From Transient to Sedentary? Changes in the Home Range Size and Environmental Patterns of Movements of European Eels (Anguilla anguilla) in a Mediterranean River


Abstract
:1. Introduction
2. Results
2.1. Effects of Passive Integrated Transponder Tags and Tag Retention
2.2. Movements and Home Range
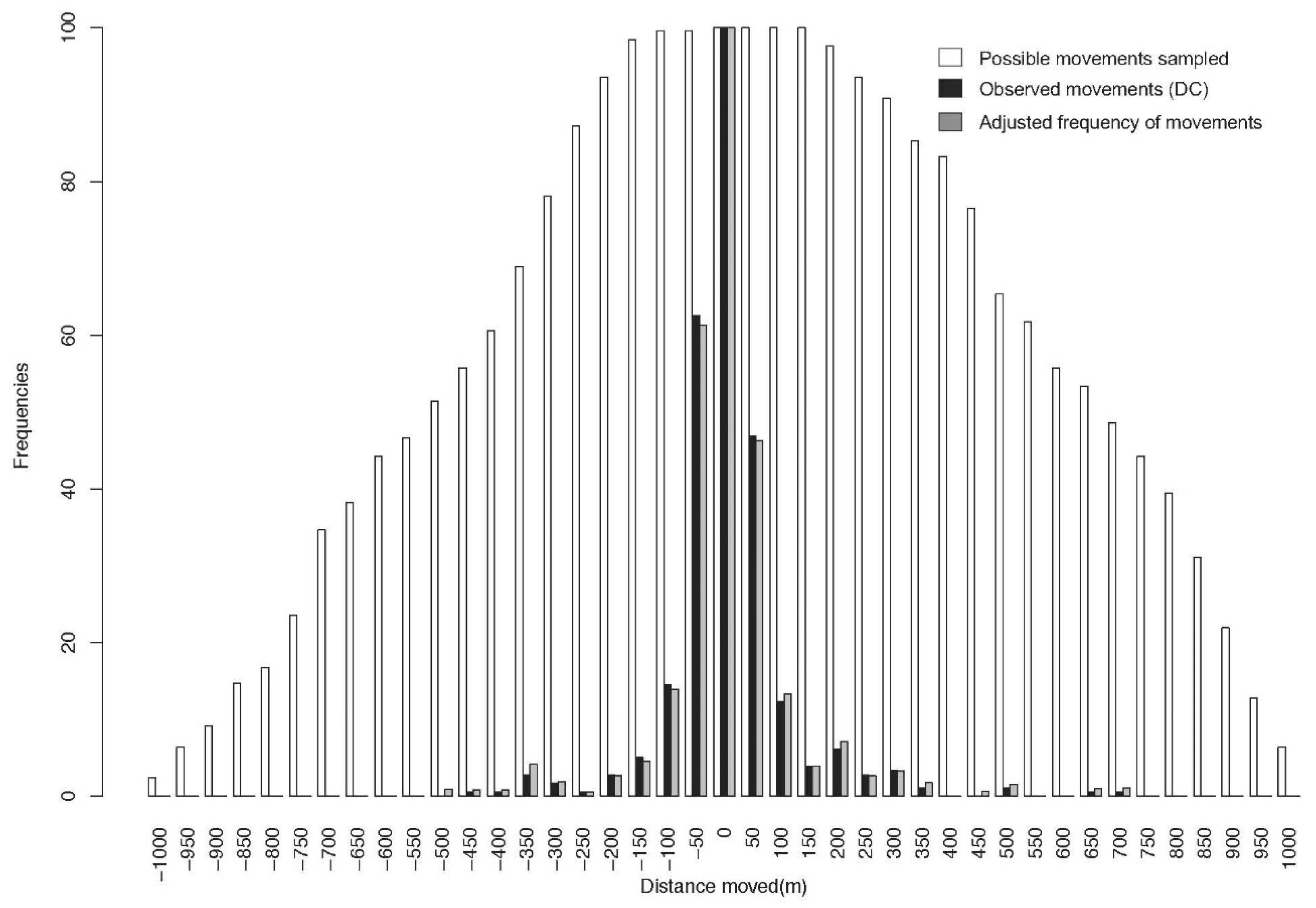
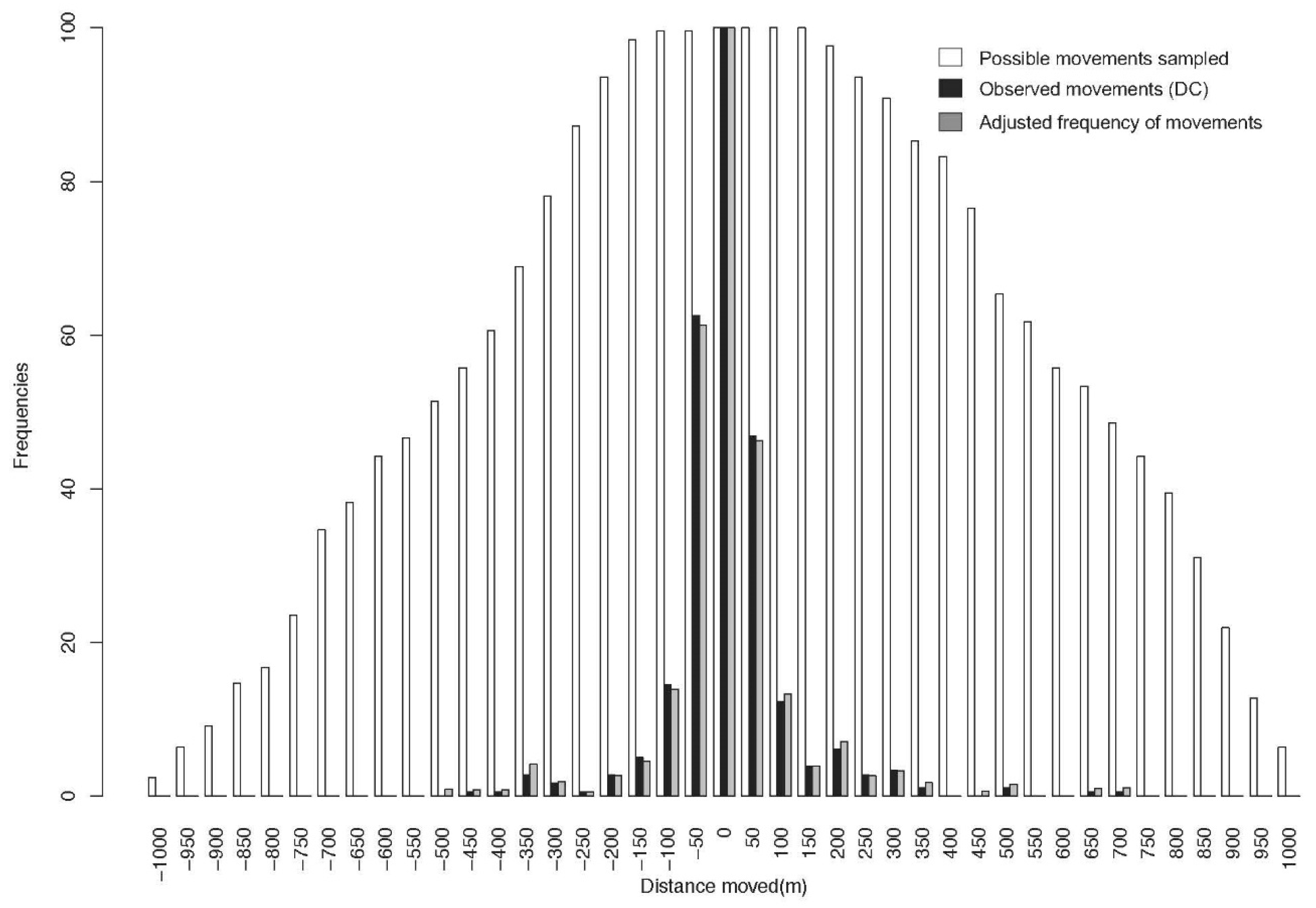
2.2.1. Deviation Estimate of Observed Movements
2.2.2. Distance between Captures (DC, Observed Movements)
2.2.3. Maximum Distance Moved (MDM, Home Range)
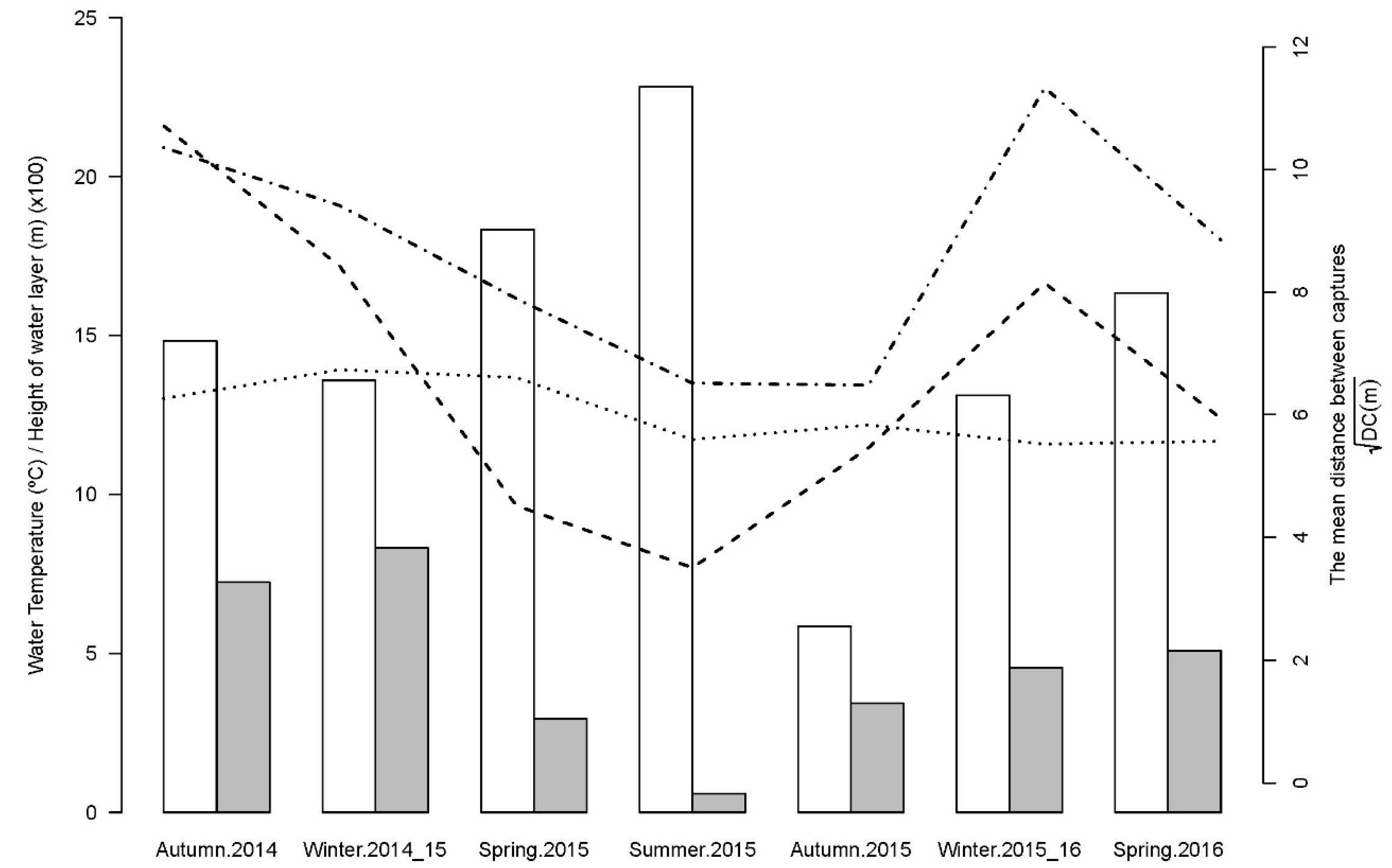
2.2.4. Factors Influencing the Extension of Eel Movements
3. Discussion
4. Material and Methods
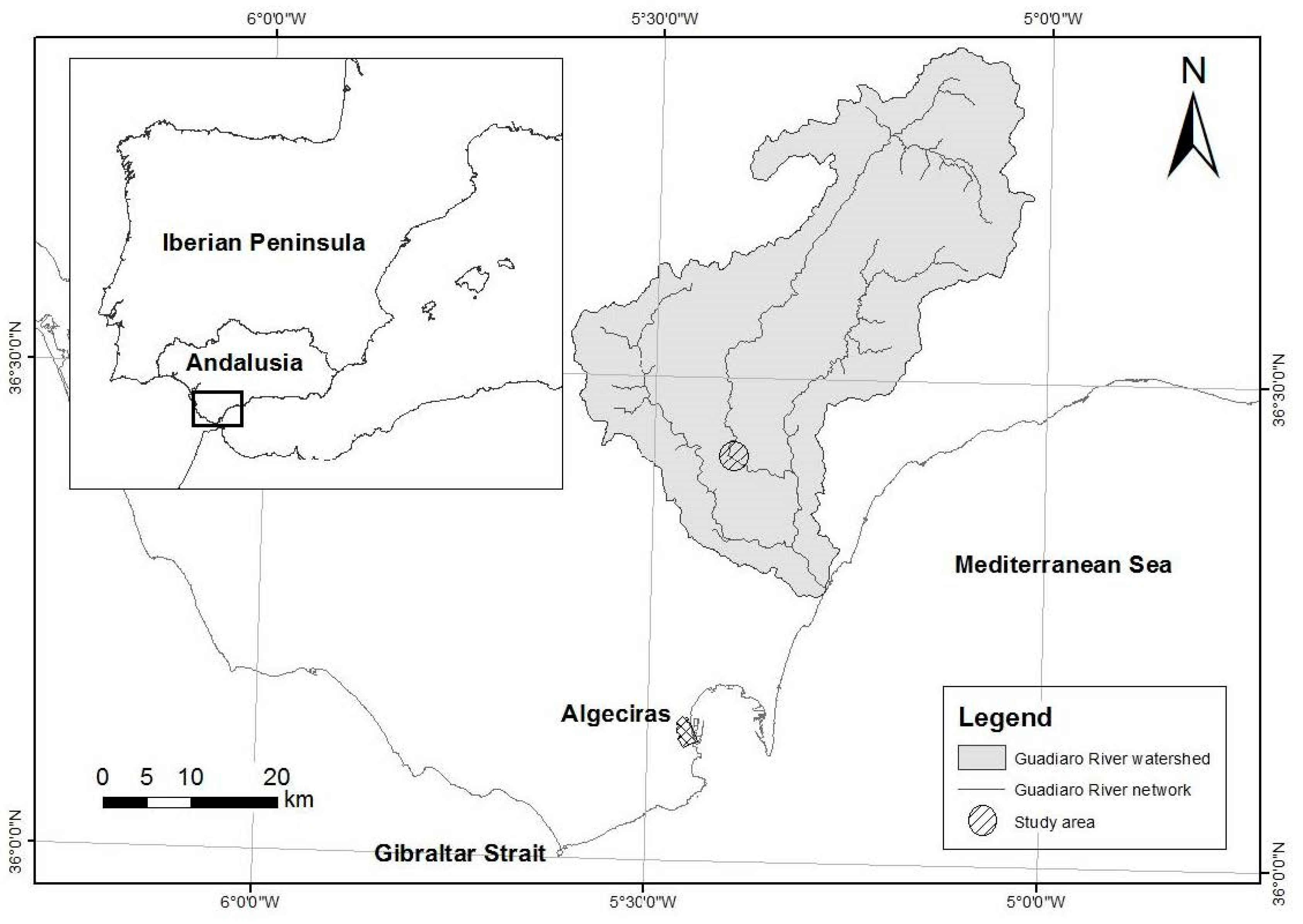
4.1. Study Area
4.2. Mark–Recapture Study
4.2.1. Tag Retention
4.2.2. Sampling Protocol
4.2.3. Data Analysis
- (1)
- DC (m; Distance between Captures or observed movements): distance moved in meters between two consecutive captures. It can be negative (downstream) or positive (upstream). DC was tested against the value of D between two consecutive captures;
- (2)
- (1)
- The TL (cm) of the individual when movement was detected;
- (2)
- CPUE (Catch per unit effort): the average number of specimens captured in a 24-h cycle, interpreted as an indicator of the degree of activity in eels.
- (1)
- River Flow (RF, m): mean river level (height of the water layer in meters) from four days before the beginning of sampling until its completion;
- (2)
- Increase in RF (ΔRF, m): the difference between the maximum and minimum water levels in the four days prior to the beginning of sampling;
- (3)
- Water Temperature (WT, °C): measured early in the morning (at around 09:00) on each day of sampling.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- ICES. Report of the Joint EIFAAC/ICES/GFCM Working Group on Eel, 3–7 November 2014, Rome, Italy; ICES CM 2014/ACOM:18; ICES: Copenhagen, Denmark, 2014; p. 203. [Google Scholar]
- ICES. Report of the Joint EIFAAC/ICES/GFCM Working Group on Eel (WGEEL), 24 November–2 December 2015, Antalya, Turkey; ICES CM 2015/ACOM:18; ICES: Copenhagen, Denmark, 2015; p. 130. [Google Scholar]
- ICES. Report of the Working Group on Eels (WGEEL), 15–22 September 2016, Cordoba, Spain; ICES CM 2016/ACOM:19; ICES: Copenhagen, Denmark, 2016; p. 107. [Google Scholar]
- Hill, J.; Grossman, G.D. Home Range Estimates for Three North American Stream Fishes. Copeia 1987, 1987, 376–380. [Google Scholar] [CrossRef]
- Belica, L.A.T.; Rahel, F.J. Movements of creek chubs, Semotilus atromaculatus, among habitat patches in a plains stream. Ecol. Freshw. Fish 2008, 17, 258–272. [Google Scholar] [CrossRef]
- Smithson, E.B.; Johnston, C.E. Movement patterns of stream fishes in an Ouachita highlands stream: An examination of the restricted movement paradigm. Trans. Am. Fish. Soc. 1999, 128, 847–853. [Google Scholar] [CrossRef]
- Skalski, G.T.; Gilliam, J.F. Modeling Diffusive Spread in a Heterogeneous Population: A Movement Study with Stream Fish. Ecology 2000, 81, 1685–1700. [Google Scholar] [CrossRef]
- Feunteun, E.; Laffaille, P.; Robinet, T.; Briand, C.; Baisez, A.; Olivier, J.-M.; Acou, A. A Review of Upstream Migration and Movements in Inland Waters by Anguillid Eels: Toward a General Theory. In Eel Biology; Aida, K., Tsukamoto, K., Yamauchi, K., Eds.; Springer: Tokyo, Japan, 2003; pp. 191–213. [Google Scholar]
- Imbert, H.; LaBonne, J.; Rigaud, C.; Lambert, P. Resident and migratory tactics in freshwater European eels are size-dependent. Freshw. Biol. 2010, 55, 1483–1493. [Google Scholar] [CrossRef]
- Hedger, R.D.; Dodson, J.J.; Hatin, D.; Caron, F.; Fournier, D.; Hedger, R. River and estuary movements of yellow-stage American eels Anguilla rostrata, using a hydrophone array. J. Fish Biol. 2010, 76, 1294–1311. [Google Scholar] [CrossRef] [PubMed]
- Ovidio, M.; Seredynski, A.L.; Philippart, J.-C.; Matondo, B.N. A bit of quiet between the migrations: The resting life of the European eel during their freshwater growth phase in a small stream. Aquat. Ecol. 2013, 47, 291–301. [Google Scholar] [CrossRef]
- Béguer-Pon, M.; Castonguay, M.; Benchetrit, J.; Hatin, D.; Legault, M.; Verreault, G.; Dodson, J.J. Large-scale, seasonal habitat use and movements of yellow American eels in the St. Lawrence River revealed by acoustic telemetry. Ecol. Freshw. Fish 2015, 24, 99–111. [Google Scholar] [CrossRef]
- Barry, J.; Newton, M.; Dodd, J.A.; Hooker, O.E.; Boylan, P.; Lucas, M.C.; Adams, C.E. Foraging specialisms influence space use and movement patterns of the European eel Anguilla anguilla. Hydrobiologia 2016, 766, 333–348. [Google Scholar] [CrossRef]
- Porter, J.H.; Dooley, J.L. Animal Dispersal Patterns: A Reassessment of Simple Mathematical Models. Ecology 1993, 74, 2436–2443. [Google Scholar] [CrossRef]
- Riley, W.D.; Walker, A.M.; Bendall, B.; Ives, M.J. Movements of the European eel (Anguilla anguilla) in a chalk stream. Ecol. Freshw. Fish 2011, 20, 628–635. [Google Scholar] [CrossRef]
- Walker, A.M.; Godard, M.J.; Davison, P. The home range and behaviour of yellow-stage European eel Anguilla anguilla in an estuarine environment. Aquat. Conserv. 2014, 24, 155–165. [Google Scholar] [CrossRef]
- Baras, E.; Jeandrain, D.; Serouge, B.; Philippart, J.C. Seasonal variations of time and space utilisation by radiotagged yellow eels Anguilla anguilla (L.) in a small stream. Hydrobiologia 1998, 371, 187–198. [Google Scholar] [CrossRef]
- Le Pichon, C.; Coustillas, J.; Zahm, A.; Bunel, M.; Gazeau-Nadin, C.; Rochard, E. Summer use of the tidal freshwaters of the River Seine by three estuarine fish: Coupling telemetry and GIS spatial analysis. Estuar. Coast. Shelf Sci. 2017, 196, 83–96. [Google Scholar] [CrossRef]
- Verhelst, P.; Reubens, J.; Pauwels, I.; Buysse, D.; Aelterman, B.; Van Hoey, S.; Goethals, P.; Moens, T.; Coeck, J.; Mouton, A. Movement behaviour of large female yellow European eel (Anguilla anguilla L.) in a freshwater polder area. Ecol. Freshw. Fish 2018, 27, 471–480. [Google Scholar] [CrossRef]
- Thibault, I.; Dodson, J.J.; Caron, F. Yellow-stage American eel movements determined by microtagging and acoustic telemetry in the St Jean River watershed, Gaspé, Quebec, Canada. J. Fish Biol. 2007, 71, 1095–1111. [Google Scholar] [CrossRef]
- LaBar, G.W.; Hernando Casal, J.A.; Fernández Delgado, C. Local movements and population size of European eels, Anguilla anguilla, in a small lake in south-western Spain. Environ. Biol. Fishes 1987, 19, 111–117. [Google Scholar] [CrossRef]
- Sinha, V.R.P.; Jones, J.W. The European Freshwater Eel; Liverpool University: Liverpool, UK, 1975. [Google Scholar]
- Albanese, B.; Angermeier, P.L.; Gowan, C. Designing Mark–Recapture Studies to Reduce Effects of Distance Weighting on Movement Distance Distributions of Stream Fishes. Trans. Am. Fish. Soc. 2003, 132, 925–939. [Google Scholar] [CrossRef]
- Crook, D.A. Is the home range concept compatible with the movements of two species of lowland river fish? J. Anim. Ecol. 2004, 73, 353–366. [Google Scholar] [CrossRef]
- Baker, M.; Nur, N.; Geupel, G.R. Correcting biased estimates of dispersal and survival due to limited study area: Theory and application using wren tits. Condor 1995, 7, 663–674. [Google Scholar] [CrossRef]
- Koenig, W.D.; Van Vuren, D.; Hooge, P.N. Detectability, philopatry, and the distribution of dispersal distances in vertebrates. Trends Ecol. Evol. 1996, 11, 514–517. [Google Scholar] [CrossRef]
- Gerking, S.D. The Restricted Movement of Fish Populations. Biol. Rev. 1959, 34, 221–242. [Google Scholar] [CrossRef]
- Gowan, C.; Young, M.K.; Fausch, K.D.; Riley, S.C. Restricted movement in resident stream salmonids: A paradigm lost? Can. J. Fish. Aquat. Sci. 1994, 51, 2626–2637. [Google Scholar] [CrossRef]
- Gowan, C.; Fausch, K.D. Mobile brook trout in two high-elevation Colorado streams: Re-evaluating the concept of restricted movement. Can. J. Fish. Aquat. Sci. 1996, 53, 1370–1381. [Google Scholar] [CrossRef]
- Fraser, D.F.; Gilliam, J.F.; Daley, M.J.; Le, A.N.; Skalski, G.T. Explaining Leptokurtic Movement Distributions: Intra population Variation in Boldness and Exploration. Am. Nat. 2001, 158, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.A. Restricted Movement in Stream Fish: The Paradigm Is Incomplete, Not Lost. Ecology 2002, 83, 1–13. [Google Scholar] [CrossRef]
- Durif, C.; Guibert, A.; Elie, P. Morphological discrimination of the silvering stages of the European eel. Am. Fish. Soc. 2009, 58, 103–111. [Google Scholar]
- Bolland, J.D.; Cowx, I.G.; Lucas, M.C. Evaluation of VIE and PIT tagging methods for juvenile cyprinid fishes. J. Appl. Ichthyol. 2009, 25, 381–386. [Google Scholar] [CrossRef]
- Hirt-Chabbert, J.A.; Young, O.A. Effects of surgically implanted PIT tags on growth, survival and tag retention of yellow shortfin eels Anguilla australis under laboratory conditions. J. Fish Biol. 2012, 81, 314–319. [Google Scholar] [CrossRef]
- Laffaille, P.; Acou, A.; Guillouët, J. The yellow European eel (Anguilla anguilla L.) may adopt a sedentary lifestyle in inland freshwaters. Ecol. Freshw. Fish 2005, 14, 191–196. [Google Scholar] [CrossRef]
- Chadwick, S.; Knights, B.; Thorley, J.L.; Bark, A. A long-term study of population characteristics and downstream migrations of the European eel Anguilla anguilla (L.) and the effects of a migration barrier in the Girnock Burn, north-east Scotland. J. Fish Biol. 2007, 70, 1535–1553. [Google Scholar] [CrossRef]
- Charrier, F.; Mazel, V.; Caraguel, J.M.; Abdallah, Y.; Le Gurun, J.; Legault, A.; Laffaille, P. Escapement of a silver-phase eel population, Anguilla anguilla, determined from fishery in a Mediterranean lagoon (Or, France). ICES J. Mar. Sci. 2012, 69, 30–33. [Google Scholar] [CrossRef]
- Mazel, V.; Charrier, F.; Legault, A.; Laffaille, P. Long-term effects of passive integrated transponder tagging (PIT tags) on the growth of the yellow European eel (Anguilla anguilla (Linnaeus, 1758)). J. Appl. Ichthyol. 2013, 29, 906–908. [Google Scholar] [CrossRef]
- Alldredge, P.; Gutierrez, M.; Duvernell, D.; Schaefer, J.; Brunkow, P.; Matamoros, W. Variability in movement dynamics of topminnow (Fundulus notatus and F. olivaceus) populations. Ecol. Freshw. Fish 2011, 20, 513–521. [Google Scholar] [CrossRef]
- Ford, T.E.; Mercer, E. Density, size distribution and home range of American eels, Anguilla rostrata, in a Massachusetts salt marsh. Environ. Biol. Fishes 1986, 17, 309–314. [Google Scholar] [CrossRef]
- Armstrong, J.D.; Braithwaite, V.A.; Huntingford, F.A. Spatial Strategies of Wild Atlantic Salmon Parr: Exploration and Settlement in Unfamiliar Areas. J. Anim. Ecol. 1997, 66, 203–211. [Google Scholar] [CrossRef]
- Gilliam, J.F.; Fraser, D.F. Corridor movement: Enhancement by predation threat, habitat structure and disturbance. Ecology 2001, 82, 258–273. [Google Scholar] [CrossRef]
- Mann, H. Über das rückkehrvermogen verpflantzer fluss-aal. Arch. Fischereiwiss. 1965, 15, 177–185. [Google Scholar]
- McGovern, P.; McCarthy, T.K. Local movements of freshwater eels (Anguilla anguilla L.) in western Ireland. In Wildlife Telemetry: Remote Sensing and Monitoring of Animals; Priede, I.G., Swift, S.M., Eds.; Ellis Horwood: Chichester, UK, 1992; pp. 319–327. [Google Scholar]
- LaBar, G.W.; Facey, D.E. Local Movements and Inshore Population Sizes of American Eels in Lake Champlain, Vermont. Trans. Am. Fish. Soc. 1983, 112, 111–116. [Google Scholar] [CrossRef]
- Bozeman, E.L.; Helfman, G.S.; Richardson, T. Population Size and Home Range of American Eels in a Georgia Tidal Creek. Trans. Am. Fish. Soc. 1985, 114, 821–825. [Google Scholar] [CrossRef]
- Dutil, J.-D.; Giroux, A.; Kemp, A.; Lavoie, G.; Dallaire, J.-P. Tidal Influence on Movements and on Daily Cycle of Activity of American Eels. Trans. Am. Fish. Soc. 1988, 117, 488–494. [Google Scholar] [CrossRef]
- Parker, S. Homing Ability and Home Range of Yellow-Phase American Eels in a Tidally Dominated Estuary. J. Mar. Biol. Assoc. UK 1995, 75, 127–140. [Google Scholar] [CrossRef]
- Rossi, R.; Bianchini, M.; Carrieri, A.; Franzoi, P. Observations on movements of yellow eels, Anguilla anguilla L., after displacement from coastal waters to sea. J. Fish Biol. 1987, 31, 155–164. [Google Scholar] [CrossRef]
- Baisez, A. Optimisation des suivis des indices d’abondances et des structures de taille de l’anguille europeénne (Anguilla anguilla, L.) dans un marais endigué de la côte atlantique: Relations espèce–habitat. Ph.D. Thesis, University of Toulouse, Toulouse, France, 2001. [Google Scholar]
- Lamothe, P.J.; Gallagher, M.; Chivers, D.P.; Moring, J.R. Homing and Movement of Yellow-phase American Eels in Freshwater Ponds. Env. Biol. Fishes 2000, 58, 393–399. [Google Scholar] [CrossRef]
- Tesch, F.-W. Homing of eels (Anguilla anguilla) in the southern North Sea. Mar. Biol. 1967, 1, 2–9. [Google Scholar] [CrossRef]
- Laffaille, P.; Feunteun, E.; Baisez, A.; Robinet, T.; Acou, A.; Legault, A.; Lek, S. Spatial organisation of European eel (Anguilla anguilla L.) in a small catchment. Ecol. Freshw. Fish 2003, 12, 254–264. [Google Scholar] [CrossRef]
- Edeline, E.; Dufour, S.; Elie, P. Proximate and Ultimate Control of Eel Continental Dispersal. In Spawning Migration of the European Eel; Van den Thillart, G., Dufour, S., Rankin, J.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 433–461. [Google Scholar]
- Braithwhaite, V.A.; De Perera, B. Short-range orientation in fish: How fish map space. Mar. Fresh. Behav. Physiol. 2006, 39, 37–47. [Google Scholar] [CrossRef]
- Knights, B. Eel biology and the status of recruitment and stocks. In Proceedings of the 2009 Eel Conference; Eel Management: The State of the Art; Bunt, D.A., Don, A.M., Eds.; Institute of Fisheries Management: Kingston upon Hull, UK, 2011; pp. 22–40. [Google Scholar]
- Hammond, S.D.; Welsh, S.A. Seasonal Movements of Large Yellow American Eels Downstream of a Hydroelectric Dam, Shenandoah River, West Virgina. In Eels at the Edge: Science, Status, and Conservation Concerns; Casselman, J.M., Cairns, D.K., Eds.; American Fisheries Society: Bethesda, MD, USA, 2009; Volume 58, pp. 309–323. [Google Scholar]
- Cullen, P.; McCarthy, T.K. Eels (Anguilla anguilla (L.)) of the lower River Shannon, with particular reference to seasonality in their activity and feeding ecology biology and environment. Proc. R. Ir. Acad. 2007, 107, 87–94. [Google Scholar] [CrossRef]
- Dekker, W. Did lack of spawners cause the collapse of the European eel, Anguilla anguilla? Fish. Manag. Ecol. 2003, 10, 365–376. [Google Scholar] [CrossRef]
- Grupo de Investigación Integrada en Áreas Litorales. Gestión Integrada de Zonas Costeras y Cuencas Hidrográficas: Introducción a un Caso de Estudio; Universidad de Cádiz: Cádiz, Spain, 2009. [Google Scholar]
- Durif, C.; Dufour, S.; Elie, P. The silvering process of Anguilla anguilla: A new classification from the yellow resident to the silver migrating stage. J. Fish Biol. 2005, 66, 1025–1043. [Google Scholar] [CrossRef]
- Sutton, S.G.; Bult, T.P.; Haedrich, R.L. Relationships among Fat Weight, Body Weight, Water Weight, and Condition Factors in Wild Atlantic Salmon Parr. Trans. Am. Fish. Soc. 2000, 129, 527–538. [Google Scholar] [CrossRef]
- Pollock, K.H. A Capture-Recapture Design Robust to Unequal Probability of Capture. J. Wildl. Manag. 1982, 46, 752–757. [Google Scholar] [CrossRef]
- Kendall, W.L.; Nichols, J.D.; Hines, J.E. Estimating Temporary Emigration Using Capture–Recapture Data with Pollock’s Robust Design. Ecology 1997, 78, 563–578. [Google Scholar]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models. Monographs on Statistics and Applied Probability 37; Chapman & Hall: London, UK, 1989. [Google Scholar]
- Lawal, H.B. Categorical Data Analysis with SAS and SPSS Applications; Erlbaum Associates Publishers: Mahwah, NJ, USA, 2003. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multi-Model Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Buckland, S.T.; Burnham, K.P.; Augustin, N. Model Selection: An Integral Part of Inference. Biometrics 1997, 53, 603–618. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Length (cm) | N | Mean ± 95% C.I. (m) | 5% Trimmed Mean (m) | M–Estimators (m) | Median (m) | Range (m) | CV | Outliers (m) | Extreme Values (m) | Mean Area ± 95% C.I. (m2) |
|---|---|---|---|---|---|---|---|---|---|---|
| 25.0–29.9 | 137 | 65.47 ± 17.98 | 47.55 | 24.38–32.02 | 26.49 | 707.04 | 1.63 | 165.08–268.11 | 319.51–707.04 | 1023.97 ± 281.21 |
| 30.0–34.9 | 122 | 78.67 ± 19.65 | 62.4 | 36.96–44.50 | 39.79 | 622.09 | 1.39 | 197.21–271.42 | 300.05–622.55 | 1230.42 ± 307.33 |
| 35.0–39.9 | 18 | 99.79 ± 51.92 | 93.13 | 40.06–58.93 | 48.72 | 316.73 | 1.05 | No | No | 1560.74 ± 812.04 |
| 40.0–49.9 | 13 | 158.61 ± 79.56 | 151.14 | 75.94–99.27 | 84.13 | 363.81 | 0.83 | No | No | 2480.70 ± 1244.34 |
| >50.No0 | 6 | 258.76 ± 99.07 | 257.44 | 249.30–252.58 | 242.21 | 264.44 | 0.37 | No | No | 4047.08 ± 1549.48 |
| Model | Omnibus Test | Deviance | AICc | ΔAICc | wi |
|---|---|---|---|---|---|
| y = intercept-TL-RF | <0.05 | 436.907 | 768.576 | 0.000 | 0.6053 |
| y = intercept-TL-WT | <0.05 | 489.689 | 769.568 | 0.992 | 0.3686 |
| y = intercept-TL-ΔRF | <0.05 | 599.930 | 776.567 | 6.999 | 0.0111 |
| y = intercept-TL | <0.05 | 578.992 | 777.577 | 9.001 | 0.0067 |
| y = intercept-TL-ΔRF-interaction | <0.05 | 612.624 | 778.378 | 9.802 | 0.0045 |
| y = intercept-TL-RF-interaction | <0.05 | 607.686 | 779.684 | 11.108 | 0.0023 |
| y = intercept-TL-WT-interaction | <0.05 | 605.243 | 781.4 | 12.824 | 0.0010 |
| y = intercept-RF | <0.05 | 625.660 | 784.828 | 16.252 | 0.0002 |
| y = intercept-WT | <0.05 | 629.322 | 784.907 | 16.331 | 0.0002 |
| y = intercept | 638.827 | 791.328 | 22.752 | 0.0000 |
| Parameter Estimate | 95% CI | w+ | |
|---|---|---|---|
| α Intercepts (Movements extension category, m) | - | ||
| <49.9 | 0.58 | ±0.12 | - |
| 50–99.9 | 1.62 | ±0.51 | - |
| 100–199.9 | 2.54 | ±0.21 | - |
| 200–299.9 | 3.20 | ±0.28 | - |
| β Coefficients | - | ||
| Total Length | 0.44 | ±0.10 | 0.99 |
| River Flow | 0.39 | ±0.16 | 0.61 |
| Water Temperature | −0.27 | ±0.16 | 0.37 |
| Reference | Spp | Environment | Home Range Size | Relation with Length | Territoriality | Methodology |
|---|---|---|---|---|---|---|
| Present study | Aa | River | 0.102–0.405 ha (65.47–258.76 lineal m) | yes | yes | M-R |
| Mann [43] | Aa | River | 30 km | * | * | M-R |
| LaBar et al. [21] | Aa | Lagoon | 0.135–0.25 ha | * | * | T |
| Baras et al. [17] | Aa | River | 0.01–0.01 ha (maximum home range 281 lineal m) | * | yes | T |
| McGovern y McCarthy [44] | Aa | River | 0.1–0.6 ha | * | yes | T |
| Ovidio et al. [11] | Aa | River | 33–341 m (average 62 m) | no | yes | T |
| Walker et al. [16] | Aa | Coastal lagoon | 630–4150 m | no | yes | T |
| Barry et al. [13] | Aa | Lagoon | 14.3–29.6 ha | yes | yes | T |
| Verhelst et al. [19] | Aa | Polder area | 300–3917.34 (lineal m) | * | yes | T |
| LaBar & Facey [45] | Ar | Lagoon | 2–65 ha | * | * | T |
| Bozeman et al. [46] | Ar | Coastal area | 1 ha | * | * | M-R |
| Ford & Mercer [40] | Ar | Marshland | 0.021 ha | no | yes | M-R |
| Dutil et al. [47] | Ar | River | 0.5-2.0 ha | * | * | T |
| Parker [48] | Ar | Estuary | 325 ± 64 ha | * | * | T |
| Thibault et al. [20] | Ar | Estuary-river | 16 ha | yes | * | T |
| Béger-Pon et al. [12] | Ar | Estuary-river-Lagoon | 11.1–200 km | no | yes | T |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrera, M.; Moreno-Valcárcel, R.; De Miguel Rubio, R.; Fernández-Delgado, C. From Transient to Sedentary? Changes in the Home Range Size and Environmental Patterns of Movements of European Eels (Anguilla anguilla) in a Mediterranean River. Fishes 2019, 4, 43. https://doi.org/10.3390/fishes4030043
Herrera M, Moreno-Valcárcel R, De Miguel Rubio R, Fernández-Delgado C. From Transient to Sedentary? Changes in the Home Range Size and Environmental Patterns of Movements of European Eels (Anguilla anguilla) in a Mediterranean River. Fishes. 2019; 4(3):43. https://doi.org/10.3390/fishes4030043
Chicago/Turabian StyleHerrera, Mercedes, Raquel Moreno-Valcárcel, Ramón De Miguel Rubio, and Carlos Fernández-Delgado. 2019. "From Transient to Sedentary? Changes in the Home Range Size and Environmental Patterns of Movements of European Eels (Anguilla anguilla) in a Mediterranean River" Fishes 4, no. 3: 43. https://doi.org/10.3390/fishes4030043
APA StyleHerrera, M., Moreno-Valcárcel, R., De Miguel Rubio, R., & Fernández-Delgado, C. (2019). From Transient to Sedentary? Changes in the Home Range Size and Environmental Patterns of Movements of European Eels (Anguilla anguilla) in a Mediterranean River. Fishes, 4(3), 43. https://doi.org/10.3390/fishes4030043