
Graph Neural Network Learning on the Pediatric Structural Connectome

 , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Datasets
2.2. Structural Connectome Processing
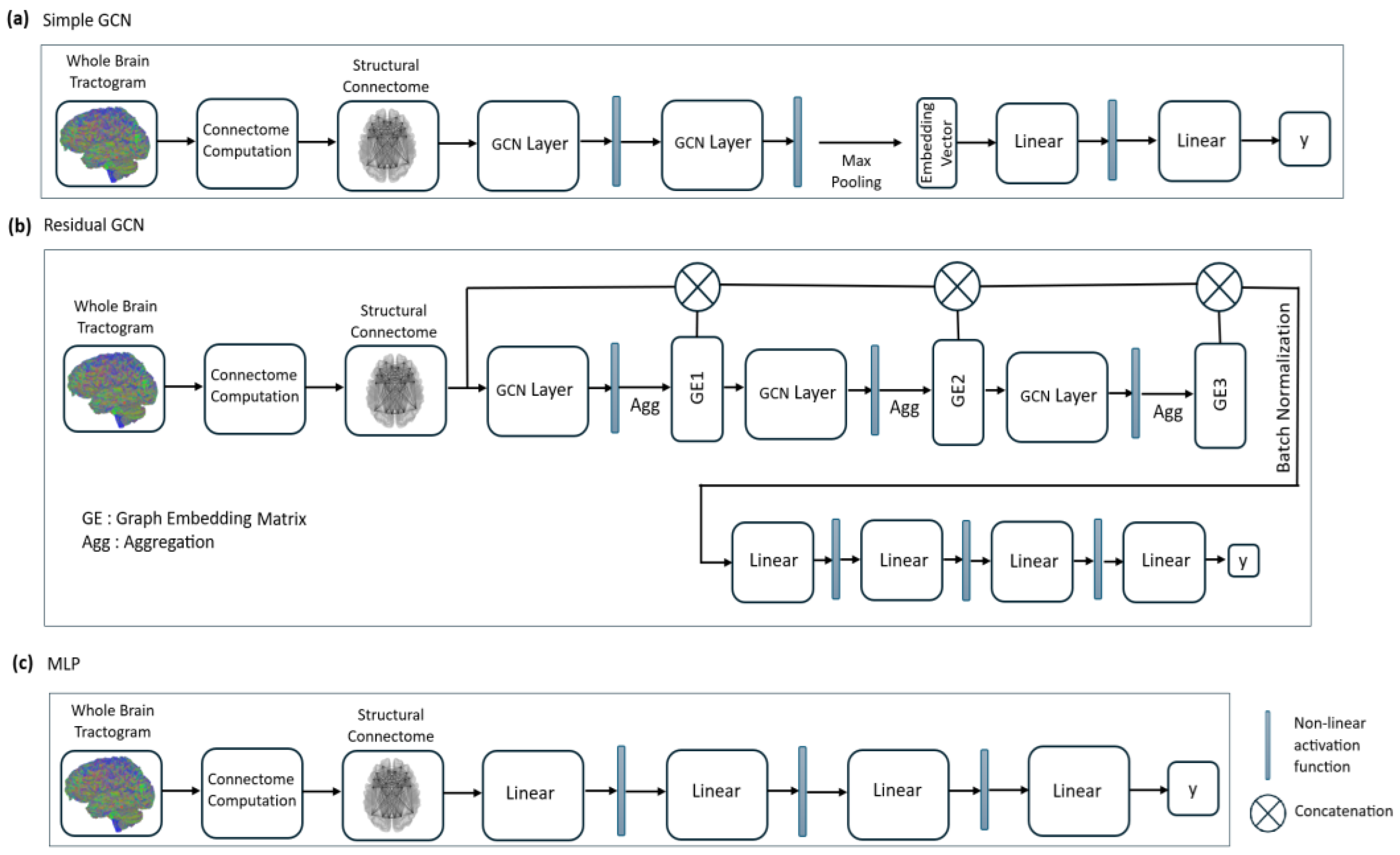
2.3. Graph Convolutional Network (GCN) Model Definition
2.4. Model Architecture
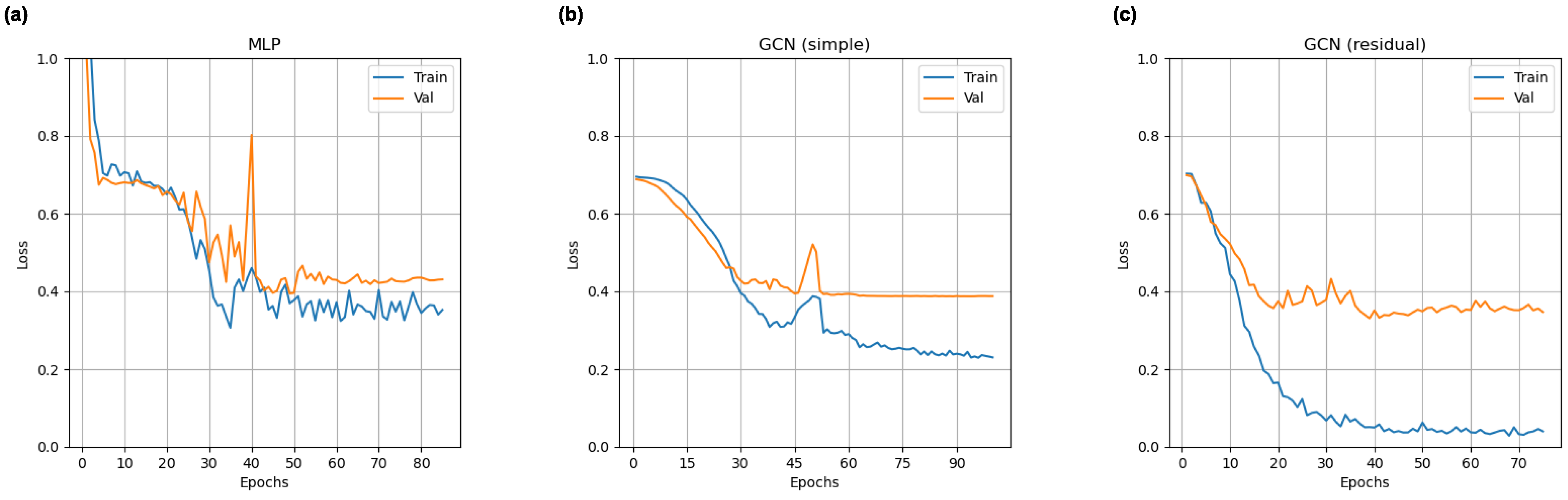
2.5. Training Procedure
2.6. Model Evaluation Experiments
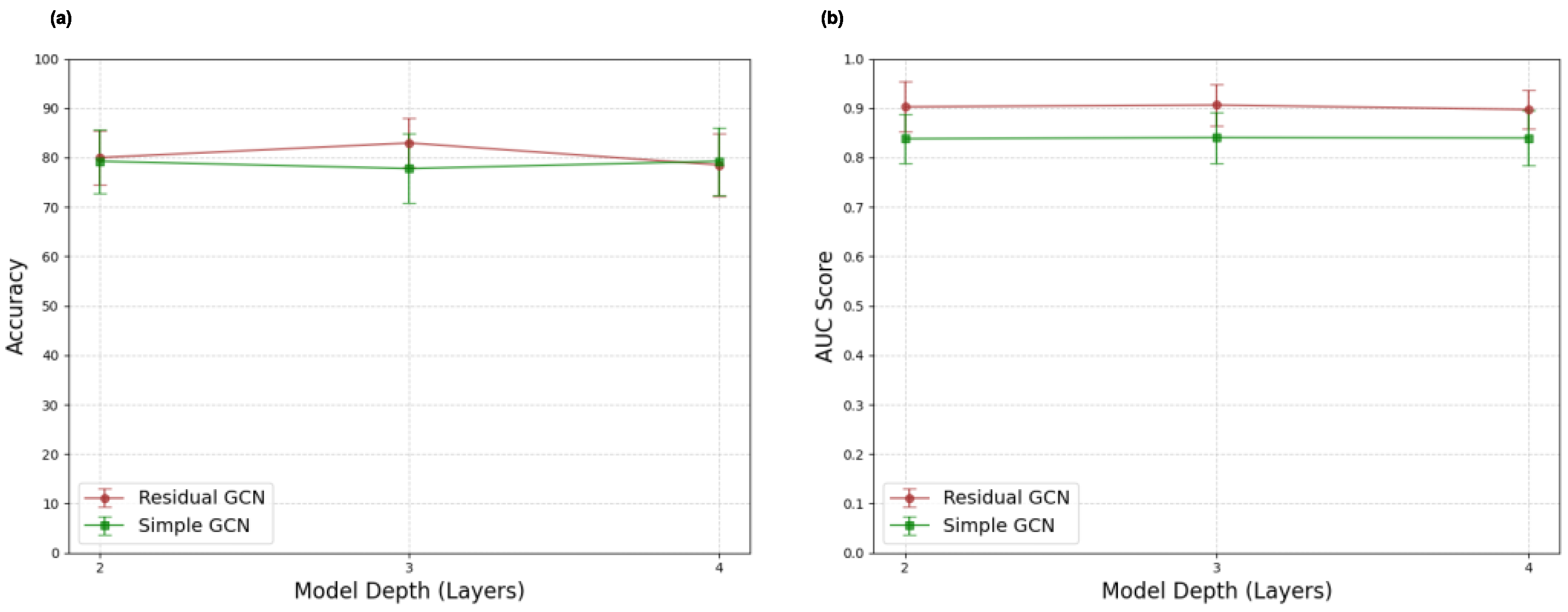
2.7. GNN Architecture Exploration Experiments
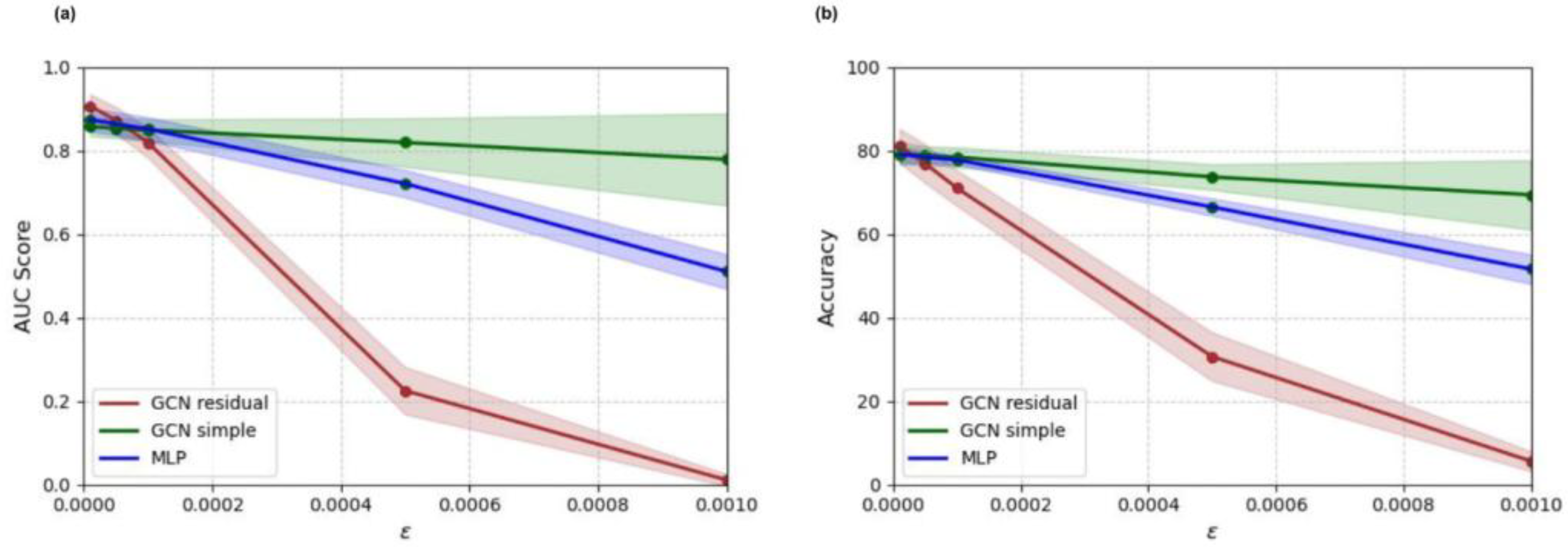
2.8. Adversarial Sensitivity Experiments
3. Results
3.1. Adult Training and Adult Testing
3.2. Adult Training and Pediatric Testing
3.3. Pediatric Training and Pediatric Testing
3.4. Adult-Enriched Pediatric Dataset Training and Testing
3.5. GNN Architecture Exploration Results
3.6. Adversarial Sensitivity Experiment Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Griffa, A.; Baumann, P.S.; Thiran, J.P.; Hagmann, P. Structural connectomics in brain diseases. Neuroimage 2013, 80, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.L.; Jahanshad, N.; McMahon, K.L.; de Zubicaray, G.I.; Martin, N.G.; Hickie, I.B.; Toga, A.W.; Wright, M.J.; Thompson, P.M. Development of brain structural connectivity between ages 12 and 30: A 4-Tesla diffusion imaging study in 439 adolescents and adults. Neuroimage 2013, 64, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Somerville, L.H.; Bookheimer, S.Y.; Buckner, R.L.; Burgess, G.C.; Curtiss, S.W.; Dapretto, M.; Elam, J.S.; Gaffrey, M.S.; Harms, M.P.; Hodge, C.; et al. The Lifespan Human Connectome Project in Development: A large-scale study of brain connectivity development in 5–21 year olds. Neuroimage 2018, 183, 456–468. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, B.C.; Chen, Z.; He, Y.; Evans, A.C.; Bernasconi, N. Graph-theoretical analysis reveals disrupted small-world organization of cortical thickness correlation networks in temporal lobe epilepsy. Cereb. Cortex 2011, 21, 2147–2157. [Google Scholar] [CrossRef]
- Gerster, M.; Taher, H.; Skoch, A.; Hlinka, J.; Guye, M.; Bartolomei, F.; Jirsa, V.; Zakharova, A.; Olmi, S. Patient-Specific Network Connectivity Combined with a Next Generation Neural Mass Model to Test Clinical Hypothesis of Seizure Propagation. Front. Syst. Neurosci. 2021, 15, 675272. [Google Scholar] [CrossRef]
- Jirsa, V.K.; Proix, T.; Perdikis, D.; Woodman, M.M.; Wang, H.; Gonzalez-Martinez, J.; Bernard, C.; Benar, C.; Guye, M.; Chauvel, P.; et al. The Virtual Epileptic Patient: Individualized whole-brain models of epilepsy spread. Neuroimage 2017, 145, 377–388. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, L.; Lin, C.P.; Zhou, Y.; Chou, K.H.; Lo, C.Y.; Su, T.P.; Jiang, T. Abnormal topological organization of structural brain networks in schizophrenia. Schizophr. Res. 2012, 141, 109–118. [Google Scholar] [CrossRef]
- Skudlarski, P.; Jagannathan, K.; Anderson, K.; Stevens, M.C.; Calhoun, V.D.; Skudlarska, B.A.; Pearlson, G. Brain connectivity is not only lower but different in schizophrenia: A combined anatomical and functional approach. Biol. Psychiatry 2010, 68, 61–69. [Google Scholar] [CrossRef]
- Yao, Z.; Zhang, Y.; Lin, L.; Zhou, Y.; Xu, C.; Jiang, T.; Alzheimer’s Disease Neuroimaging Initiative. Abnormal cortical networks in mild cognitive impairment and Alzheimer’s disease. PLoS Comput. Biol. 2010, 6, e1001006. [Google Scholar] [CrossRef]
- Wang, J.; Khosrowabadi, R.; Ng, K.K.; Hong, Z.; Chong, J.S.X.; Wang, Y.; Chen, C.Y.; Hilal, S.; Venketasubramanian, N.; Wong, T.Y.; et al. Alterations in Brain Network Topology and Structural-Functional Connectome Coupling Relate to Cognitive Impairment. Front. Aging Neurosci. 2018, 10, 404. [Google Scholar] [CrossRef]
- Lo, C.Y.; Wang, P.N.; Chou, K.H.; Wang, J.; He, Y.; Lin, C.P. Diffusion tensor tractography reveals abnormal topological organization in structural cortical networks in Alzheimer’s disease. J. Neurosci. 2010, 30, 16876–16885. [Google Scholar] [CrossRef] [PubMed]
- Tolan, E.; Isik, Z. Graph Theory Based Classification of Brain Connectivity Network for Autism Spectrum Disorder. In Proceedings of the Bioinformatics and Biomedical Engineering: 6th International Work-Conference, IWBBIO 2018, Granada, Spain, 25–27 April 2018; Proceedings, Part I 6; Springer: Cham, Switzerland, 2018; pp. 520–530. [Google Scholar]
- Li, H.; Xue, Z.; Ellmore, T.M.; Frye, R.E.; Wong, S.T. Network-based analysis reveals stronger local diffusion-based connectivity and different correlations with oral language skills in brains of children with high functioning autism spectrum disorders. Hum. Brain Mapp. 2014, 35, 396–413. [Google Scholar] [CrossRef] [PubMed]
- Tymofiyeva, O.; Yuan, J.P.; Huang, C.Y.; Connolly, C.G.; Henje Blom, E.; Xu, D.; Yang, T.T. Application of machine learning to structural connectome to predict symptom reduction in depressed adolescents with cognitive behavioral therapy (CBT). Neuroimage Clin. 2019, 23, 101914. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, K.; Amemiya, S.; Suzuki, Y.; Kunii, N.; Kawai, K.; Mori, H.; Kunimatsu, A.; Saito, N.; Aoki, S.; Ohtomo, K. Machine Learning of DTI Structural Brain Connectomes for Lateralization of Temporal Lobe Epilepsy. Magn. Reson. Med. Sci. 2016, 15, 121–129. [Google Scholar] [CrossRef]
- Chen, V.C.; Lin, T.Y.; Yeh, D.C.; Chai, J.W.; Weng, J.C. Functional and Structural Connectome Features for Machine Learning Chemo-Brain Prediction in Women Treated for Breast Cancer with Chemotherapy. Brain Sci. 2020, 10, 851. [Google Scholar] [CrossRef]
- Zhou, J.; Cui, G.; Hu, S.; Zhang, Z.; Yang, C.; Liu, Z.; Wang, L.; Li, C.; Sun, M. Graph neural networks: A review of methods and applications. AI Open 2020, 1, 57–81. [Google Scholar] [CrossRef]
- Scarselli, F.; Gori, M.; Tsoi, A.C.; Hagenbuchner, M.; Monfardini, G. The graph neural network model. IEEE Trans. Neural Netw. 2009, 20, 61–80. [Google Scholar] [CrossRef]
- Bessadok, A.; Mahjoub, M.A.; Rekik, I. Graph Neural Networks in Network Neuroscience. IEEE Trans. Pattern Anal. Mach. Intell. 2023, 45, 5833–5848. [Google Scholar] [CrossRef]
- Kazi, A.; Mora, J.; Fischl, B.; Dalca, A.V.; Aganj, I. Multi-Head Graph Convolutional Network for Structural Connectome Classification. In Proceedings of the Graphs in Biomedical Image Analysis, and Overlapped Cell on Tissue Dataset for Histopathology: 5th MICCAI Workshop, GRAIL 2023 and 1st MICCAI Challenge, OCELOT 2023, Held in Conjunction with MICCAI 2023, Vancouver, BC, Canada, 23 September, and 4 October 2023, Proceedings; Springer: Berlin/Heidelberg, Germany, 2024; Volume 14373, pp. 27–36. [Google Scholar] [CrossRef]
- Weis, S.; Patil, K.R.; Hoffstaedter, F.; Nostro, A.; Yeo, B.T.T.; Eickhoff, S.B. Sex Classification by Resting State Brain Connectivity. Cereb. Cortex 2020, 30, 824–835. [Google Scholar] [CrossRef]
- Gur, R.E.; Gur, R.C. Sex differences in brain and behavior in adolescence: Findings from the Philadelphia Neurodevelopmental Cohort. Neurosci. Biobehav. Rev. 2016, 70, 159–170. [Google Scholar] [CrossRef]
- Beacher, F.D.; Minati, L.; Baron-Cohen, S.; Lombardo, M.V.; Lai, M.C.; Gray, M.A.; Harrison, N.A.; Critchley, H.D. Autism attenuates sex differences in brain structure: A combined voxel-based morphometry and diffusion tensor imaging study. AJNR Am. J. Neuroradiol. 2012, 33, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Williamson, J.; Yabluchanskiy, A.; Mukli, P.; Wu, D.H.; Sonntag, W.; Ciro, C.; Yang, Y. Sex differences in brain functional connectivity of hippocampus in mild cognitive impairment. Front. Aging Neurosci. 2022, 14, 959394. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gao, J.; Shi, H.; Huang, B.; Wang, X.; Situ, W.; Cai, W.; Yi, J.; Zhu, X.; Yao, S. Sex differences of uncinate fasciculus structural connectivity in individuals with conduct disorder. Biomed. Res. Int. 2014, 2014, 673165. [Google Scholar] [CrossRef] [PubMed]
- Tyan, Y.S.; Liao, J.R.; Shen, C.Y.; Lin, Y.C.; Weng, J.C. Gender differences in the structural connectome of the teenage brain revealed by generalized q-sampling MRI. Neuroimage Clin. 2017, 15, 376–382. [Google Scholar] [CrossRef]
- Said, A.; Bayrak, R.G.; Derr, T.; Shabbir, M.; Moyer, D.; Chang, C.; Koutsoukos, X. NeuroGraph: Benchmarks for graph machine learning in brain connectomics. In Proceedings of the 37th International Conference on Neural Information Processing Systems, New Orleans, LA, USA, 10–16 December 2023. [Google Scholar]
- Supekar, K.; Uddin, L.Q.; Prater, K.; Amin, H.; Greicius, M.D.; Menon, V. Development of functional and structural connectivity within the default mode network in young children. Neuroimage 2010, 52, 290–301. [Google Scholar] [CrossRef]
- Mitchell, W.J.; Tepfer, L.J.; Henninger, N.M.; Perlman, S.B.; Murty, V.P.; Helion, C. Developmental Differences in Affective Representation Between Prefrontal and Subcortical Structures. Soc. Cogn. Affect. Neurosci. 2021, 17, 311–322. [Google Scholar] [CrossRef]
- Smith, S.M.; Jenkinson, M.; Woolrich, M.W.; Beckmann, C.F.; Behrens, T.E.; Johansen-Berg, H.; Bannister, P.R.; De Luca, M.; Drobnjak, I.; Flitney, D.E.; et al. Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage 2004, 23 (Suppl. S1), S208–S219. [Google Scholar] [CrossRef]
- Tournier, J.D.; Smith, R.; Raffelt, D.; Tabbara, R.; Dhollander, T.; Pietsch, M.; Christiaens, D.; Jeurissen, B.; Yeh, C.H.; Connelly, A. MRtrix3: A fast, flexible and open software framework for medical image processing and visualisation. Neuroimage 2019, 202, 116137. [Google Scholar] [CrossRef]
- Smith, R.E.; Tournier, J.D.; Calamante, F.; Connelly, A. SIFT2: Enabling dense quantitative assessment of brain white matter connectivity using streamlines tractography. Neuroimage 2015, 119, 338–351. [Google Scholar] [CrossRef]
- Glasser, M.F.; Coalson, T.S.; Robinson, E.C.; Hacker, C.D.; Harwell, J.; Yacoub, E.; Ugurbil, K.; Andersson, J.; Beckmann, C.F.; Jenkinson, M.; et al. A multi-modal parcellation of human cerebral cortex. Nature 2016, 536, 171–178. [Google Scholar] [CrossRef]
- Kipf, T.; Welling, M. Semi-Supervised Classification with Graph Convolutional Networks. arXiv 2016, arXiv:1609.02907. [Google Scholar] [CrossRef]
- Jernigan, T.L.; Brown, T.T.; Hagler, D.J., Jr.; Akshoomoff, N.; Bartsch, H.; Newman, E.; Thompson, W.K.; Bloss, C.S.; Murray, S.S.; Schork, N.; et al. The Pediatric Imaging, Neurocognition, and Genetics (PING) Data Repository. Neuroimage 2016, 124, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Casey, B.J.; Cannonier, T.; Conley, M.I.; Cohen, A.O.; Barch, D.M.; Heitzeg, M.M.; Soules, M.E.; Teslovich, T.; Dellarco, D.V.; Garavan, H.; et al. The Adolescent Brain Cognitive Development (ABCD) study: Imaging acquisition across 21 sites. Dev. Cogn. Neurosci. 2018, 32, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef]
- Hagmann, P.; Cammoun, L.; Gigandet, X.; Meuli, R.; Honey, C.J.; Wedeen, V.J.; Sporns, O. Mapping the structural core of human cerebral cortex. PLoS Biol. 2008, 6, e159. [Google Scholar] [CrossRef]
- van den Heuvel, M.P.; Sporns, O. Network hubs in the human brain. Trends Cogn. Sci. 2013, 17, 683–696. [Google Scholar] [CrossRef]
- Tomasi, D.; Volkow, N.D. Functional connectivity hubs in the human brain. Neuroimage 2011, 57, 908–917. [Google Scholar] [CrossRef]
- Collin, G.; Sporns, O.; Mandl, R.C.; van den Heuvel, M.P. Structural and functional aspects relating to cost and benefit of rich club organization in the human cerebral cortex. Cereb. Cortex 2014, 24, 2258–2267. [Google Scholar] [CrossRef]
- Hofer, S.; Frahm, J. Topography of the human corpus callosum revisited—Comprehensive fiber tractography using diffusion tensor magnetic resonance imaging. Neuroimage 2006, 32, 989–994. [Google Scholar] [CrossRef]
- Wahl, M.; Lauterbach-Soon, B.; Hattingen, E.; Jung, P.; Singer, O.; Volz, S.; Klein, J.C.; Steinmetz, H.; Ziemann, U. Human motor corpus callosum: Topography, somatotopy, and link between microstructure and function. J. Neurosci. 2007, 27, 12132–12138. [Google Scholar] [CrossRef]
- Lebel, C.; Caverhill-Godkewitsch, S.; Beaulieu, C. Age-related regional variations of the corpus callosum identified by diffusion tensor tractography. Neuroimage 2010, 52, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Jancke, L.; Staiger, J.F.; Schlaug, G.; Huang, Y.; Steinmetz, H. The relationship between corpus callosum size and forebrain volume. Cereb. Cortex 1997, 7, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Ingalhalikar, M.; Smith, A.; Parker, D.; Satterthwaite, T.D.; Elliott, M.A.; Ruparel, K.; Hakonarson, H.; Gur, R.E.; Gur, R.C.; Verma, R. Sex differences in the structural connectome of the human brain. Proc. Natl. Acad. Sci. USA 2014, 111, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, J.H.; Knickmeyer, R.C.; Gao, W. Imaging structural and functional brain development in early childhood. Nat. Rev. Neurosci. 2018, 19, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Huang, H.; He, Y. Developmental Connectomics from Infancy through Early Childhood. Trends Neurosci. 2017, 40, 494–506. [Google Scholar] [CrossRef]
- Sydnor, V.J.; Larsen, B.; Bassett, D.S.; Alexander-Bloch, A.; Fair, D.A.; Liston, C.; Mackey, A.P.; Milham, M.P.; Pines, A.; Roalf, D.R.; et al. Neurodevelopment of the association cortices: Patterns, mechanisms, and implications for psychopathology. Neuron 2021, 109, 2820–2846. [Google Scholar] [CrossRef]
- Margulies, D.S.; Ghosh, S.S.; Goulas, A.; Falkiewicz, M.; Huntenburg, J.M.; Langs, G.; Bezgin, G.; Eickhoff, S.B.; Castellanos, F.X.; Petrides, M.; et al. Situating the default-mode network along a principal gradient of macroscale cortical organization. Proc. Natl. Acad. Sci. USA 2016, 113, 12574–12579. [Google Scholar] [CrossRef]
- Hagmann, P.; Sporns, O.; Madan, N.; Cammoun, L.; Pienaar, R.; Wedeen, V.J.; Meuli, R.; Thiran, J.P.; Grant, P.E. White matter maturation reshapes structural connectivity in the late developing human brain. Proc. Natl. Acad. Sci. USA 2010, 107, 19067–19072. [Google Scholar] [CrossRef]
- Dennis, E.L.; Thompson, P.M. Typical and atypical brain development: A review of neuroimaging studies. Dialogues Clin. Neurosci. 2013, 15, 359–384. [Google Scholar] [CrossRef]
- Lebel, C.; Beaulieu, C. Longitudinal development of human brain wiring continues from childhood into adulthood. J. Neurosci. 2011, 31, 10937–10947. [Google Scholar] [CrossRef]
- Westlye, L.T.; Walhovd, K.B.; Dale, A.M.; Bjornerud, A.; Due-Tonnessen, P.; Engvig, A.; Grydeland, H.; Tamnes, C.K.; Ostby, Y.; Fjell, A.M. Life-span changes of the human brain white matter: Diffusion tensor imaging (DTI) and volumetry. Cereb. Cortex 2010, 20, 2055–2068. [Google Scholar] [CrossRef]
- Rodriguez, D.; Nayak, T.; Chen, Y.; Krishnan, R.; Huang, Y. On the role of deep learning model complexity in adversarial robustness for medical images. BMC Med. Inform. Decis. Mak. 2022, 22, 160. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | Patient Population | N | % Female |
|---|---|---|---|
| BRIGHT | Adult | 309 | 53.40 |
| HCP-D | Pediatric | 135 | 56.30 |
| MLP | GCN (Simple) | GCN (Residual) | |
|---|---|---|---|
| Number of Model Parameters | 7.37 × 107 | 3.89 × 104 | 4.78 × 106 |
| Metric | RF | SVM | MLP | GCN (Simple) | GCN (Residual) |
|---|---|---|---|---|---|
| Accuracy (%) | 73.13 ± 2.05 | 76.37 ± 5.84 | 77.66 ± 3.18 | 85.10 ± 2.84 | 82.82 ± 5.30 |
| AUC Score | 0.80 ± 0.01 | 0.86 ± 0.03 | 0.89 ± 0.01 | 0.91 ± 0.03 | 0.93 ± 0.03 |
| Metric | RF | SVM | MLP | GCN (Simple) | GCN (Residual) |
|---|---|---|---|---|---|
| Accuracy (%) | 55.70 ± 3.23 | 56.15 ± 3.79 | 53.93 ± 3.85 | 60.74 ± 3.47 | 74.96 ± 2.41 |
| AUC Score | 0.62 ± 0.01 | 0.64 ± 0.03 | 0.60 ± 0.01 | 0.71 ± 0.03 | 0.86 ± 0.03 |
| Metric | RF | SVM | MLP | GCN (Simple) | GCN (Residual) |
|---|---|---|---|---|---|
| Accuracy (%) | 66.67 ± 8.76 | 65.93 ± 8.25 | 56.30 ± 7.55 | 71.11 ± 10.84 | 66.67 ± 12.83 |
| AUC Score | 0.73 ± 0.13 | 0.80 ± 0.04 | 0.67 ± 0.05 | 0.76 ± 0.09 | 0.85 ± 0.05 |
| Metric | RF | SVM | MLP | GCN (Simple) | GCN (Residual) |
|---|---|---|---|---|---|
| Accuracy (%) | 65.93 ± 10.32 | 67.41 ± 8.25 | 72.59 ± 6.46 | 79.26 ± 6.46 | 82.96 ± 5.02 |
| AUC Score | 0.74 ± 0.01 | 0.80 ± 0.08 | 0.79 ± 0.06 | 0.84 ± 0.05 | 0.91 ± 0.04 |
| Pooling Function | Accuracy (%) | AUC Score |
|---|---|---|
| Mean | 79.26 ± 6.46 | 0.91 ± 0.05 |
| Max | 77.04 ± 7.18 | 0.85 ± 0.04 |
| Aggregation Function | Accuracy (%) | AUC Score |
|---|---|---|
| Mean | 82.96 ± 5.02 | 0.91 ± 0.04 |
| Max | 81.48 ± 5.24 | 0.90 ± 0.04 |
| Residual GCN Model (With/Without Skips) | Accuracy (%) | AUC Score |
|---|---|---|
| Without Skips | 68.89 ± 9.54 | 0.74 ± 0.07 |
| With Skips | 82.96 ± 5.02 | 0.91 ± 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srinivasan, A.; Raja, R.; Glass, J.O.; Hudson, M.M.; Sabin, N.D.; Krull, K.R.; Reddick, W.E. Graph Neural Network Learning on the Pediatric Structural Connectome. Tomography 2025, 11, 14. https://doi.org/10.3390/tomography11020014
Srinivasan A, Raja R, Glass JO, Hudson MM, Sabin ND, Krull KR, Reddick WE. Graph Neural Network Learning on the Pediatric Structural Connectome. Tomography. 2025; 11(2):14. https://doi.org/10.3390/tomography11020014
Chicago/Turabian StyleSrinivasan, Anand, Rajikha Raja, John O. Glass, Melissa M. Hudson, Noah D. Sabin, Kevin R. Krull, and Wilburn E. Reddick. 2025. "Graph Neural Network Learning on the Pediatric Structural Connectome" Tomography 11, no. 2: 14. https://doi.org/10.3390/tomography11020014
APA StyleSrinivasan, A., Raja, R., Glass, J. O., Hudson, M. M., Sabin, N. D., Krull, K. R., & Reddick, W. E. (2025). Graph Neural Network Learning on the Pediatric Structural Connectome. Tomography, 11(2), 14. https://doi.org/10.3390/tomography11020014