Development and Characterization of a Luminescence-Based High-Throughput Serum Bactericidal Assay (L-SBA) to Assess Bactericidal Activity of Human Sera against Nontyphoidal Salmonella

 ,
, 
 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Reagents
2.2. Serum Samples
2.3. Luminescence-Based SBA (L-SBA)
2.4. Calculations
2.5. Statistical Analysis
2.6. Ethical Statement
3. Results
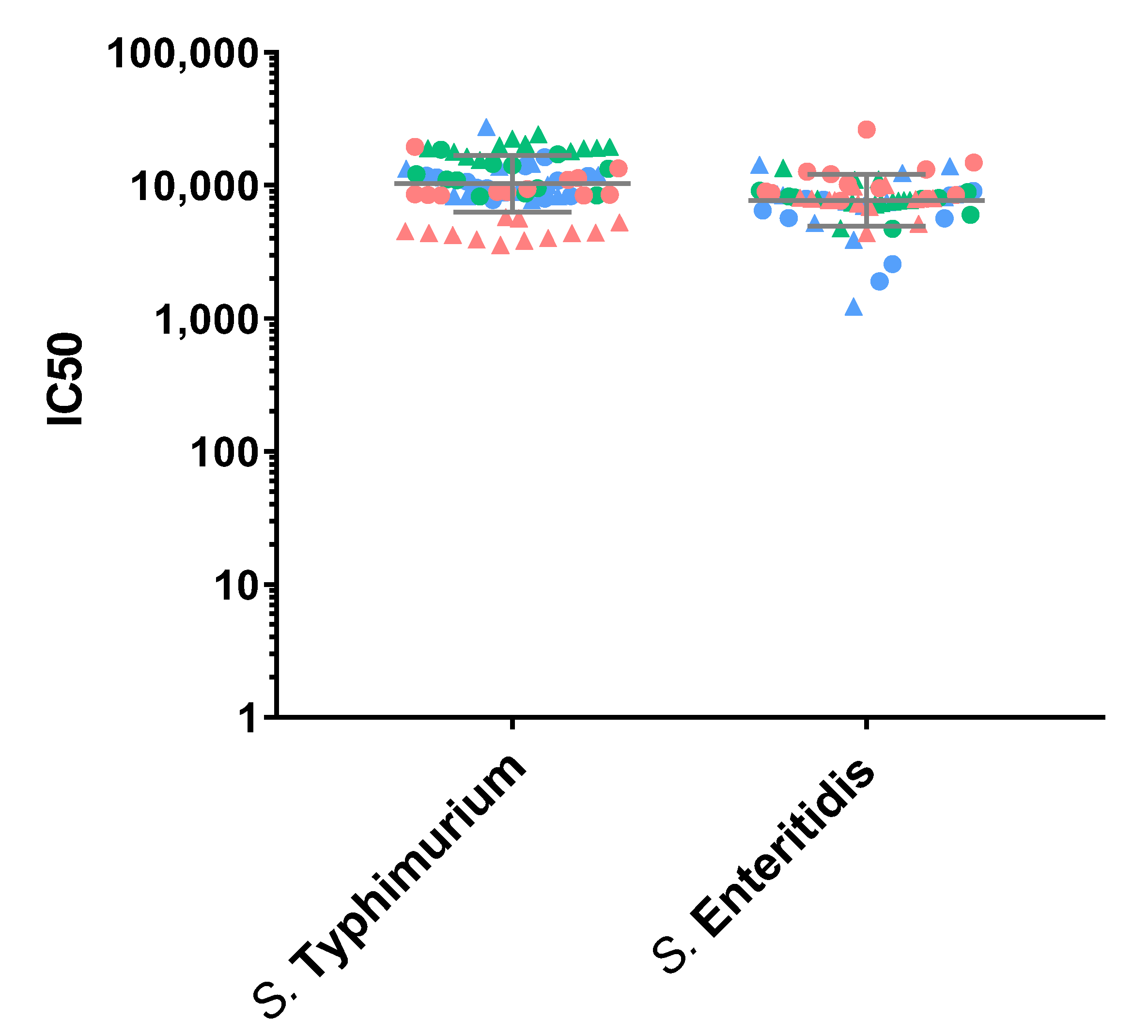
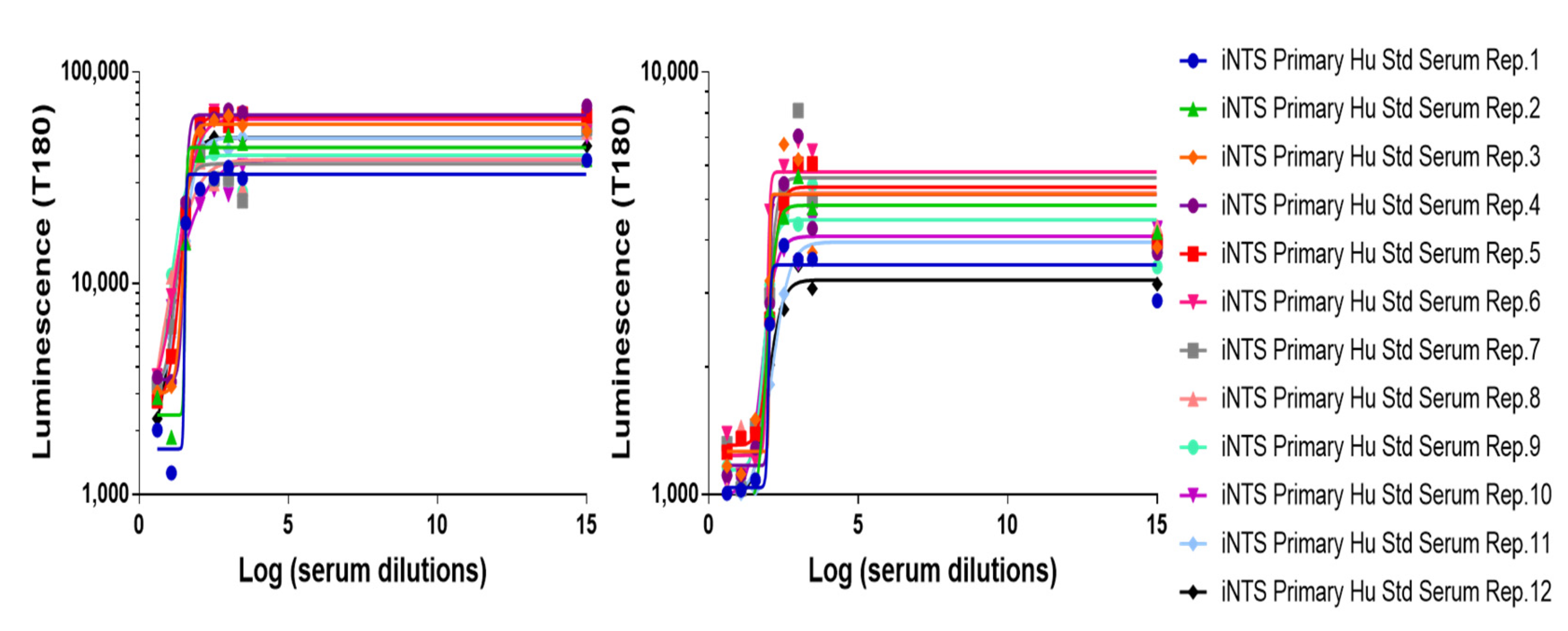
3.1. Precision
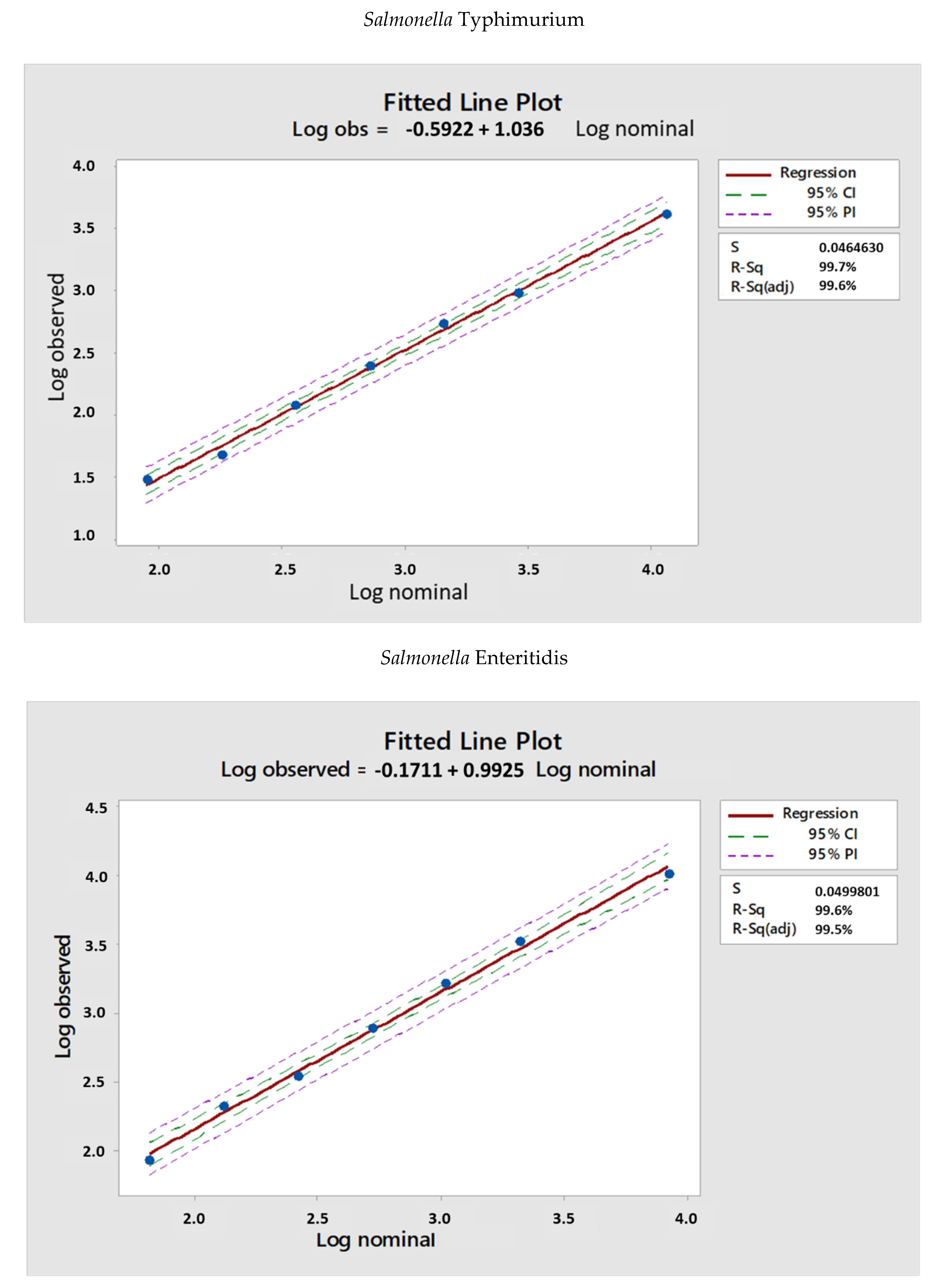
3.2. Linearity
3.3. Limit of Detection (LoD) and Limit of Quantification (LoQ)
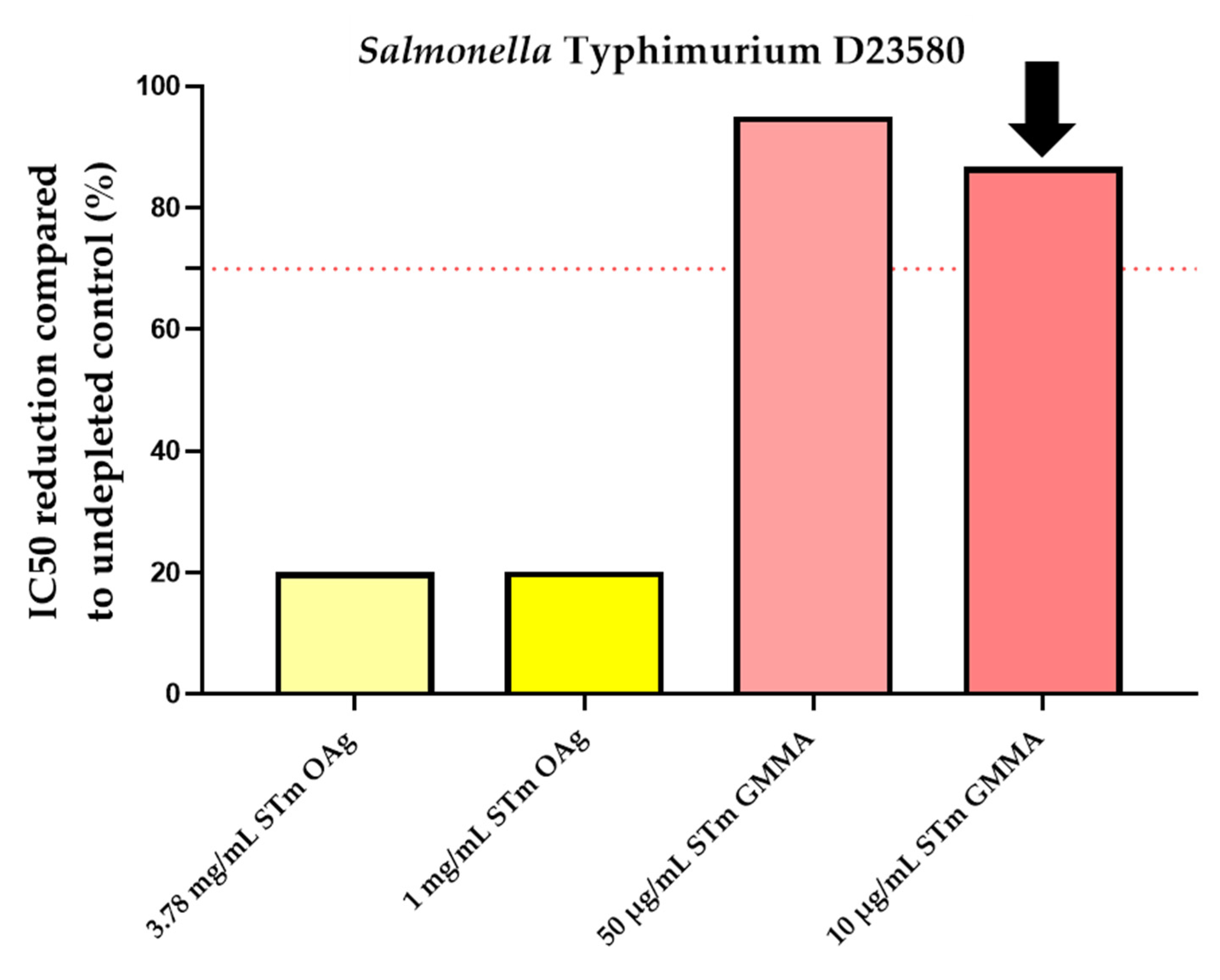
3.4. Specificity
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sponsorship
References
- Bula-Rudas, F.J.; Rathore, M.H.; Maraqa, N.F. Salmonella Infections in Childhood. Adv. Pediatr. 2015, 62, 29–58. [Google Scholar] [CrossRef] [PubMed]
- Feasey, N.A.; Dougan, G.; Kingsley, R.A.; Heyderman, R.S.; Gordon, M.A. Invasive non-typhoidal salmonella disease: An emerging and neglected tropical disease in Africa. Lancet 2012, 379, 2489–2499. [Google Scholar] [CrossRef] [PubMed]
- GBD 2017 Non-Typhoidal Salmonella Invasive Disease Collaborators. The global burden of non-typhoidal salmonella invasive disease: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Infect. Dis. 2019, 19, 1312–1324. [Google Scholar] [CrossRef]
- Marchello, C.S.; Fiorino, F.; Pettini, E.; Crump, J.A.; Vacc-iNTS Consortium Collaborators. Incidence of non-typhoidal Salmonella invasive disease: A systematic review and meta-analysis. J. Infect. 2021, 83, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Bronzan, R.N.; Taylor, T.E.; Mwenechanya, J.; Tembo, M.; Kayira, K.; Bwanaisa, L.; Njobvu, A.; Kondowe, W.; Chalira, C.; Walsh, A.L.; et al. Bacteremia in Malawian children with severe malaria: Prevalence, etiology, HIV coinfection, and outcome. J. Infect. Dis. 2007, 195, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Calis, J.C.; Phiri, K.S.; Faragher, E.B.; Brabin, B.J.; Bates, I.; Cuevas, L.E.; de Haan, R.J.; Phiri, A.I.; Malange, P.; Khoka, M.; et al. Severe anemia in Malawian children. N. Engl. J. Med. 2008, 358, 888–899. [Google Scholar] [CrossRef] [PubMed]
- Feasey, N.A.; Everett, D.; Faragher, E.B.; Roca-Feltrer, A.; Kang’ombe, A.; Denis, B.; Kerac, M.; Molyneux, E.; Molyneux, M.; Jahn, A.; et al. Modelling the Contributions of Malaria, HIV, Malnutrition and Rainfall to the Decline in Paediatric Invasive Non-typhoidal Salmonella Disease in Malawi. PLoS Negl. Trop. Dis. 2015, 9, e0003979. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.A. Invasive nontyphoidal Salmonella disease: Epidemiology, pathogenesis and diagnosis. Curr. Opin. Infect. Dis. 2011, 24, 484–489. [Google Scholar] [CrossRef]
- Greenwood, B.M.; Bradley-Moore, A.M.; Bryceson, A.D.; Palit, A. Immunosuppression in children with malaria. Lancet 1972, 1, 169–172. [Google Scholar] [CrossRef]
- MacLennan, C.A.; Gilchrist, J.J.; Gordon, M.A.; Cunningham, A.F.; Cobbold, M.; Goodall, M.; Kingsley, R.A.; van Oosterhout, J.J.; Msefula, C.L.; Mandala, W.L.; et al. Dysregulated humoral immunity to nontyphoidal Salmonella in HIV-infected African adults. Science 2010, 328, 508–512. [Google Scholar] [CrossRef]
- MacLennan, C.A.; Gondwe, E.N.; Msefula, C.L.; Kingsley, R.A.; Thomson, N.R.; White, S.A.; Goodall, M.; Pickard, D.J.; Graham, S.M.; Dougan, G.; et al. The neglected role of antibody in protection against bacteremia caused by nontyphoidal strains of Salmonella in African children. J. Clin. Investig. 2008, 118, 1553–1562. [Google Scholar] [CrossRef] [PubMed]
- Marchello, C.S.; Birkhold, M.; Crump, J.A.; Vacc-iNTS Consortium Collaborators. Complications and mortality of non-typhoidal salmonella invasive disease: A global systematic review and meta-analysis. Lancet Infect. Dis. 2022, 22, 692–705. [Google Scholar] [CrossRef] [PubMed]
- Post, A.S.; Diallo, S.N.; Guiraud, I.; Lompo, P.; Tahita, M.C.; Maltha, J.; Van Puyvelde, S.; Mattheus, W.; Ley, B.; Thriemer, K.; et al. Supporting evidence for a human reservoir of invasive non-Typhoidal Salmonella from household samples in Burkina Faso. PLoS Negl. Trop. Dis. 2019, 13, e0007782. [Google Scholar] [CrossRef] [PubMed]
- Kariuki, S.; Revathi, G.; Kariuki, N.; Kiiru, J.; Mwituria, J.; Muyodi, J.; Githinji, J.W.; Kagendo, D.; Munyalo, A.; Hart, C.A. Invasive multidrug-resistant non-typhoidal Salmonella infections in Africa: Zoonotic or anthroponotic transmission? J. Med. Microbiol. 2006, 55, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Koolman, L.; Prakash, R.; Diness, Y.; Msefula, C.; Nyirenda, T.S.; Olgemoeller, F.; Wigley, P.; Perez-Sepulveda, B.; Hinton, J.C.D.; Owen, S.V.; et al. Case-control investigation of invasive Salmonella disease in Africa reveals no evidence of environmental or animal reservoirs of invasive strains. medRxiv 2022. [Google Scholar] [CrossRef]
- Tack, B.; Vanaenrode, J.; Verbakel, J.Y.; Toelen, J.; Jacobs, J. Invasive non-typhoidal Salmonella infections in sub-Saharan Africa: A systematic review on antimicrobial resistance and treatment. BMC Med. 2020, 18, 212. [Google Scholar] [CrossRef] [PubMed]
- Van Puyvelde, S.; Pickard, D.; Vandelannoote, K.; Heinz, E.; Barbe, B.; de Block, T.; Clare, S.; Coomber, E.L.; Harcourt, K.; Sridhar, S.; et al. An African Salmonella Typhimurium ST313 sublineage with extensive drug-resistance and signatures of host adaptation. Nat. Commun. 2019, 10, 4280. [Google Scholar] [CrossRef]
- Park, S.E.; Pham, D.T.; Pak, G.D.; Panzner, U.; Maria Cruz Espinoza, L.; von Kalckreuth, V.; Im, J.; Mogeni, O.D.; Schutt-Gerowitt, H.; Crump, J.A.; et al. The genomic epidemiology of multi-drug resistant invasive non-typhoidal Salmonella in selected sub-Saharan African countries. BMJ Glob. Health 2021, 6, e005659. [Google Scholar] [CrossRef]
- MacLennan, C.A.; Martin, L.B.; Micoli, F. Vaccines against invasive Salmonella disease: Current status and future directions. Hum. Vaccines Immunother. 2014, 10, 1478–1493. [Google Scholar] [CrossRef]
- Tennant, S.M.; MacLennan, C.A.; Simon, R.; Martin, L.B.; Khan, M.I. Nontyphoidal salmonella disease: Current status of vaccine research and development. Vaccine 2016, 34, 2907–2910. [Google Scholar] [CrossRef]
- De Benedetto, G.; Alfini, R.; Cescutti, P.; Caboni, M.; Lanzilao, L.; Necchi, F.; Saul, A.; MacLennan, C.A.; Rondini, S.; Micoli, F. Characterization of O-antigen delivered by Generalized Modules for Membrane Antigens (GMMA) vaccine candidates against nontyphoidal Salmonella. Vaccine 2017, 35, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Rossi, O.; Caboni, M.; Negrea, A.; Necchi, F.; Alfini, R.; Micoli, F.; Saul, A.; MacLennan, C.A.; Rondini, S.; Gerke, C. Toll-like Receptor Activation by Generalized Modules for Membrane Antigens from Lipid A Mutants of Salmonella enterica Serovars Typhimurium and Enteritidis. Clin. Vaccine Immunol. 2016, 23, 304–314. [Google Scholar] [CrossRef]
- Micoli, F.; Rondini, S.; Alfini, R.; Lanzilao, L.; Necchi, F.; Negrea, A.; Rossi, O.; Brandt, C.; Clare, S.; Mastroeni, P.; et al. Comparative immunogenicity and efficacy of equivalent outer membrane vesicle and glycoconjugate vaccines against nontyphoidal Salmonella. Proc. Natl. Acad. Sci. USA 2018, 115, 10428–10433. [Google Scholar] [CrossRef] [PubMed]
- Berlanda Scorza, F.; Colucci, A.M.; Maggiore, L.; Sanzone, S.; Rossi, O.; Ferlenghi, I.; Pesce, I.; Caboni, M.; Norais, N.; Di Cioccio, V.; et al. High yield production process for Shigella outer membrane particles. PLoS ONE 2012, 7, e35616. [Google Scholar] [CrossRef]
- Gerke, C.; Colucci, A.M.; Giannelli, C.; Sanzone, S.; Vitali, C.G.; Sollai, L.; Rossi, O.; Martin, L.B.; Auerbach, J.; Di Cioccio, V.; et al. Production of a Shigella sonnei Vaccine Based on Generalized Modules for Membrane Antigens (GMMA), 1790GAHB. PLoS ONE 2015, 10, e0134478. [Google Scholar] [CrossRef] [PubMed]
- Kis, Z.; Shattock, R.; Shah, N.; Kontoravdi, C. Emerging Technologies for Low-Cost, Rapid Vaccine Manufacture. Biotechnol. J. 2019, 14, 1800376. [Google Scholar] [CrossRef] [PubMed]
- Mastroeni, P.; Rossi, O. Antibodies and Protection in Systemic Salmonella Infections: Do We Still Have More Questions than Answers? Infect. Immun. 2020, 88(10), e00219-20. [Google Scholar] [CrossRef] [PubMed]
- Borrow, R.; Carlone, G.M.; Rosenstein, N.; Blake, M.; Feavers, I.; Martin, D.; Zollinger, W.; Robbins, J.; Aaberge, I.; Granoff, D.M.; et al. Neisseria meningitidis group B correlates of protection and assay standardization—International meeting report Emory University, Atlanta, Georgia, United States, 16–17 March 2005. Vaccine 2006, 24, 5093–5107. [Google Scholar] [CrossRef]
- Necchi, F.; Saul, A.; Rondini, S. Setup of luminescence-based serum bactericidal assay against Salmonella Paratyphi A. J. Immunol. Methods 2018, 461, 117–121. [Google Scholar] [CrossRef]
- Necchi, F.; Saul, A.; Rondini, S. Development of a high-throughput method to evaluate serum bactericidal activity using bacterial ATP measurement as survival readout. PLoS ONE 2017, 12, e0172163. [Google Scholar] [CrossRef]
- Aruta, M.G.; Carducci, M.; Micoli, F.; Necchi, F.; Rossi, O. Increasing the High Throughput of a Luminescence-Based Serum Bactericidal Assay (L-SBA). BioTech 2021, 10, 19. [Google Scholar] [CrossRef]
- Rossi, O.; Molesti, E.; Saul, A.; Giannelli, C.; Micoli, F.; Necchi, F. Intra-Laboratory Evaluation of Luminescence Based High-Throughput Serum Bactericidal Assay (L-SBA) to Determine Bactericidal Activity of Human Sera against Shigella. High-Throughput 2020, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, R.A.; Msefula, C.L.; Thomson, N.R.; Kariuki, S.; Holt, K.E.; Gordon, M.A.; Harris, D.; Clarke, L.; Whitehead, S.; Sangal, V.; et al. Epidemic multiple drug resistant Salmonella Typhimurium causing invasive disease in sub-Saharan Africa have a distinct genotype. Genome Res. 2009, 19, 2279–2287. [Google Scholar] [CrossRef] [PubMed]
- Onsare, R.S.; Micoli, F.; Lanzilao, L.; Alfini, R.; Okoro, C.K.; Muigai, A.W.; Revathi, G.; Saul, A.; Kariuki, S.; MacLennan, C.A.; et al. Relationship between antibody susceptibility and lipopolysaccharide O-antigen characteristics of invasive and gastrointestinal nontyphoidal Salmonellae isolates from Kenya. PLoS Negl. Trop. Dis. 2015, 9, e0003573. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Rondini, S.; Gavini, M.; Pisoni, I.; Lanzilao, L.; Colucci, A.M.; Giannelli, C.; Pippi, F.; Sollai, L.; Pinto, V.; et al. A scalable method for O-antigen purification applied to various Salmonella serovars. Anal. Biochem. 2013, 434, 136–145. [Google Scholar] [CrossRef]
- Darton, T.C.; Meiring, J.E.; Tonks, S.; Khan, M.A.; Khanam, F.; Shakya, M.; Thindwa, D.; Baker, S.; Basnyat, B.; Clemens, J.D.; et al. The STRATAA study protocol: A programme to assess the burden of enteric fever in Bangladesh, Malawi and Nepal using prospective population census, passive surveillance, serological studies and healthcare utilisation surveys. BMJ Open 2017, 7, e016283. [Google Scholar] [CrossRef]
- Giersing, B.K.; Porter, C.K.; Kotloff, K.; Neels, P.; Cravioto, A.; MacLennan, C.A. How can controlled human infection models accelerate clinical development and policy pathways for vaccines against Shigella? Vaccine 2019, 37, 4778–4783. [Google Scholar] [CrossRef]
- Nahm, M.H.; Yu, J.; Weerts, H.P.; Wenzel, H.; Tamilselvi, C.S.; Chandrasekaran, L.; Pasetti, M.F.; Mani, S.; Kaminski, R.W. Development, Interlaboratory Evaluations, and Application of a Simple, High-Throughput Shigella Serum Bactericidal Assay. mSphere 2018, 3, e00146-18. [Google Scholar] [CrossRef]
- Mak, P.A.; Santos, G.F.; Masterman, K.A.; Janes, J.; Wacknov, B.; Vienken, K.; Giuliani, M.; Herman, A.E.; Cooke, M.; Mbow, M.L.; et al. Development of an automated, high-throughput bactericidal assay that measures cellular respiration as a survival readout for Neisseria meningitidis. Clin. Vaccine Immunol. 2011, 18, 1252–1260. [Google Scholar] [CrossRef][Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. Typhimurium | S. Enteritidis | |
|---|---|---|
| LoD (IC50) | 9.3 | 8.7 |
| LoQ (IC50) | 51.3 | 42.3 |
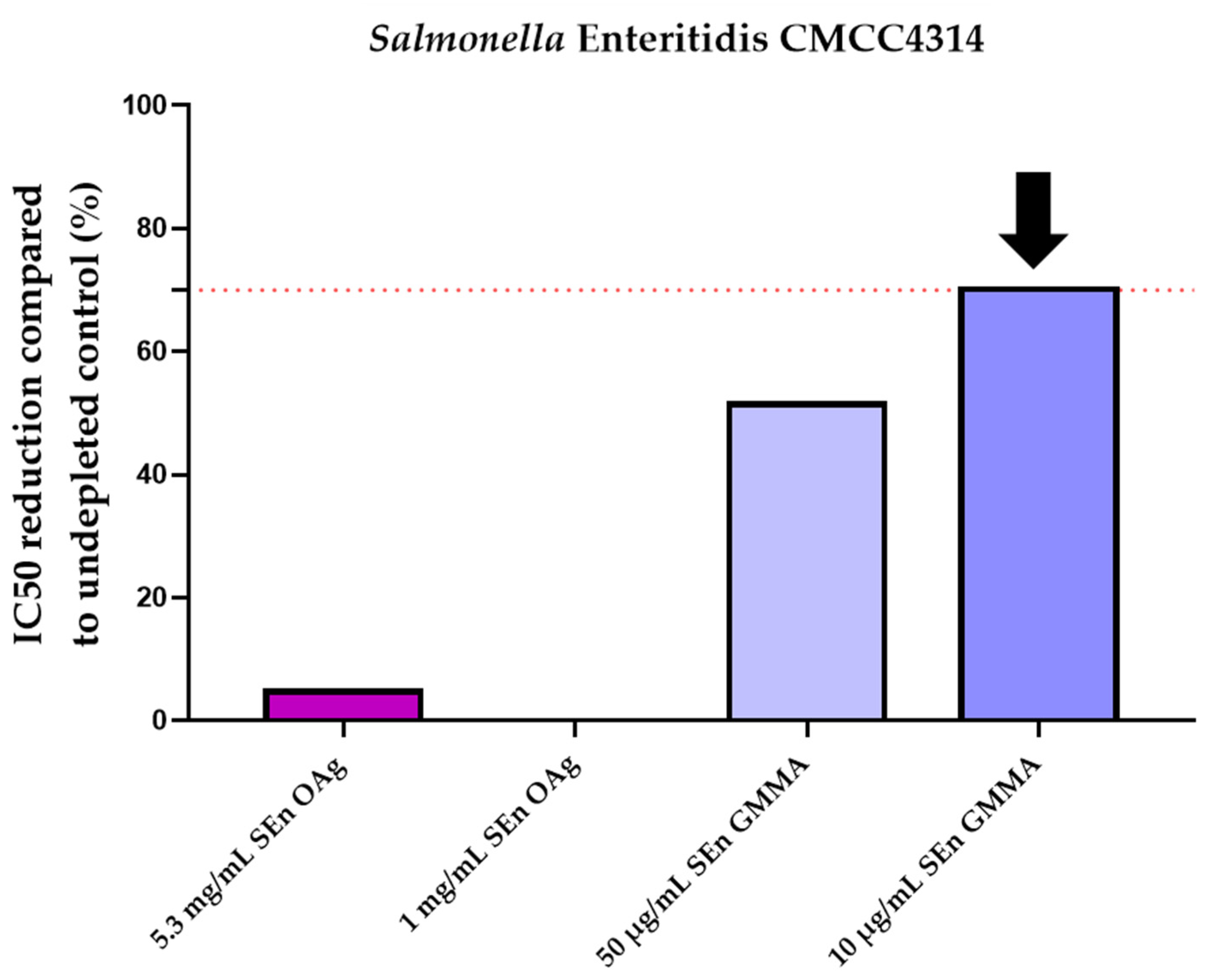
| SetUp of Minimum Homologous OAg Concentration to Inhibit IC50 ≥ 70% | |||
|---|---|---|---|
| S. Typhimurium | S. Enteritidis | ||
| Homologous GMMA concentration (µg/mL) | Percentage of inhibition compared to undepleted control | Homologous GMMA concentration (µg/mL) | Percentage of inhibition compared to undepleted control |
| 10 | 86.2 | 10 | 70.1 |
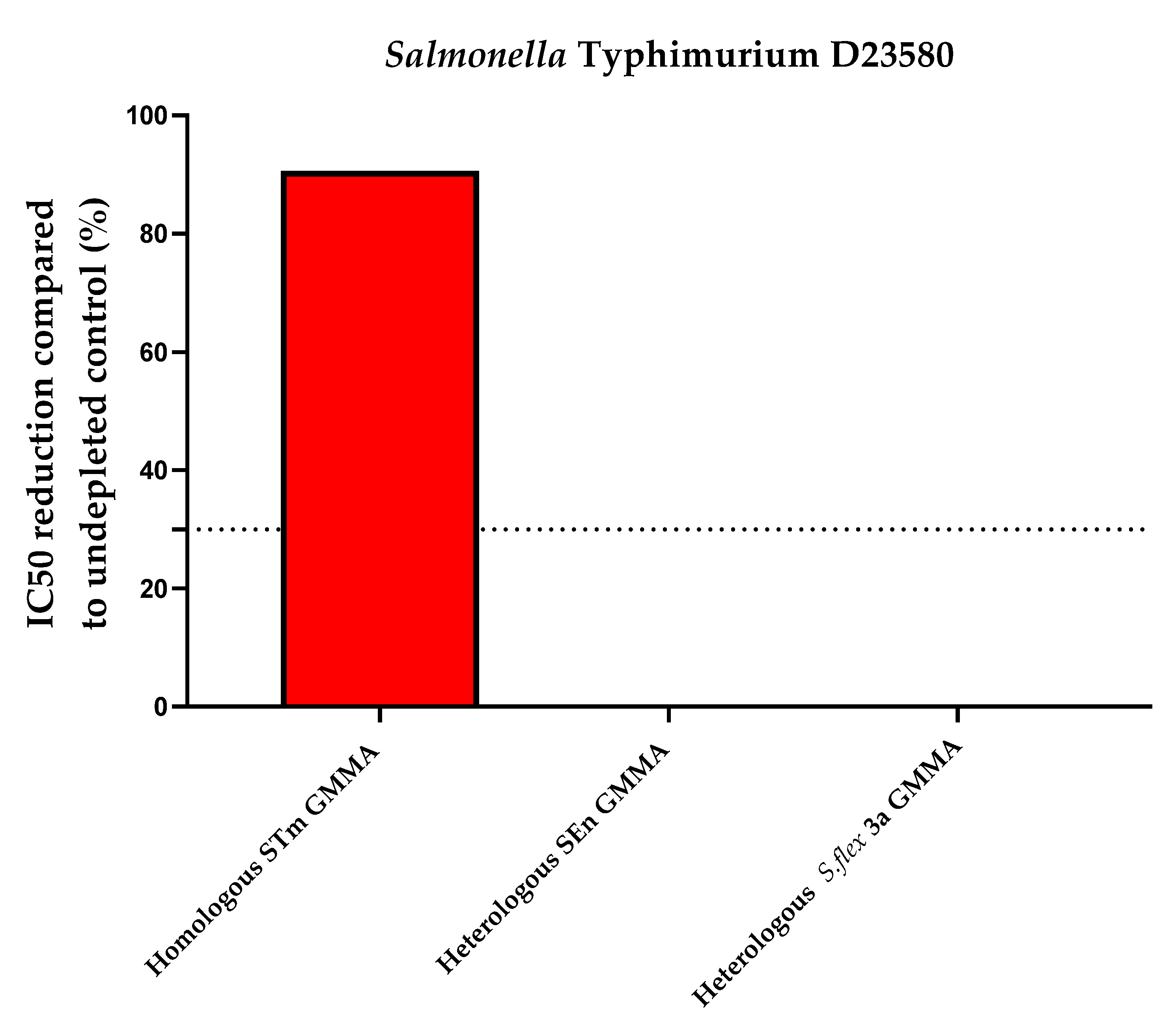
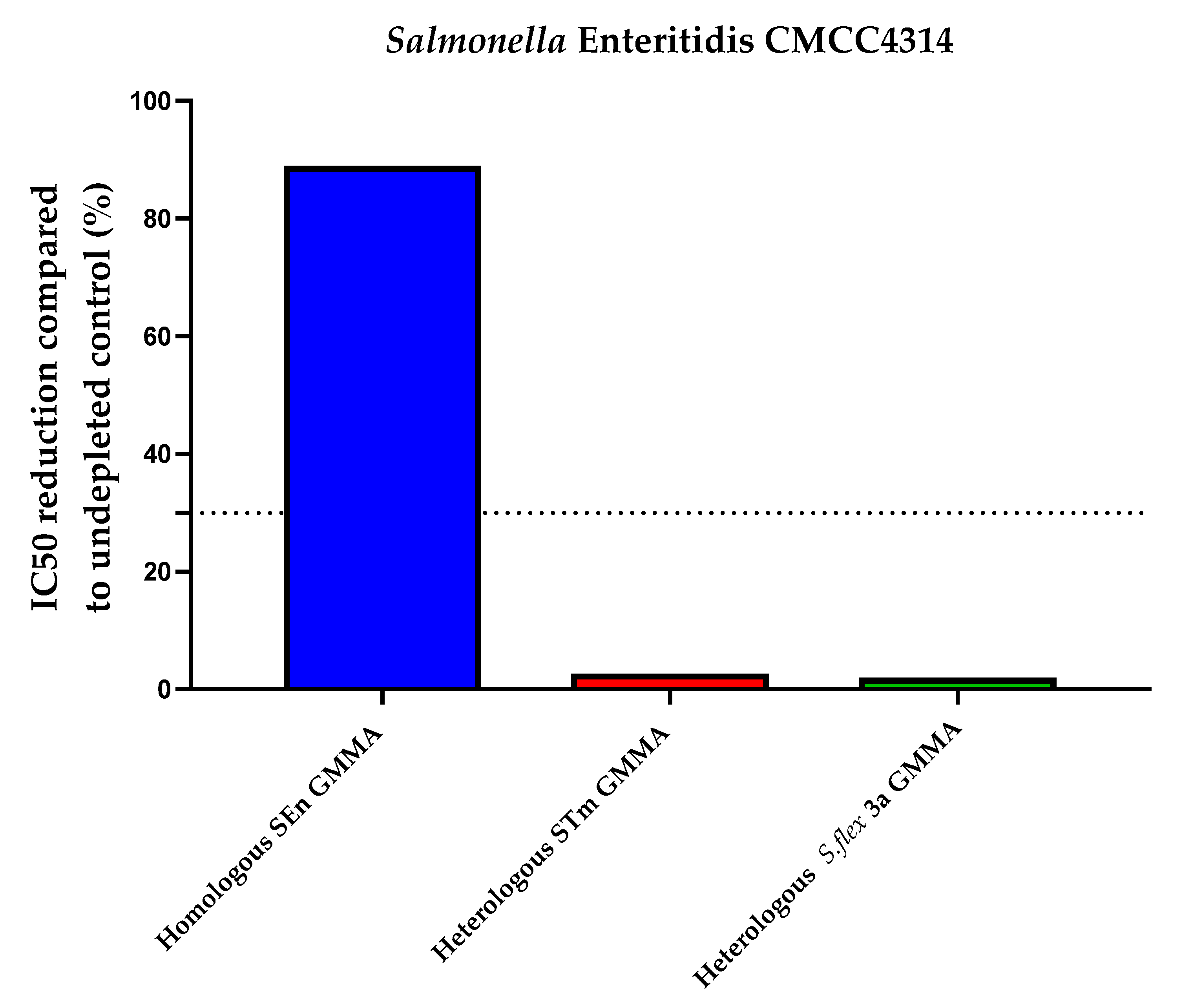
| Heterologous Specificity (Percentage of Inhibition Compared to Undepleted Control) | |||||
|---|---|---|---|---|---|
| S. Typhimurium | S. Enteritidis | ||||
| iNTS Primary Human Standard Serum depleted with 10 µg/mL STm GMMA | iNTS Primary Human Standard Serum depleted with 10 µg/mL SEn GMMA | iNTS Primary Human Standard Serum depleted with 10 µg/mL S.flex 3a GMMA | iNTS Primary Human Standard Serum depleted with 10 µg/mL SEn GMMA | iNTS Primary Human Standard Serum depleted with 10 µg/mL STm GMMA | iNTS Primary Human Standard Serum depleted with 10 µg/mL S.flex 3a GMMA |
| 90.1 | 0 | 0 | 88.4 | 2.2 | 1.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aruta, M.G.; De Simone, D.; Dale, H.; Chirwa, E.; Kadwala, I.; Mbewe, M.; Banda, H.; Gordon, M.; Pizza, M.; Berlanda Scorza, F.; et al. Development and Characterization of a Luminescence-Based High-Throughput Serum Bactericidal Assay (L-SBA) to Assess Bactericidal Activity of Human Sera against Nontyphoidal Salmonella. Methods Protoc. 2022, 5, 100. https://doi.org/10.3390/mps5060100
Aruta MG, De Simone D, Dale H, Chirwa E, Kadwala I, Mbewe M, Banda H, Gordon M, Pizza M, Berlanda Scorza F, et al. Development and Characterization of a Luminescence-Based High-Throughput Serum Bactericidal Assay (L-SBA) to Assess Bactericidal Activity of Human Sera against Nontyphoidal Salmonella. Methods and Protocols. 2022; 5(6):100. https://doi.org/10.3390/mps5060100
Chicago/Turabian StyleAruta, Maria Grazia, Daniele De Simone, Helen Dale, Esmelda Chirwa, Innocent Kadwala, Maurice Mbewe, Happy Banda, Melita Gordon, Mariagrazia Pizza, Francesco Berlanda Scorza, and et al. 2022. "Development and Characterization of a Luminescence-Based High-Throughput Serum Bactericidal Assay (L-SBA) to Assess Bactericidal Activity of Human Sera against Nontyphoidal Salmonella" Methods and Protocols 5, no. 6: 100. https://doi.org/10.3390/mps5060100
APA StyleAruta, M. G., De Simone, D., Dale, H., Chirwa, E., Kadwala, I., Mbewe, M., Banda, H., Gordon, M., Pizza, M., Berlanda Scorza, F., Nyirenda, T., Canals, R., Rossi, O., & on behalf of the Vacc-iNTS Consortium Collaborators. (2022). Development and Characterization of a Luminescence-Based High-Throughput Serum Bactericidal Assay (L-SBA) to Assess Bactericidal Activity of Human Sera against Nontyphoidal Salmonella. Methods and Protocols, 5(6), 100. https://doi.org/10.3390/mps5060100