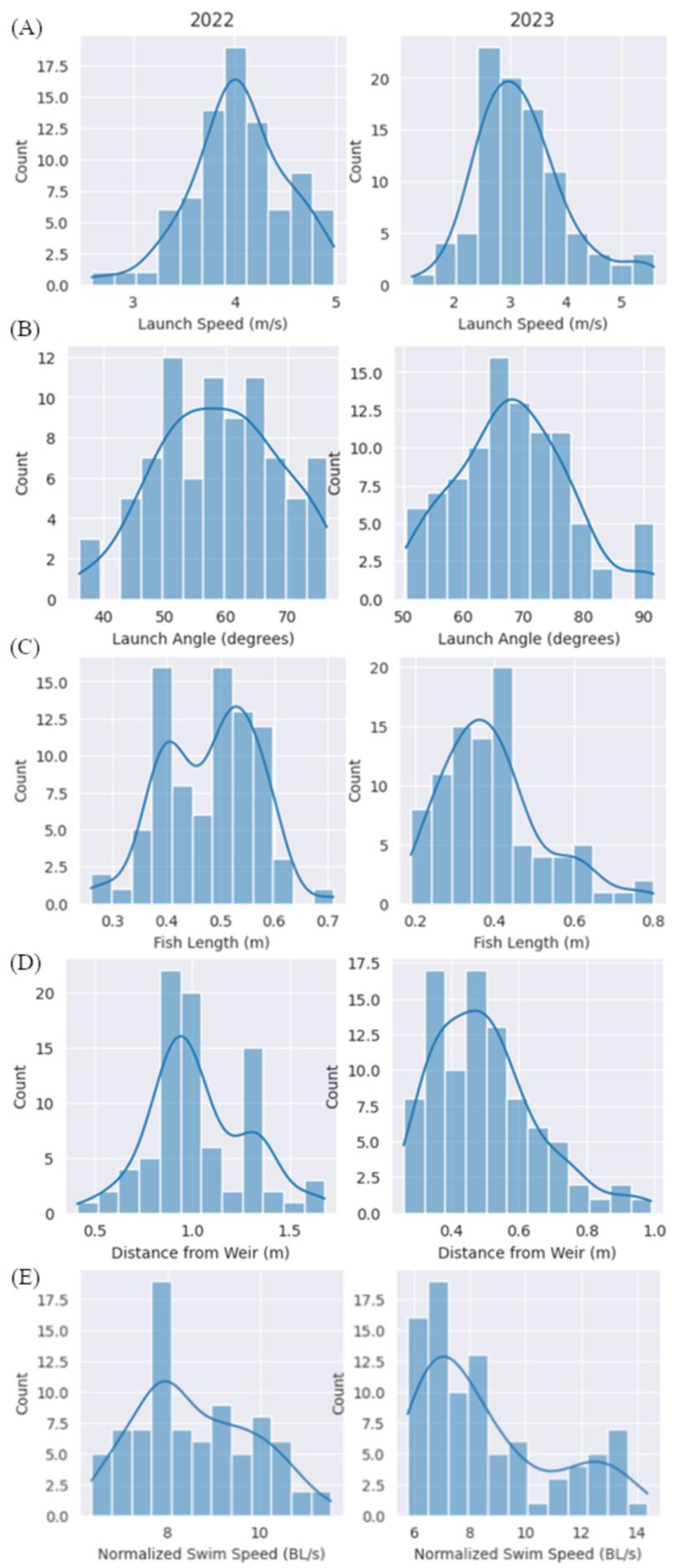
3.1. Jump Characteristics
3.2. Observer Variability
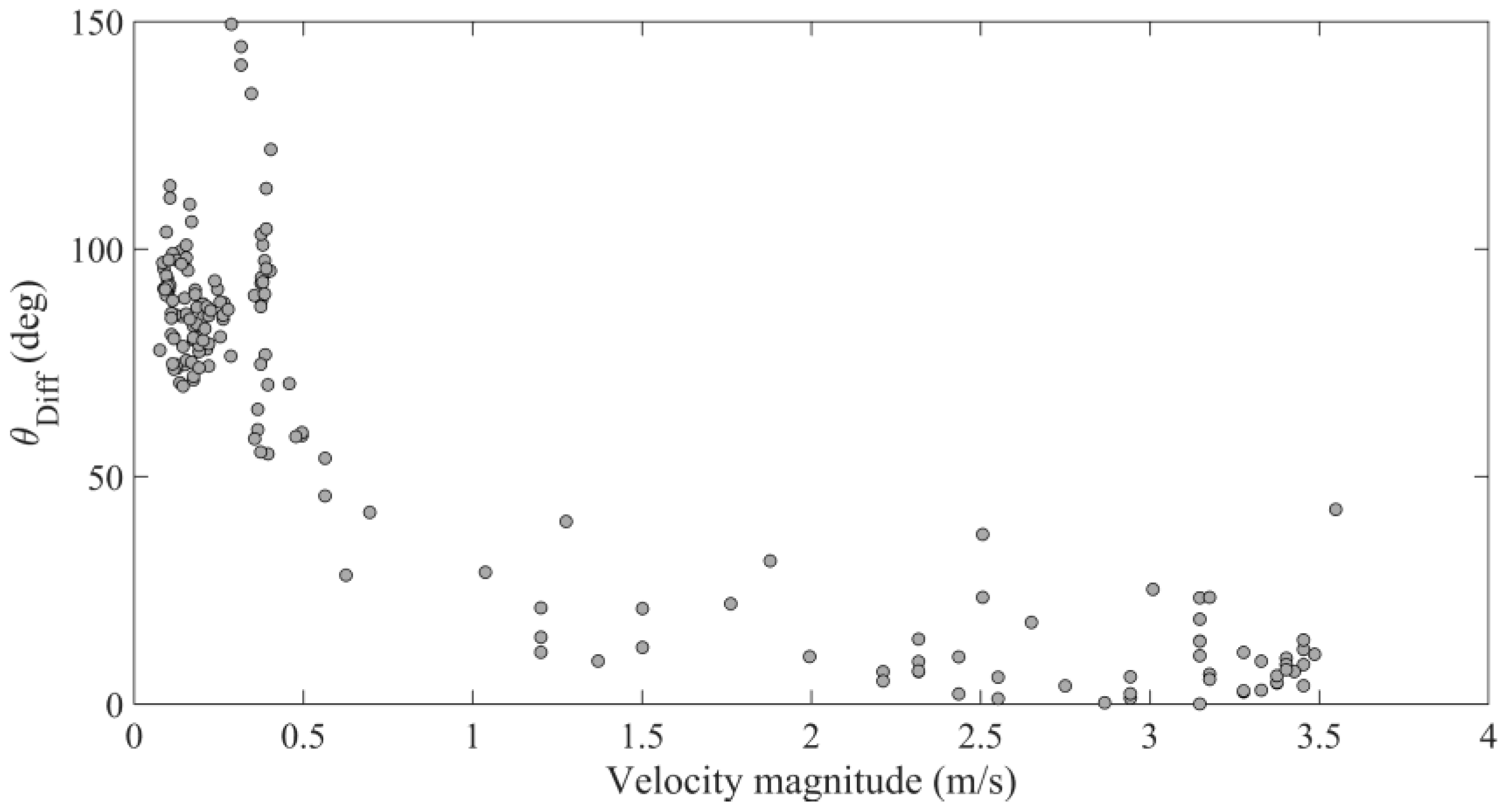
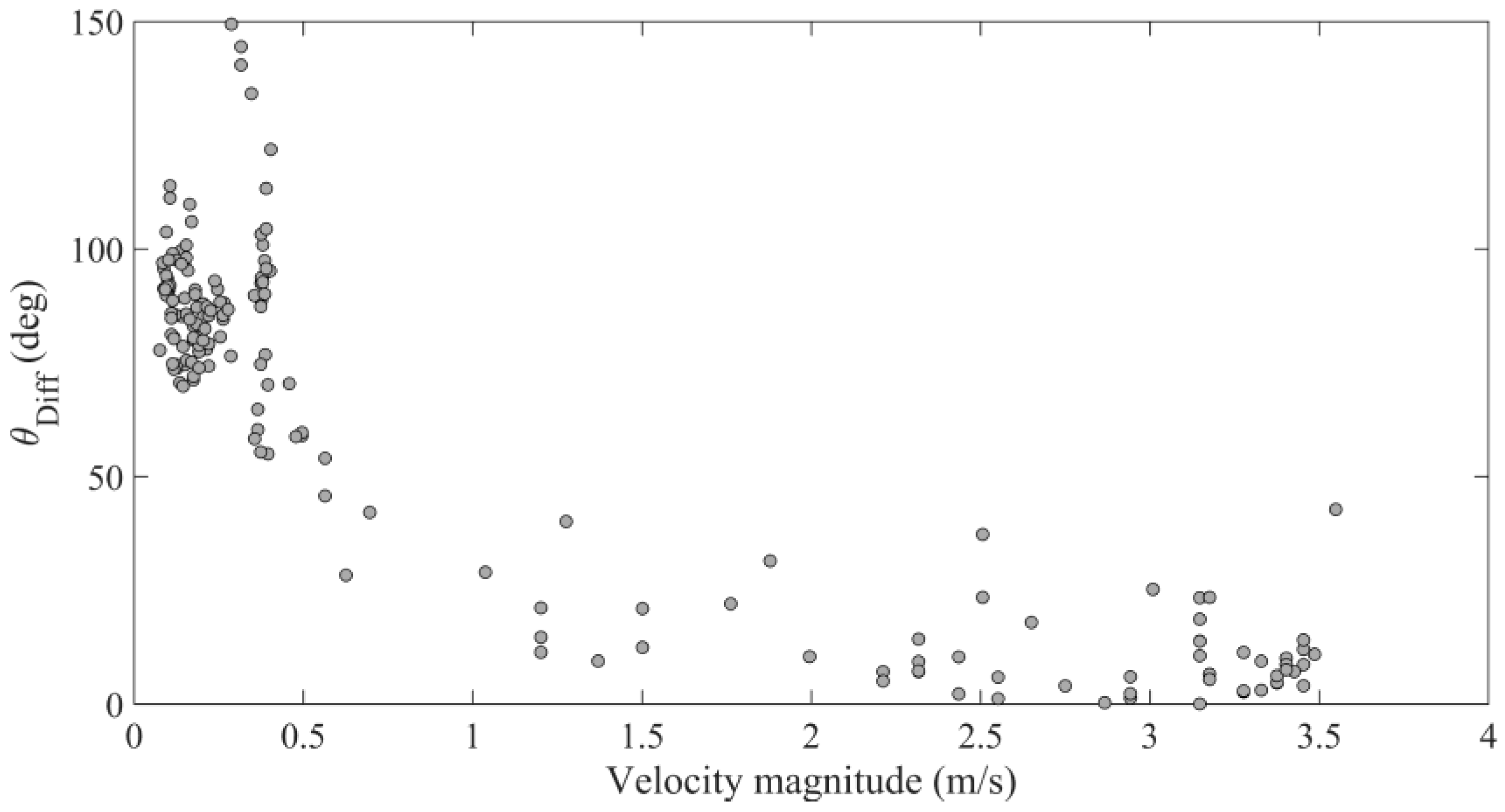
3.3. Hydraulic Conditions and Difference Angle
4. Discussion
4.1. Comparison of Jump Characteristics
4.2. Hydraulic Cues of Launch Angle
4.3. Additional Uses of Custom Annotation Application
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lovell, S.J.; Stone, S.F.; Fernandez, L. The Economic Impacts of Aquatic Invasive Species: A Review of the Literature. Agric. Resour. Econ. Rev. 2006, 35, 195–208. [Google Scholar] [CrossRef]
- Coble, D.W.; Bruesewitz, R.E.; Fratt, T.W.; Scheirer, J.W. Lake Trout, Sea Lampreys, and Overfishing in the Upper Great Lakes: A Review and Reanalysis. Trans. Am. Fish. Soc. 1990, 119, 985–995. [Google Scholar] [CrossRef]
- Fetterolf, C.M., Jr. M. Why a Great Lakes Fishery Commission and Why a Sea Lamprey International Symposium. Can. J. Fish. Aqua. Sci. 1980, 37, 1588–1593. [Google Scholar] [CrossRef]
- McLaughlin, R.L.; Smyth, E.R.; Castro-Santos, T.; Jones, M.L.; Koops, M.A.; Pratt, T.C.; Vélez-Espino, L.-A. Unintended consequences and trade-offs of fish passage. Fish Fish. 2013, 14, 580–604. [Google Scholar] [CrossRef]
- Siefkes, M.J.; Steeves, T.B.; Sullivan, W.P.; Twohey, M.B.; Li, W. Sea Lamprey Control: Past, Present, and Future. In Great Lakes Fisheries Policy and Management: A Binational Perspective, 2nd ed.; Taylor, W.W., Lynch, A.J., Leonard, N.J., Eds.; Michigan State University Press: East Lansing, MI, USA, 2013; pp. 651–704. [Google Scholar]
- Zielinski, D.P.; McLaughlin, R.; Castro-Santos, T.; Paudel, B.; Hrodey, P.; Muir, A. Alternative sea lamprey barrier technologies: History as a control tool. Rev. Fish. Sci. Aqua 2019, 27, 438–457. [Google Scholar] [CrossRef]
- Lake Michigan’s Tributary and Nearshore Fish Habitats. Available online: https://www.researchgate.net/publication/242113783_Lake_Michigan’s_tributary_and_nearshore_fish_habitats (accessed on 2 October 2022).
- Lane, J.A.; Portt, C.B.; Minns, C.K. Nursery Habitat Characteristics of Great Lakes Fishes; Can. MS Rpt. Fish. Aquati. Sci. 2338; Fisheries and Oceans Canada: Ottawa, ON, Canada, 1996; pp. 1–44. [Google Scholar]
- River Renewal: Restoring Rivers Through Hydropower Dam Relicensing|Hydropower Reform Coalition. Available online: https://hydroreform.org/resource/river-renewal-restoring-rivers-through-hydropower-dam-relicensing/ (accessed on 3 April 2023).
- Williams, J.G.; Amstrong, G.; Katopodis, C.; Larinier, M.; Travade, F. Thinking like a fish: A key ingredient for development of effective fish passage facilities at river obstructions. River Res. App. 2012, 28, 407–417. [Google Scholar] [CrossRef]
- Mallen-Cooper, M.; Brand, D.A. Non-salmonids in a salmonid fishway: What do 50 years of data tell us about past and future fish passage? Fish. Manag. Ecol. 2007, 14, 319–332. [Google Scholar] [CrossRef]
- Bunt, C.M.; Castro-Santos, T.; Haro, A. Reinforcement and validation of the analyses and conclusions related to fishway evaluation data from Bunt et al.:‘Performance of fish passage structures at upstream barriers to migration’. River Res. App. 2016, 32, 2125–2137. [Google Scholar]
- Clay, C.H.; Eng, P. Design of Fishways and Other Fish Facilities, 1st ed.; CRC Press: Boca Raton, FL, USA, 2017; pp. 1–247. [Google Scholar]
- Buysse, D.; Coeck, J.; Maes, J. Potential re-establishment of diadromous fish species in the River Scheldt (Belgium). Hydrobiologia 2008, 602, 155–159. [Google Scholar] [CrossRef]
- Kareiva, P.; Marvier, M.; McClure, M. Recovery and management options for spring/summer chinook salmon in the Columbia River basin. Science 2000, 290, 977–979. [Google Scholar] [CrossRef]
- Lauritzen, D.V.; Hertel, F.S.; Jordan, L.K.; Gordon, M.S. Salmon jumping: Behavior, kinematics and optimal conditions, with possible implications for fish passageway design. Bioinspir. Biomim. 2010, 5, 035006. [Google Scholar] [CrossRef]
- Lundqvist, H.; Rivinoja, P.; Leonardsson, K.; McKinnell, S. Upstream passage problems for wild Atlantic salmon (Salmo salar L.) in a regulated river and its effect on the population. Hydrobiologia 2008, 602, 111–127. [Google Scholar] [CrossRef]
- Stuart, I.G.; Marsden, T.J. Evaluation of cone fishways to facilitate passage of small-bodied fish. Aquac. Fish. 2019, 6, 125–134. [Google Scholar] [CrossRef]
- Silva, A.T.; Lucas, M.C.; Castro-Santos, T.; Katopodis, C.; Baumgartner, L.J.; Thiem, J.D.; Cooke, S.J. The future of fish passage science, engineering, and practice. Fish Fish. 2018, 19, 340–362. [Google Scholar] [CrossRef]
- Zielinski, D.P.; McLaughlin, R.L.; Pratt, T.C.; Goodwin, R.A.; Muir, A.M. Single-stream recycling inspires selective fish passage solutions for the connectivity conundrum in aquatic ecosystems. Bioscince 2020, 70, 871–886. [Google Scholar] [CrossRef] [PubMed]
- Benoit, D.M.; Zielinski, D.P.; Swanson, R.G.; McLaughlin, R.L.; Castro-Santos, T.R.; Goodwin, R.A.; Pratt, T.C.; Muir, A.M. FishPass sortable attribute database: Phenological, morphological, physiological, and behavioral characteristics related to passage and movement of Laurentian Great Lakes fishes. J. Great Lakes Res. 2023. [Google Scholar] [CrossRef]
- Zorn, T.G.; Hessenauer, J.M.; Wills, T.C. Increasing connectivity of Great Lakes tributaries: Interspecific and intraspecific effects on resident brook trout and brown trout populations. Ecol. Freshw. Fish. 2019, 29, 519–532. [Google Scholar] [CrossRef]
- Reiser, D.W.; Huang, C.M.; Beck, S.; Gagner, M.; Jeanes, E. Defining flow windows for upstream passage of adult anadromous salmonids at cascades and falls. Trans. Am. Fish. Soc. 2006, 135, 668–679. [Google Scholar] [CrossRef]
- Powers, P.D.; Osborn, J.F. New Concepts in Fish Ladder Design: Analysis of Barrier to Upstream Fish Migration, Volume IV of IV; Investigation of the Physical and Biological Conditions Affecting Fish Passage Success at Culverts and Waterfalls. 1982-1984 Final Report, Contract DE-A179-82BP36523, Project 82–14, USA. 1985. Available online: http://www.efw.bpa.gov/Publications/U36523–1.pdf (accessed on 18 December 2018).
- Zielinski, D.P.; Freiburger, C. Advances in fish passage in the Great Lakes basin. J. Great Lakes Res. 2021, 47, S439–S447. [Google Scholar] [CrossRef]
- Scott, W.B.; Crossman, E.J. Freshwater Fishes of Canada. Fish. Res. Board Can. Bull. 1973, 1, 1–966. [Google Scholar]
- Biette, R.M.; Dodge, D.P.; Hassinger, R.L.; Stauffer, T.M. Life History and Timing of Migrations and Spawning Behavior of Rainbow Trout (Salmo gairdneri) Populations of the Great Lakes. Can. J. Fish. Aqua. Sci. 1981, 38, 1759–1771. [Google Scholar] [CrossRef]
- Goodyear, C.D.; Edsall, T.A.; Dempsey, D.O.; Moss, G.D.; Polanski, P.E. Atlas of the Spawning and Nursery Areas of Great Lakes Fishes; Volume VIII; FWS/OBS-82/52; U.S. Fish and Wildlife Service: Washington, DC, USA, 1982; pp. 1–144. [Google Scholar]
- Weaver, C. Influence of water velocity upon orientation and performance of adult migrating salmonids. U.S. Fish Wildl. Serv. Fish. Bull. 1963, 63, 97–121. [Google Scholar]
- Katopodis, C.; Gervais, R. Fish Swimming Performance Database and Analyses; Research Document 2016/002; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2016; p. 550. [Google Scholar]
- Webb, P.W. Temperature effects on acceleration of rainbow trout, Salmo gairdneri. J. Fish. Res. Board. Can. 1978, 35, 1417–1422. [Google Scholar] [CrossRef]
- Lauritzen, D.V.; Hertel, F.S.; Gordon, M.S. A kinematic examination of wild sockeye salmon jumping up natural waterfalls. J. Fish. Biol. 2005, 67, 1010–1020. [Google Scholar] [CrossRef]
- Stuart, T.A. The leaping behaviour of salmon and trout at falls and obstructions. Freshw. Salmon Fish. Res. 1962, 28, 1–46. [Google Scholar]
- Aaeserude, R.G.; Osborn, J.F. New Concepts in Fish Ladder Design, Volume II of IV.; Results of Laboratory and Field Research on New Concepts in Weir and Pool Fishways. 1982–1984 Final Report, Contract DE-AI79-82BP36523, Project 82-14, USA. 1985. Available online: https://www.osti.gov/servlets/purl/5965536 (accessed on 18 December 2018).
- Morán-López, R.; Uceda-Tolosa, Ó. Biomechanics of fish swimming and leaping under waterfalls: A realistic field, image-based biophysical model with bioengineering implications. Bioinspir. Biomim. 2020, 15, 056011. [Google Scholar] [CrossRef] [PubMed]
- VLC: Official Site—Free Multimedia Solutions for all OS!—VideoLAN. Available online: https://www.videolan.org/ (accessed on 4 January 2023).
- FLOW-3D. FLOW-3D User Manual. Flow Science Inc. Available online: https://flow3d.sharefile.com/share/view/scc68be45df33406698c1b39eee7d3b14 (accessed on 14 September 2023).
- Pope, S.B. Turbulent Flows; Cambridge University Press: Cambridge, UK, 2000; p. 771. [Google Scholar]
- Ho, J.; Coonrod, J.; Hanna, L.J.; Mefford, B.W. Hydrodynamic modelling study of a fish exclusion system for a river diversion. River Res. Applic. 2011, 27, 184–192. [Google Scholar] [CrossRef]
- Box, G.E.P.; Cox, D.R. An analysis of transformations. J. Roy. Stat. Soc. Ser B 1964, 26, 211–252. [Google Scholar] [CrossRef]
- Silva, A.T.; Hatry, C.; Thiem, J.D.; Gutowsky, L.F.G.; Hatin, D.; Zhu, D.Z.; Dawson, J.W.; Katopodis, C.; Cooke, S.J. Behaviour and locomotor activity of a migratory catostomid during fishway passage. PLoS ONE 2015, 10, 0123051. [Google Scholar] [CrossRef]
- Tudorache, C.; O’Keefe, R.A.; Benfey, T.J. Flume length and post-exercise impingement affect anaerobic metabolism in brook charr Salvelinus fontinalis. J. Fish Biol. 2010, 76, 729–733. [Google Scholar] [CrossRef]
- Tudorache, C.; Viaenen, P.; Blust, R.; De Boeck, G. Longer flumes increase critical swimming speeds by increasing burst and glide swimming duration in carp (Cyprinus carpio L.). J. Fish Biol. 2007, 71, 1630–1638. [Google Scholar] [CrossRef]
- Goodwin, R.A.; Lai, Y.G.; Taflin, D.E.; Smith, D.L.; McQuirk, J.; Trang, R.; Reeves, R. Predicting near-term, out-of-sample fish passage, guidance, and movement across diverse river environments by cognitively relating momentary behavioral decisions to multiscale memories of past hydrodynamic experiences. Front. Ecol. Evol. 2023, 11, 703946. [Google Scholar] [CrossRef]
- Bravata, N.; Kelly, D.; Eickholt, J.; Bryan, J.; Miehls, S.; Zielinski, D. Applications of deep convolutional neural networks to predict length, circumference, and weight from mostly dewatered images of fish. Ecol. Evol. 2020, 10, 9313–9325. [Google Scholar] [CrossRef] [PubMed]
- Eickholt, J.; Kelly, D.; Bryan, J.; Miehls, S.; Zielinski, D. Advancements towards selective barrier passage by automatic species identification: Applications of deep convolutional neural networks on images of dewatered fish. ICES J. Mar. Sci. 2020, 77, 2804–2813. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Field | Sample Value for Field |
|---|---|
| Video file ID | 63efd4cb5e1e50998c52e672 |
| Start frame ID | 63efd5385e1e50998c537129 |
| Coordinate1 | (747,164) |
| Coordinate2 | (770,214) |
| Coordinate3 | (791,263) |
| Coordinate4 | (818,307) |
| Calibration1 | (836,361) |
| Calibration2 | (836,469) |
| Calibration3 | (836,582) |
| Calibration4 | (838,690) |
| Nose | (845,346) |
| Tail | (720,160) |
| Depth | 35 |
| Angle offset | 0.12 |
| Launch angle | 65.41 |
| Launch speed | 3.76 |
| Length | 0.51 |
| Annotated by | PN |
| Start Frame | BoardmanSpillway-2022-04-15-16h34m04s-GP102357-025.png |
| Video file name | BoardmanSpillway-2022-04-15-16h34m04s |
| Horizontal edge | 883,381 |
| Vertical edge | 1,033,717 |
| Visible weir coordinates | 1,363,119 |
| Distance from the wall | 1.3 |
| Date | US WSEL (m) | DS WSEL (m) | Calculated Discharge (cm) |
|---|---|---|---|
| 15 April 2022 | 179.82 | 176.4 | 0.85 |
| 14 April 2023 | 179.79 | 176.4 | 0.78 |
| Launch Speed | |||||
|---|---|---|---|---|---|
| Year | Absolute (m/s) | Normalized (BL/s) | Launch Angle (°) | Fish Length (m) | Distance from Crest (m) |
| 2022 | 4.05 ± 0.46 | 8.60 ± 1.25 | 58.73 ± 9.88 | 0.48 ± 0.09 | 1.03 ± 0.25 |
| 2023 | 3.24 ± 0.81 | 8.61 ± 2.42 | 68.2 ± 9.5 | 0.40 ± 0.13 | 0.50 ± 0.15 |
| Model | AIC | ΔAIC |
|---|---|---|
| VM + (Year) | 101.17 | - |
| VM + LS + (Year) | 102.48 | 1.31 |
| LS + (Year) | 320.42 | 219.25 |
| Model Term | Coefficient | SE | df | t | p-Value |
|---|---|---|---|---|---|
| Intercept | 0.549 | 0.032 | 171 | 17.4 | <<0.05 |
| Velocity Magnitude | −0.524 | 0.020 | 171 | −27.0 | <<0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nallamothu, P.; Gregory, J.; Leh, J.; Zielinski, D.P.; Eickholt, J.L. Semi-Automated Inquiry of Fish Launch Angle and Speed for Hazard Analysis. Fishes 2023, 8, 476. https://doi.org/10.3390/fishes8100476
Nallamothu P, Gregory J, Leh J, Zielinski DP, Eickholt JL. Semi-Automated Inquiry of Fish Launch Angle and Speed for Hazard Analysis. Fishes. 2023; 8(10):476. https://doi.org/10.3390/fishes8100476
Chicago/Turabian StyleNallamothu, Prathyush, Jonathan Gregory, Jordan Leh, Daniel P. Zielinski, and Jesse L. Eickholt. 2023. "Semi-Automated Inquiry of Fish Launch Angle and Speed for Hazard Analysis" Fishes 8, no. 10: 476. https://doi.org/10.3390/fishes8100476
APA StyleNallamothu, P., Gregory, J., Leh, J., Zielinski, D. P., & Eickholt, J. L. (2023). Semi-Automated Inquiry of Fish Launch Angle and Speed for Hazard Analysis. Fishes, 8(10), 476. https://doi.org/10.3390/fishes8100476