Abstract
The sophisticated, elegant protein-polymers designed by nature can serve as inspiration to redesign and biomanufacture protein-based materials using synthetic biology. Historically, petro-based polymeric materials have dominated industrial activities, consequently transforming our way of living. While this benefits humans, the fabrication and disposal of these materials causes environmental sustainability challenges. Fortunately, protein-based biopolymers can compete with and potentially surpass the performance of petro-based polymers because they can be biologically produced and degraded in an environmentally friendly fashion. This paper reviews four groups of protein-based polymers, including fibrous proteins (collagen, silk fibroin, fibrillin, and keratin), elastomeric proteins (elastin, resilin, and wheat glutenin), adhesive/matrix proteins (spongin and conchiolin), and cyanophycin. We discuss the connection between protein sequence, structure, function, and biomimetic applications. Protein engineering techniques, such as directed evolution and rational design, can be used to improve the functionality of natural protein-based materials. For example, the inclusion of specific protein domains, particularly those observed in structural proteins, such as silk and collagen, enables the creation of novel biomimetic materials with exceptional mechanical properties and adaptability. This review also discusses recent advancements in the production and application of new protein-based materials through the approach of synthetic biology combined biomimetics, providing insight for future research and development of cutting-edge bio-inspired products. Protein-based polymers that utilize nature’s designs as a base, then modified by advancements at the intersection of biology and engineering, may provide mankind with more sustainable products.
1. Introduction
The increase in demand for plastics has caused a major expansion in the production of plastic goods [1,2]. Every year, more than 100 million tons of plastic trash are disposed of globally, resulting in considerable environmental pollution due to the inherent resistance of plastic to natural disintegration [3,4,5,6]. The COVID-19 epidemic exacerbated the environmental problems associated with plastic wastes. Thus, it is important to reduce use of throwaway plastics [7].
Bio-sourced materials, possessing intrinsic biodegradability, are an attractive alternative to nondegradable plastic materials [8,9,10,11]. Natural materials possess the capacity to compete with synthetic polymers, particularly when they undergo appropriate processing.
Polylactic acid has gained significant attention as one alternative, due to its robust mechanical capabilities [12,13,14]. Unfortunately, degradation of polylactic acid requires a composting environment with suitable microbes to achieve degradation in a rapid timeframe [15]. Thus, scientists continue to focus on finding/developing polymers that have both exceptional mechanical qualities and sufficiently rapid degradation rates to be considered environmentally sustainable.
The protein-based polymers discussed in this study are predominantly sourced from natural products (Table 1). Proteins can be categorized according to structural and functional attributes, which determine their activities.
Given their lightweight properties and mechanical robustness, protein-based fibers demonstrate exceptional performance compared to typical artificial fibers (nylon and kevlar), particularly with regards to their unique biocompatibility, biodegradability, and sustainability [16,17,18,19]. This makes protein fibers especially favorable for biomedical uses, thereby broadening the spectrum commercial applications [20,21,22,23,24,25]. Alternative carbohydrate polymers, such as cellulose, hemicellulose, and chitosan, are described in other scholarly investigations [15,26,27].

Table 1.
Structural characteristics of major nature-occurring proteins and their bio-inspired applications.
This review is organized into eight sections. In Section 2, Section 3, Section 4 and Section 5, we review four different groups of protein-based polymers, including fibrous proteins (collagen, silk fibroin, fibrillins, and keratin), elastomeric proteins (elastin, resilin, and wheat glutenin), adhesive/matrix proteins (spongin and conchiolin), and cyanophycin. In Section 6, we review the techniques for application of protein-based materials in the post-treatments of endogenous and recombinant proteins, including the covalent crosslinking and hydrogel production. Section 7 describes novel approaches for the fabrication of protein fabrics. Section 8 gives perspectives.
2. Fibrous Proteins
Fibrous, globular, and membrane proteins are the three major classes of cellular proteins [115]. Fibrous proteins play a structural role by forming filamentous and sheet-like structures, which are typically inert and water-insoluble. Collagen, fibrin, keratin, and elastin are the four major fibrous protein families.
2.1. Collagen
Collagen is a fundamental structural component ubiquitously present in tissues [28]. It accounts for ~30% of the total protein content within the soma and serves as the principal constituent of the extracellular matrix (ECM). The primary function of ECM is to facilitate crucial structural reinforcement by augmenting tensile strength in various tissues and organs. Moreover, ECM is extremely important in providing tissue pliability, specifically for tendons [116]. Collagen is the most abundant fibrous protein in vertebrate connective tissues (tendon, cartilage, and bone) [117].
In total, 29 distinct vertebrate collagens have been found, originating from a minimum of 42 unique genes [29,118]. Collagen has a unique structural attribute known as the amino acid triplet repeat, conserved Gly–X–Y motifs (Figure 1). “Gly” denotes the amino acid glycine, and the “X” and “Y” positions can accommodate any amino acid residue, however, proline and hydroxyproline are frequently found in these positions. Collagen types exhibit significant variations in their amino acid sequences, leading to distinct chemical and physical properties. Collagen types I to V are the most abundant and have been extensively studied for their biomedical applications [118]. Not all these collagens are part of the ECM.
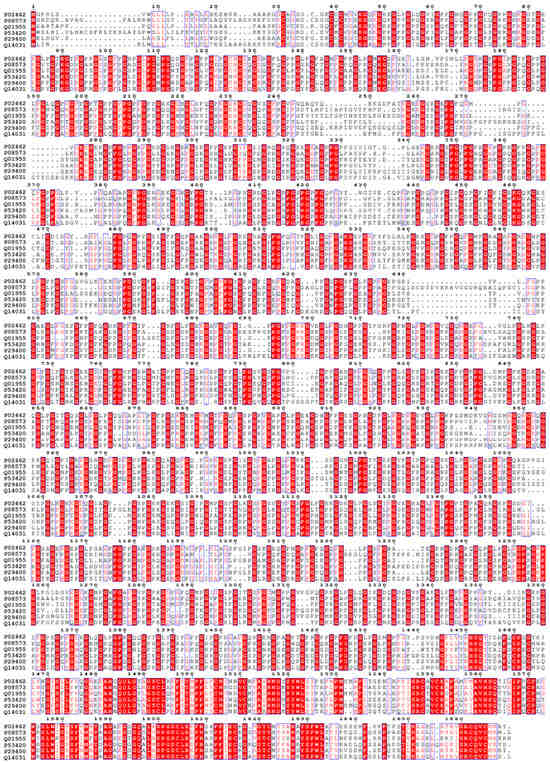
Figure 1.
Multiple protein sequence alignment of Collagen type IV: Alpha 1–6 subunits shown the conserved G-X-Y repeated motifs.
Collagens are classified based on their α-chain composition and supramolecular structure. Fibrillar collagens possess significant quantities of continuous triple-helix structures (Gly-X-Y repeat motifs). On the other hand, α-chains have the potential to possess varying quantities of non-collagenous domains, hence introducing the possibility of interfering with the helical conformation.
Collagen type IV consists of six genetically distinct isoforms (Alpha 1–6 subunits). Their sequence alignment is shown in Figure 1 and each subunit’s structure shown in Figure 2 as predicted by AlphaFold [119,120]. Type IV collagen serves as a key constituent of basement membranes.
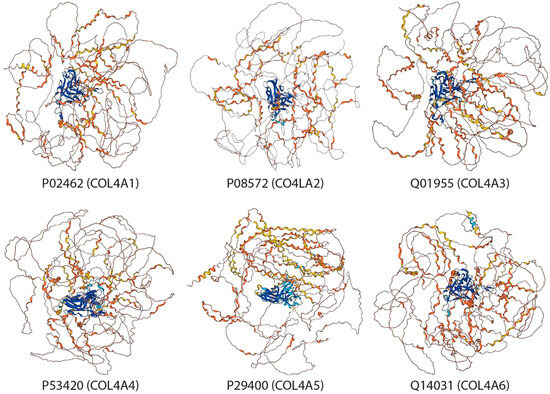
Figure 2.
Collagen type IV Alpha 1–6 subunits’ structures predicted by AlphaFold Monomer v2.0 pipelne. The predicted local distance difference test (pLDDT) is a per-residue measure of local confidence. It is scaled from 0 to 100, with higher scores indicating higher confidence. Color legend: Very high (pLDDT > 90); High (90 > pLDDT > 70); Low (70 > pLDDT > 50); Very low (pLDDT < 50).
Various forms of collagens are involved in the formation of diverse supramolecular architectures, such as geometric networks, membranous fibrils, and beaded filaments. Multiple kinds of collagen are present in many tissues, and they can co-distribute with diverse structural classes [30,31]. The presence of structural heterogeneity is associated with an increase in functionality. Various forms of collagens play significant roles in diverse biological processes and the inhibition of cancer growth [32].
2.2. Silk Fibroin
Several applications have been developed for silk due to its superior mechanical strength, which exceeds that of collagen [121]. Arthropod silk from arachnids, insects, and myriapods has potential to be an exceptional biomaterial. Silkworm domestication can be traced back more than 5000 years, as indicated by the oldest references originating from Chinese regions [122,123]. The Bombyx mori species has undergone significant domestication and cultivation in industrial farms owing to its capacity to generate superior silk that is well suited for textile applications [124]. The material exhibits several attributes, such as robustness, sheen, flexibility, elasticity, and the ability to establish chemical dye linkages. Moreover, its medical application for suturing wounds has also been practiced. The silk market exerts a significant influence on the worldwide textile economy, [75] with a total production of about 120,000 tons per annum of silk, primarily concentrated in Asia [125].
Due to the abundance of silk produced by Bombyx mori through sericulture, much research has been done on this subject. On the other hand, silk obtained from alternative arthropod sources, such as spiders, has not received the same degree of research interest. Arachnid species are deemed unsuitable for cultivation in controlled situations due to their predatory tendencies. Nevertheless, the silk produced by these organisms exhibits a distinctive amalgamation of robustness, high tensile strength, and excellent extensibility [34,73,74,126] surpassing that of other high-performance synthetic fibers. Due to their exceptional biocompatibility and biodegradability, the fibers are especially suitable for biological usage and exhibit extraordinary mechanical properties [23,36,127]. For instance, spider silk has been used in fishing lines in previous times, and more recently as microsutures [128,129]. In 2012, artist Simon Peers and entrepreneur Nicholas Godley created a cloak with a naturally golden hue, crafted from the silk of around 1.2 million Nephila spiders. The showing of this cloak in London had a significant role in establishing the silk’s status as a luxury item [130].
Researchers have been motivated by the remarkable mechanical attributes of natural spider silks, leading them to explore the creation of biomimetic protein fibers by recombining spider silk proteins [23,33]. Synthetic biology and recombinant DNA technology have facilitated the research and production of many variants of spider silk proteins (known as spidroins) [131,132]. Various spinning processes have been developed in the field of biological silk production [133,134,135,136,137,138]. The preservation of the hierarchical organization of recombinant spidroins exhibits limitations, which lead to missing functional domains. As a result, a considerable proportion of recombinant protein fibers demonstrate mechanical characteristics which are inferior when compared to innate spider silk. The examples in nature encompass mussel byssus [139,140], bagworm silks [141,142], sandcastle worm glue [143], and squid ring teeth [144,145]. The extensive range of patterns and shapes exhibited by these native proteins presents numerous possibilities for development of protein fibers possessing exceptional mechanical strength. Therefore, developing native structural proteins as biomimetic fibers is particularly attractive.
To better understand the mechanical properties of silk protein fibers, researchers have investigated the relationship among patterns of amino acids, structures, and mechanical attributes [35]. Fiber properties are affected by the arrangement of hydrophobic and hydrophilic domains within protein sequences that are commonly characterized by repetition. The presence of polyalanine within the hydrophobic zone is a contributing factor to high tensile strength, as indicated by previous studies [37,146]. Hydrophilic glycine/proline-rich areas facilitate hydrogen bonding between crystalline β-sheets. This interaction has a role in regulating fiber elasticity [147]. Silk fibers exhibit remarkable mechanical characteristics, rendering them exceedingly appropriate for the development of biomaterials employed in wound suturing, tissue regeneration, optical devices, biosensing systems, and drug administration endeavors (illustrated in Figure 3 [6,148,149]).
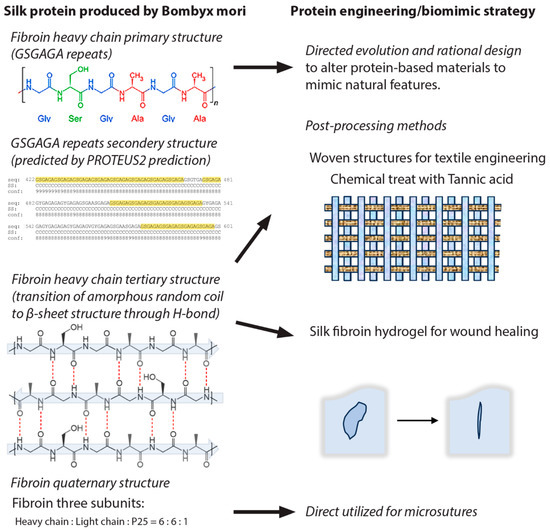
Figure 3.
Schematic illustration of the naturally occurring protein-based biomaterial structures and their application, using silk fibroin as an example. (Left) Silk fibroin produced by the silkworm Bombyx mori consists of a heavy chain, a light chain, and a glycoprotein, P25. (Right) Examples of utilizing silk fibrin for various applications, including the silk fibroin hydrogel.
Silk protein fiber is composed of two components, fibroin and sericin [150,151]. The production of silk proteins occurs within epithelial cells that line certain glands of the silkworm. Subsequently, the proteins are discharged into the cavity of the gland and further converted into fibrous structures [152,153,154]. Glycine-X repeats make up the crystalline domains of silk fibroin, where X stands for the amino acids alanine, serine, threonine, and valine [155]. Fibroin is a dimer, consisting of a thin chain weighing approximately 26 kilodaltons and a thick chain weighing roughly 390 kilodaltons. There are identical numbers of both of these chains, and a single disulfide bond connects them [156]. The sericins are a class of sticky proteins that facilitate the binding and encapsulate of the fibroin fibers within silk. Sericins have a molecular weight range between 20 and 310 kDa [157,158] and are comprised of 44% glycine, 29% alanine, and 11% serine [159]. A bacterium bearing the genes from B. mori has been used to overexpress fibrion [101,160,161] and sericin proteins [162,163].
There are two different types of silk fibroin in its solid state. Silk I is the water-soluble initial state before crystallization and spinning. Silk I is then converted by exposure to heat, organic solvents or physical manipulation into silk II, which is the secondary structure also called a β-sheet [164,165]. The water solubility of the silk II structure is limited. However, it can be effectively dissolved in several chaotropic agents [34,166]. The β-sheets are stacked in anti-parallel alignment, with hydrogen bonds both inside and across chains and the influence of van der Waals force [156,167,168]. This structure provides high thermodynamic stability as verified using FTIR and circular dichroism measuring techniques [169].
Fibroin has found extensive application in medicinal products. The extraction of fibroin from the silkworm cocoon normally involves the removal of sericin, followed by a subsequent purification process [170]. There are numerous ways to extract and purify the protein from silk fibroin. The extraction of sericin, usually referred to as “degumming”, is frequently achieved using Na2CO3 boiling and autoclaving procedures. The degummed silk is next dissolved in a strong lithium bromide solution to separate the fibroin [171,172], or occasionally in a ternary solvent system comprising CaCl2, CH3CH2OH, and H2O [173,174]. Following the process of solvent evaporation, fibroin can undergo additional purification [159,175,176,177]. The potential of silk proteins for many biomedical uses is noteworthy owing to their capability to undergo easy processing in water or other solvents, resulting in the creation of gels, fibers, or sponges that can be chemically modified in a diverse manner. Furthermore, it is worth noting that these proteins exhibit notable biocompatibility, enzymatic degradability, and mechanical resilience as mentioned in previous studies [178,179].
Hydrogels can be formed via the sol–gel transition in reconstituted silk fibroin (SF) solutions, which can be triggered by the addition of acid, ions, or other supplementary substances [38,180,181,182,183,184]. Temperature, SF concentration, and pH are only a few examples of the variables that affect the gelation process. In general, gelation time exhibits a decreasing trend when the concentration of SF is increased, as well as when the temperature is elevated. Moreover, previous research has demonstrated a correlation between a reduction in pH and a corresponding decrease in the time required for gelation [182,184]. The hydrogel’s pore size diminishes while its mechanical strength and stiffness enhance as the concentrations of SF or gelation temperature increase [182]. The gelation period in silk fibroin can be controlled by adjusting the power output, duration of sonication, and concentration of the silk fibroin. This enables the manipulation of the gelation period, spanning from minutes to hours. Ultrasonication can enhance the creation of sheets. This, in turn, results in accelerated physical crosslinking and enhances the stability of gels. Conversely, subjecting SF hydrogels to vortexing results in a reduction in the gelation rate, hence prolonging the gelation process from a period of minutes to hours.
2.3. Fibrillins
Fibrillins are large, cysteine-rich glycoproteins, abundant in the ECM and distinguished by their large molecular weight of ~350 kilodaltons. Fibrillin consists of three members in humans: fibrillin-1, -2, and -3. They are the major structural components of microfibrils that are ubiquitously distributed in connective tissues. About 10% of completely grown elastic fibers are microfibrils, which are multiprotein fibrils. Fibrillin-1 is expressed throughout a human’s full lifespan, whereas fibrillin-2 and -3 are thought to be primarily present in early human development [40,41,42,185,186]. Fibrillin-2 is typically found in embryonic tissues but scarcely in adult skin, yet a marked increased expression was found in wound healing and sclerotic skin tissues [185].
All three fibrillins (≈350-kD) have similar modular organization that consists of 46 to 47 epidermal growth factor (EGF)-like domains (42 to 43 out of these are calcium-binding type cbEGF interspersed with seven 8-cysteine-containing TGF-β-binding (TB) modules found in LTBPs [43,187]. During the process of elastic fiber construction, microfibrils that are rich in fibrillin temporarily interact with various matrix proteins. The formation of fibrils with a thickness of around 10–12 nm is caused by the alignment of fibrillin monomers in a head-to-tail configuration. The fibrils have a beaded-string morphology characterized by the presence of beads separated by an average distance of roughly 56 nm. Additionally, it should be noted that the bead shoulder area of the fibrils consists of two symmetrical “arms” [44]. Calcium is of utmost importance in facilitating proper organization [45]. Microfibrils have traditionally been recognized as structural and organizational frameworks for elastin, as evidenced by their presence during embryonic development and their co-localization with tropoelastin [46]. The outer areas are the primary location for microfibrils in mature elastic fibers [47]. The functions of fibrillin microfibrils are distinct and autonomous, operating independently from their primary interaction with elastin. These functions entail enhancing the integrity of the ciliary zonules inside the ocular structure.
The maintenance of tissue structural integrity and flexibility is reliant upon the existence of collagen and fibrillins (elastic fibers). Tissues that possess collagen or elastic fibers frequently experience varying degrees and durations of deformations and provide mechanical resilience through unique mechanisms. The strength of collagen arises from its highly structured semi-crystalline molecular structure, which adopts a triple helical conformation. Nevertheless, the primary source of the material’s elasticity stems from the increased disorder of the polypeptide chains while in a relaxed condition and not subjected to stretching.
2.4. Keratins
Keratins are essential constituents of many biological structures, such as scales, hair, nails, feathers, horns, claws, hooves, and the outer layer of skin among vertebrates [188]. The silk fibroins produced by insects and spiders are often classified as keratins. Keratin has been used in wound healing, tissue engineering, and drug delivery [189]. The human genome has 54 functional annotated keratin genes (28 type I and 26 type II) [190]. The regulation of keratin production is affected by a diverse array of growth factors and cytokines, totaling more than 30 [191,192]. The formation of keratin fiber entails the development of a sturdy structure by employing covalent connections. Crosslinking can occur in two distinct manners: intermolecularly, which involves the bonding between separate polypeptide chains, and intramolecularly, which occurs inside a single polypeptide chain at different sites [49,193].
Keratins can be categorized into three discrete classes, specifically α-keratins, β-keratins, and γ-keratins. The α-keratins have an alpha-helical tertiary structure predicted by AlphaFold [117,118]. The predicted tertiary structures of type I keratin and type II keratin from human hair are shown in Figure 4. The average molecular mass typically ranges 60–80 kDa. The α-keratins possess the capacity for self-assembly, leading to the creation of elongated filamentous fibers that exhibit high tensile strength and great stretchability without experiencing breaking [189,194]. The β-keratins are primarily protective and form most of the cuticle. β-keratins do not form useful reconstituted structures.
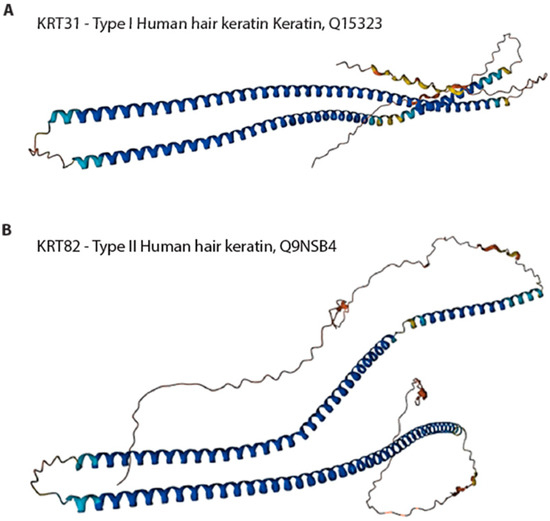
Figure 4.
AlphaFold v2.0-predicted tertiary structure of type I keratin KRT 31 (A) and type II keratin IKRT 82 (B) from human hair. Gene ID: Q15323 (KRT31); Q9NSB4 (KRT82).The predicted local distance difference test (pLDDT) is a per-residue measure of local confidence. It is scaled from 0 to 100, with higher scores indicating higher confidence. Color legend: Very high (pLDDT > 90); High (90 > pLDDT > 70); Low (70 > pLDDT > 50); Very low (pLDDT < 50).
The extraction of β-keratins is difficult [195]. The γ-keratins are globular, high in sulfur content, with a molecular mass of approximately 15 kDa. They demonstrate a compact, spherical conformation and are characterized by their heightened concentrations of cysteine, glycine, and tyrosine residues. The γ-keratins function as disulfide crosslinkers, holding the cortical superstructure together, which in turn enhances cohesion. The cross-linking mechanism described above [49,193] is responsible for the mechanical toughness and inertness seen in the cortical structure of hair and wool [196].
A variety of methodologies have emerged for the extraction of keratin. Both oxidative and reductive solvents have been employed in disrupting the disulfide crosslinks between cysteine residues, leading to the transformation of keratins into their non-crosslinked condition. Keratose is a protein combination that is acquired by employing oxidative solvents, facilitating the conversion of cysteine into cysteic acid. The protein combination obtained using reductive solvents, which maintain the cysteines in their native shape and facilitate the production of further crosslinks, is commonly known as kerateines. Keratose obtained using oxidative extraction methods, such as peracetic acid or hydrogen peroxide, demonstrates hygroscopic properties, water solubility, and the ability to form non-disulfide crosslinks, and susceptibility to hydrolytic breakdown in very acidic or alkaline environments. Biomaterials produced from keratose demonstrate a substantially accelerated disintegration rate when placed in an in vivo environment, often taking place within a time span that varies from a few days to a few weeks [197]. In contrast, biomaterials derived from kerateines using a reductive extraction method exhibit a prolonged duration of biocompatibility, as evidenced by their capacity to persist within the organism over a span of weeks to months. The reduced solubility of keratins in water-based solutions and their increased ability to withstand extreme pH conditions can be attributed to the oxidative coupling capability of cysteine groups, leading to the formation of additional crosslinks. Most frequently, sodium disulfite, 2-mercaptoethanol, or dithiothreitol are used in the reductive extraction technique [48,198,199,200,201,202].
The intrinsic self-organization of keratin solutions has been the subject of extensive research, encompassing both the small-scale and large-scale domains [203,204,205]. This phenomenon results in development of a three-dimensional fiber network, distinguished by a uniform configuration of fibers and porosity [206]. Films, sponges, and hydrogels are just a few examples of biomaterials made from keratin. In the early 1970s, attempts were made to create gels, films, and scaffolds using keratin protein solutions [207]. Blanchard was granted a patent for their use of a keratin hydrogel derivative of human hair to enhance the process of wound healing. To achieve the desired characteristics, the hair underwent an initial oxidation process using peracetic acid to synthesize hydrogels with these specific features. To cleave the disulfide bonds within the protein, the protein suspension was exposed to 60 °C for 4 h. Following this, the suspension was permitted to cool to ambient temperature, resulting in the breaking of disulfide bonds and the consequent synthesis of cysteine groups. This methodology improved protein solubility. Therefore, the development of a keratin-based hydrogel using only disulfide and hydrogen bonds seems to be a feasible strategy, eliminating the need for additional crosslinker chemicals.
2.5. The Development and Utilization of Fibrous Proteins
Fibrous proteins, which are alternatively referred to as scleroproteins, comprise a substantial category of proteins, alongside globular and membrane proteins. Collagen, elastin, and keratin are proteins that exhibit the ability to produce durable and flexible fibers, hence providing structural support for a wide range of tissues and organs [208,209,210]. Numerous animals, including silkworms, spiders, and insects, use fibrous proteins, such as silk fibroins found in cocoons, spidroins in spider webs, and resilins in insect tendons [208]. These proteins demonstrate a shared characteristic of repeating motifs in their sequences. Furthermore, the self-assembly of these proteins has an intrinsic attribute that facilitates a hierarchical self-organization, leading to the formation of fibrous structures characterized by well-defined mechanical properties [210]. The biocompatibility and mechanical resilience of these protein fibers make them particularly desirable in a range of medical and technical applications [211,212,213]. Furthermore, the engineering of recombinant protein variations enables the adjustment of multiple parameters, such as protein sequence, molecular sizes, and charge. The modifications have a significant impact on the ability to convert proteins into various configurations, such as particles, capsules, foams, nonwoven meshes, films, or hydrogels. The versatile structures of fibrous proteins have a wide range of applications in various fields such as medication delivery, tissue regeneration, optical systems, bioelectronics [210,213] and in biomaterial industries such as biofibers [214], bioplastics [215], biorubbers [216,217] used in textile innovations, environmental solutions, and biomedical applications [218].
3. Elastomeric Proteins
3.1. Elastin
Elastin is a stretchy protein that resembles a rubber band, which enables many tissues to sustain repetitive stretching or contracting and thereafter resume their shape upon the cessation of the deforming action [54]. Elastin is predominantly found within the ECM of vertebrates, except for lamprey and hagfish, which belong to the agnathans taxonomic group [56,219]. Owing to its extensive crosslinked structures, elastin is a long-lived protein that degrades slowly in healthy tissues and has a half-life of about 70 years [57,58]. The elastic fibers that are generated during embryonic development are required to endure repetitive stretching and recoiling over the course of an individual’s lifespan without experiencing permanent deformation or structural collapse [59].
Tropoelastin is the soluble precursor of elastin with a 60 kDa molecular mass, which can be characterized by the existence of two or three lysine domains. Domains of the KA type, exemplified by the sequence AAKAAKA, frequently demonstrate a recurring pattern in which lysine residues are surrounded by alanine residues. Lysine residues are also present inside KP-type domains, which are distinguished by a sequence that is abundant in proline and glycine, shown by the sequence PGAGVKPGKGP. The motifs of KA-type or KP-type form a helical structure that is left-handed and lacks intramolecular hydrogen bonding [60,61,62,63].
The flexible attributes of elastin are evident from its capacity to undergo substantial elastic deformation, as seen by its elastic modulus range of 0.3–1 MPa. Elastins exhibit a remarkable ability to undergo significant elongation with minimal application of stress. Elastin demonstrates the highest degree of linear elastic extension compared to all other documented biological substances, with a minimum extension threshold of 150% [220]. Moreover, this phenomenon exhibits a noteworthy degree of resilience, approximated at 90%, and exemplifies excellent longevity, as substantiated by the limited turnover throughout an individual’s lifespan [57,58]. Nevertheless, it should be noted that elastin exhibits a modest tensile strength of approximately 2 MPa.
It is crucial to underscore that the elasticity of elastin fibers is contingent upon their state of hydration. In the absence of moisture, elastin undergoes a transformation that renders it inflexible and susceptible to breakage [221]. The existence of water molecules within the polypeptide backbone has a significant impact in preserving the structural flexibility and disorder observed in the tropoelastin monomer, which is characterized by a state of high entropy. This phenomenon is widely acknowledged as a pivotal catalyst for the elastic recoil process [63,222,223]. Despite being in a disordered condition, tropoelastin monomers exhibit significant aggregation [63]. Proteins comprised of a significant percentage of non-polar amino acids are anticipated to exhibit an increased propensity for the establishment of a hydrophobic core, hence eliciting a repulsive effect towards water molecules. The core region exhibits a densely packed configuration of secondary structural elements, which serves as an efficient shield to safeguard the non-polar side. Elastin has distinctive properties due to its hydrophobic monomer, which consists predominantly of four non-polar amino acids (proline, glycine, valine, alanine), accounting for around 80% of its composition. The monomer exhibits notable disorder and flexibility when present in a solution [61,224,225,226].
The mobility of the elastin backbone remains preserved in both the aggregated condition [46] and fully grown cross-linked fibers [227]. The presence of structural instability in hydrophobic elastin domains and other elastomeric proteins can be ascribed to the significant proportion of elastic glycine and proline deposits. To meet the specified criteria, it is necessary for elastin to have a minimum proportion of 60% glycine and proline residues combined. Additionally, it is imperative that for every glycine residue, there are at least two proline residues [63]. The existence of these Gly and Pro residues hinders the development of elongated secondary structures. This phenomenon mostly arises due to the energetic disadvantage associated with the confinement of glycine residues within a certain structural arrangement. According to previous research [62], elastin sequences exhibiting hydrophobic properties and an average proline spacing exceeding eight are prone to β-sheet aggregation. The process of aggregation can result in the development of fibrils that resemble amyloid structures [228,229,230]. The presence of disorganized monomers and aggregates provides evidence for the entropic force that facilitates elastic recoil and the manifestation of rubber-like elasticity [231].
The sequence alignment for human 13 elastin isoforms is shown in Figure 5. The human elastin K structure shown in Figure 6 was generated by the AlphaFold [119,120].
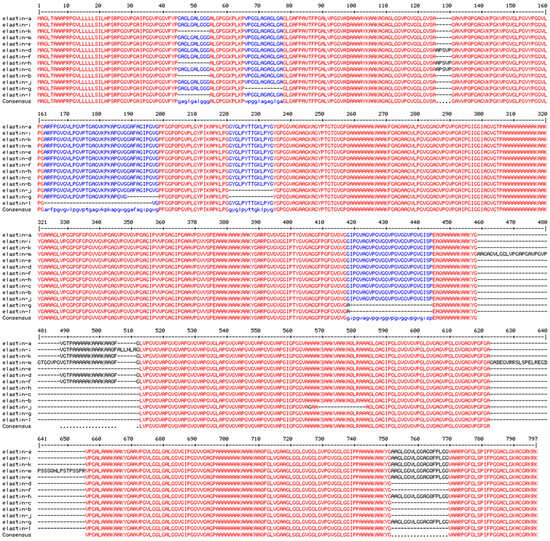
Figure 5.
Sequence alignment for 13 human elastin isoforms (a-m).
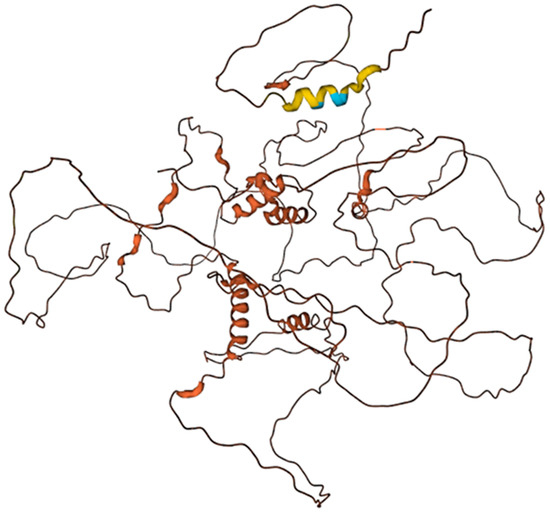
Figure 6.
Human elastin K (E7EN65) structure predicted by the AlphaFold v2.0. The predicted local distance difference test (pLDDT) is a per-residue measure of local confidence. It is scaled from 0 to 100, with higher scores indicating higher confidence. Color legend High (90 > pLDDT > 70); Low (70 > pLDDT > 50); Very low (pLDDT < 50).
3.2. Resilin
Resilin is an elastomeric protein found in the exoskeletons of insects and arthropods. Its rubber-like elasticity is what allows insects to jump long distances, fly, and vocalize [64,65]. Resilin, one of the most flexible elastomeric proteins known, stores and releases energy due to its unique structure [232,233]. The estimated elasticity of resilin in dragonfly tendons is between 600 and 700 kilopascals (kPa). Resilin contains two unique amino acids (di- and tri-tyrosine), which give it a characteristic sapphire-blue fluorescence in ultraviolet light. Exceptionally resilient, resilin can elongate up to three times its initial length before reaching the point of breakage [234].
A recently identified Drosophila melanogaster proresilin has glycine-rich repetitive sequences; the N- and C-terminal regions of the protein contain 18 repeats of a 15-residue sequence (SDTYGAPGGGNGGRP) and 11 repeats of a 13-residue sequence (GYSGGRPGGQDLG) [235], respectively. Conformational studies discovered the coexistence of two main features: folded β-turns and (quasi)extended structures (e.g., poly-L-proline II conformation). These features are common to other elastomeric proteins, suggesting a shared elasticity mechanism for resilin and other elastomeric proteins [235]. In addition to elasticity, resilin also lends itself well to fiber formations. A great tendency of resilin to aggregate in fibrous structures has been observed, particularly for the resilin-inspired polypeptide (PGGGN)10. This is encouraging for the development of resilin-based biomaterials to produce biocompatible medical devices, as well as high performing elastic materials. The sequences of resilin A and B from D. melanogaster are shown in Figure 7. The resilin A structure was predicted by AlphaFold and is shown in Figure 8.

Figure 7.
Sequence alignment for resilin isoform A (NP_611157.1, 620 aa) and isoform B (NP_995860.1, 575 aa) missing 45 aa (aa341–385) from D. melanogaster.
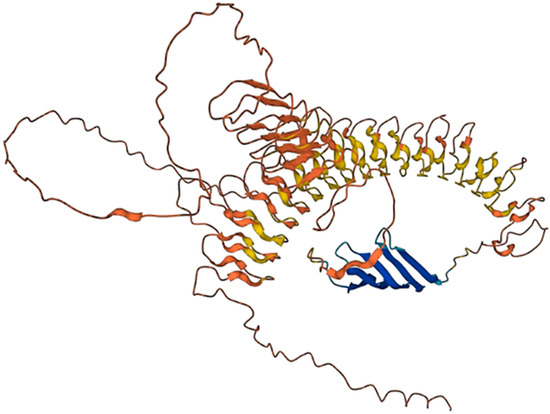
Figure 8.
D. melanogaster resilin isoform A (NP_611157.1, 620 aa) structure predicted by AlphaFold v2.0. The predicted local distance difference test (pLDDT) is a per-residue measure of local confidence. It is scaled from 0 to 100, with higher scores indicating higher confidence. Color legend: Very high (pLDDT > 90); High (90 > pLDDT > 70); Low (70 > pLDDT > 50); Very low (pLDDT < 50).
3.3. Wheat Glutenin
Bioplastics can be made from proteins [236,237] from different sources. For example, wheat gluten [71,72,97] and casein show promising properties as a raw material for biodegradable polymers. Glutenin proteins consist of high-molecular-weight (HMW-GS) and low-molecular-weight (LMW-GS) glutenin subunits [72]. Wheat HMW and LMW glutenin structures predicted by the AlphaFold are shown in Figure 9. These GS are known to polymerize in the bread-making processes, forming one of the largest polymers in nature [73]. Wheat glutenin has been used in developing bioplastics [73,98].
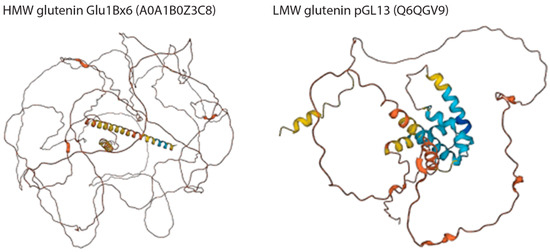
Figure 9.
Wheat HMW and LMW glutenin structures predicted by AlphaFold v2.0. The predicted local distance difference test (pLDDT) is a per-residue measure of local confidence. It is scaled from 0 to 100, with higher scores indicating higher confidence. Color legend: Very high (pLDDT > 90); High (90 > pLDDT > 70); Low (70 > pLDDT > 50); Very low (pLDDT < 50).
Unlike wheat, N2-fixing cyanobacteria can use N2 gas as a sole nitrogen source to produce large quantities of proteins. Therefore, the Zhou laboratory at South Dakota State University is interested in engineering N2-fixing cyanobacteria to produce wheat glutenin: HMW-GS and LMW-GS to make biodegradable, proteinaceous plastics. To do so, the coding sequences for hmw-GS [238] and lmw-GS ((GenBank: AFU48612.1) will be codon-optimized and chemically synthesized at Integrated DNA Technologies (IDT). The chemically synthesized genes will be cloned into a cyanobacterial expression vector pZR1188 [239] to construct the cargo plasmid. Next, the cargo plasmid will be conjugatively transformed into Anabaena sp. PCC 7120 [240] for overproduction of the recombinant HMW-GS & LMW-GS proteins.
4. Adhesive/Extracellular Matrix Proteins
4.1. Spongin
Spongin is a modified type of collagen. It is believed to have emerged around 800–900 million years ago, during the emergence of multicellular animals. The poor solubility of natural spongin in acids and enzymatic treatments hinders its unequivocal identification as collagen, keratin, or a glycosylated derivative [84,241]. Spongin is responsible for the formation of 3D structured fibrous skeletons and remains an enigma of complex chemistry in water-insoluble structural proteins [241].
Spongin plays a crucial role as specialized templates for extreme biomimetics. It consists of biopolymers that exhibit high resistance to chemically harsh and thermally extreme environments. Renewable, chemically, and thermally durable biopolymers have not yet been used to fabricate novel biologically inspired composite materials. Such materials have potential applications in the most challenging environments of modern industry, including high-volume manufacturing. Recent research has shown that it is possible to create thermostable biopolymeric scaffolds using renewable structural biopolymers, such as proteinaceous spongin and aminopolysaccharide chitin. More precisely, the Hippospongia communis spongin scaffold was coated with TiO2 using an advanced biomimetic method. This novel biocomposite successfully eliminated C.I. Basic Blue 9 by means of adsorption and photocatalysis [85]. Furthermore, the synthesis of hematite (α-Fe2O3) was achieved using hydrothermal synthesis, employing the scaffold of H. communis as a template. The combination of hematite and spongin was proven to enhance the electrochemical properties of the capacitor electrode [86].
Due to their intricate 3D fibrous network structure and exceptional sorption properties, spongin scaffolds are a very suitable matrix for the immobilization of enzymes. H. communis spongin was studied as a model for immobilizing Candida antarctica’s lipase B (CALB). Surprisingly, even after being stored at 4 °C for 20 days, this biocatalytic system continued to show its efficacy. The immobilized lipase successfully facilitated the transformation of triglycerides into glycerol and fatty acid methyl esters, which holds great potential for the biofuel industry. Further studies were focused on the immobilization of enzymes found in the matrix of sponges [242], for example, the immobilized laccase enzyme from the Trametes versicolor mushroom substantially facilitated the degradation of bisphenols, which are dangerous compounds used in the manufacturing of polycarbonates [88]. In a separate study, a distinct biocomposite composed of H. communis spongin and iron phthalocyanine effectively eliminated several contaminants, including phenol, chlorophenol, fluorophenol, and bisphenol A [81]. In addition, researchers created a scaffold of a 3D carbonized spongin-Cu/Cu2O utilizing H. communis. This scaffold effectively catalyzed the conversion of the toxic compound 4-nitrophenol into 4-aminophenol [243]. The use of spongin-based scaffolds generated from the marine demospongiae Haliclona sp. has proven to be successful in the preconcentration and extraction of compounds such as ketamine [244] and fenitrothion [82].
4.2. Conchiolin
Conchiolin is a tough, insoluble protein secreted by mollusks, forming a major organic component of the shell with deposited calcium carbonate. Hydrolyzed conchiolin protein, obtained from pearl shells, is a frequently used cosmetic ingredient in hair and skin conditioning products [91,245]. Latire et al. analyzed shell extracts derived from the marine bivalve Pecten maximus [92]. These extracts were categorized as water soluble (WS), acid soluble (AS), and acid insoluble (AI) [93]. Following a 24 h period of incubation, AS significantly increased the metabolic activity of human fibroblasts. In addition, extracts from the mussels Mytilus edulis (AS and WS) and the oyster Crassostrea gigas (AS) were found to enhance metabolic activity and cell proliferation in primary human skin fibroblasts. These findings indicate the potential medical applications of these matrix proteins or their extracts, specifically for treating various skin conditions and promoting wound healing. Indeed, a study conducted on live rats with dorsal skin wounds shown that the gradual application of an ointment comprising powdered shells of Megalobulimus lopesi resulted in a reduction of the wound [89].
5. Cyanophycin
Cyanophycin is a natural biopolymer produced by a wide range of bacteria and is the third known naturally occurring amino-acid-based polymer, the other two being polyglutamate and polylysine [246]. Cyanophycin [247] is an N-rich reserve biopolymer in cyanobacteria consisting of equimolar amounts of aspartate and arginine residues, polymerized through non-ribosomal peptide synthesis. This polymerization is catalyzed by cyanophycin synthetase (CphA1), a unique amino acid polymerase [248,249]. Arginine is the most common side chain linked to the poly-aspartate backbone; however, lysine, ornithine, and citrulline have also been identified in cyanophycin produced in heterologous hosts [250].
Accumulation of Cyanophycin Granule Peptide (CGP) forms amorphous granular structures in the cytoplasm that are visible by light microscopy. Expressing CphA1 of Anabaena sp. PCC 7120 in S. meliloti led to a 2.8-fold increased accumulation of CGP, with the CGP content reaching 43.8% (w/w) of cell dry weight in recombinant cells of S. meliloti [251].
A single amino acid replacement in cyanobacterial PII signalling protein (I86N) generated a PII mutant (PII-I86N) strain Synechocystis-BW86 that accumulated up to 15.6% CGP by dry weight (DW), while CGP is barely detectable in the wild-type [252,253]. Phosphate or potassium starvation further increased the CGP content in strain BW86 up to 47.4% or 57.3% per DW, respectively [254].
Proposed applications of cyanophycin include bioplastic [115] and neutraceuticals [111]. Cyanophycin can form a poly-Asp polymer when arginine is cleaved from the poly-Asp backbone. Poly-Asp has potential uses in biodegradable plastics [105,115] or a possible substituent for polyacrylates [255,256]. Research on microbial production of cyanophycin has been well documented [102,256,257]. Many cyanobacteria are noted for accumulating CPG as a N-rich reserve polymer [258], but only N2-fixing cyanobacteria offer the opportunity to sustainably produce this N-rich biopolymer using sunlight and N2 gas as a sole nitrogen source. To genetically engineer N2-fixing cyanobacteria to produce a high yield of cyanophycin, Zhou’s lab will create a PII mutant of Anabaena sp. PCC 7120 using previously developed molecular genetic approaches [239,259,260,261,262,263,264,265].
Cyanophycin-Based Biopolymers
Since aspartic acid is a component of the cyanophycin backbone, it is theoretically possible to hydrolyze cyanophycin using mild chemicals or enzymes to produce copolymers of poly-aspartic acid (PASP) with lower arginine concentration or even PASP homopolymer, a water-soluble and biodegradable polymer. PASP can be used as a dispersant for a range of organic and inorganic substances or as an antiscalant component in dishwashing and laundry detergents [112]. Furthermore, according to Mooibroek et al. [113], cyanophycin has the ability to be converted into a number of bulk compounds that contain nitrogen, including urea, 1,4-butanediol, and acrylonitrile. When cyanophycin is manufactured in plants, no extra facilities or specialized machinery are needed. Therefore, the product’s cost is very comparable to that of other polyamino acids, expanding the range of widely recognized uses for microbial cyanophycin [114].
Cyanophycin is also known as cyanophycin granule polypeptide. Many cyanobacteria synthesize cyanophycin as a short-term nitrogen reserve material. The cyanophycin synthetase enzyme catalyzes the polymerization reaction. Because purified cyanophycin may be chemically changed into a polymer with a lower arginine content, like poly-aspartic acid, it may be utilized as a biodegradable alternative to synthetic polyacrylates. This makes cyanophycin interesting for usage in biomedicine. However, bacterial production of cyanophycin on a large scale is severely impeded by the low yield and the highly complicated fermentation process, and the search is underway for new methods for large-scale production [110,113,257,266,267,268].
6. Protein Hydrogels
Proteins have four levels of structure: primary, secondary, tertiary, and quaternary. The primary structure is the sequence of amino acids in a polypeptide chain. The secondary structure is the α helix and/or the β sheet along with turn and loop components. The tertiary structure, also called 3D structure, is primarily due to interactions between the R groups of the amino acids that make up the polypeptide chain. Many proteins are composed of a single polypeptide chain and only have three levels of structure, while some proteins have multiple polypeptide chains (subunits) to make their quaternary structure. The sequence of amino acids in a protein is the primary factor that determines the bioactivity and nature of proteins and the four levels of structures [269]. The transition from the tertiary structure to the secondary structure is accompanied by conformational changes that result in an increased random coil structure. The increased random coil structure improves the flexibility of protein structure, making it more suitable for gelation [270,271].
Protein gelation is a phenomenon in which proteins undergo denaturation, resulting in their subsequent deposition inside a gel matrix, which is widely recognized and commonly applied for making protein hydrogels. Protein hydrogels can be generated through several methods, including physical, chemical, and enzymatic cross-linking procedures [272], which are facilitated by the intrinsic carboxyl and amino functional groups present in proteins. It has been shown that proteins containing cysteine residues demonstrate an enhanced capacity in the formation of hydrogels. The molecular structure of cysteine has a sulfhydryl (-SH) group, which accounts for its capacity to augment water retention and absorption capabilities. The swelling ratio of a material can be affected by several factors, including concentration of proteins, pH levels, and the presence of hydrophilic residues. Manipulation these parameters can be used to achieve the desired swelling characteristics in certain applications [273]. Yan et al. [274] altered the soy protein isolate and sugar beet pectin concentrations to change the swelling ratio of the hydrogel’s interpenetrating polymeric network. Joseph et al. [275] investigated the effects on the swelling ratio and adhesion characteristics through incorporating fibrin microparticles into hydrogels made of polyethylene glycol (PEG) and fibrinogen. Yan et al. [276] revealed that the hydrogel exhibited a lower degree of swelling in stomach fluid compared to intestinal fluid, which can be attributed to variations in pH levels.
Protein hydrogels are ideal materials for diverse medical applications, as they are biocompatible, biodegradable, accessible, and renewable. They can be redesigned and optimized through protein engineering. Protein-based hydrogels demonstrated favorable attributes, such as structural integrity, stability, strength, and a variety of unique properties like responsiveness to external stimuli and self-repair capabilities [277,278,279,280].
Polymeric hydrogels possess the capability to form a gel matrix through both covalent and non-covalent cross-linking methodologies. The incorporation of physical cross-linking methods in the production of protein-based hydrogels offers two significant advantages. Firstly, these procedures provide a simple and environmentally friendly operation, therefore distinguishing them from alternatives. Furthermore, they contribute to the retention of many of the protein’s intrinsic properties. Physical cross-linking plays a crucial role in augmenting the intermolecular contacts of macromolecular chains, hence promoting the production of polymeric biomaterial hydrogels [281]. Physically cross-linked hydrogels are formed through the synergistic effects of many weak intermolecular forces, including stacking, electrostatic interactions, hydrogen bonding, and physical entanglement between molecules [282]. A noteworthy advantage of physically cross-linked polybutylene hydrogels is their ability to undergo injection and self-healing at a temperature of 25 °C. Previous studies have demonstrated the efficacy of employing physical cross-linking techniques in maintaining the biocompatibility of proteins, thereby augmenting their potential utility within the realm of tissue engineering [282].
In the field of physical cross-linking, it is essential to initiate the denaturation of proteins, followed by their subsequent reassembly into their secondary structure. This process is crucial for the development of a gel matrix that exhibits a higher proportion of β-sheet composition [283]. Hu et al. employed ultrasound as a method to induce conformational unfolding for silk proteins, leading to an augmentation in the β-sheet content [284]. They later utilized hyaluronic acid to achieve physical cross-linking. Yan sought to achieve a similar objective by integrating hyaluronic acid into a silk protein solution. The researchers used a solvent exchange technique to produce a conformational arrangement typified by beta-sheets in the protein molecules. Consequently, the researchers achieved successful production of a hydrogel that demonstrated characteristics comparable to the silk protein [285]. Moreover, the concentration of proteins plays a significant role in the occurrence of physical cross-linking in this specific setting. It has been empirically shown that the protein demonstrates self-crosslinking as the concentration of the protein rises [217,286].
Polymeric biohybrid hydrogels can be synthesized by employing hydrogels with either a single network wrapping or multiple networks superposition. The mechanical characteristics of hydrogels composed of a single matrix are often limited. Tang et al. [287] utilized a protein misfolding methodology to produce a protein matrix. The protein was subjected to thermal exposure, leading to denaturation, aggregation, and subsequent gelation. Xu et al. [288] utilized a freeze–thawing method to generate a composite solution consisting of polyvinyl alcohol (PVA) and bovine serum albumin (BSA). In summary, the researchers synthesized the main hydrogel matrix as mentioned earlier. Following that, a secondary hydrogel matrix was produced through the process of physically cross-linking tannic acid with PVA and BSA, using hydrophobic interactions and hydrogen bonding. The study showed that the incorporation of both developed networks led to improved mechanical characteristics of the protein-based double-matrix hydrogel. The enhancement of feeble connections among lengthy macromolecular chains of polymers and proteins is encouraged by the elongated structure and the existence of electrostatic interactions or hydrogen bonds. The process of hydrogel production does not necessitate the complete denaturation of proteins. Without a doubt, the phenomenon of denaturation might be considered negative. The facilitation of the process can be effectively achieved through the structural transitions from tertiary to secondary conformation, or through the enhancement of β-sheet content. The occurrence of hydrogen bond production is responsible for a range of material characteristics, including thermoplasticity, self-healing ability, recyclability, customizable remodeling, and reusability. The presence of hydrogen bonds between residues and polymers can potentially aid the formation of dynamic bonds, leading to an increased likelihood of bond formation. The formation of hydrogen bonding by polar amino acids has been reported [289,290].
Numerous chemical cross-linking methodologies have been documented in scholarly publications, encompassing covalent cross-linking of protein chains and target residues, chemical coupling, and click reactions [291,292,293,294,295,296]. Prior research has indicated that the application of the cross-linking technique can enhance stability, regulate degradation rate, and exhibit notable mechanical properties in physiological conditions [297]. Moreover, the structural attributes of synthesized hydrogels can be modulated by the source of covalent linkages. The inclusion of polymers enhances the establishment of covalent linkages between proteins, leading to the development of hydrogels with unique properties. Hu et al. [284] examined the mechanism underlying the creation of covalent bonds. The chemical cross-linking of graphene, which had been modified with resilin-like proteins, was effectively achieved by the researchers. The technique of cross-linking resulted in significant improvements in several aspects, such as adhesion, elongation, and sensitivity. Wang et al. utilized a comparable modification methodology to facilitate the formation of C=C bonds by the integration of the vinyl group into gelatin. The aim was to produce methacrylate gelatin nanoparticles (MA-GNP) as the principal component for the crosslinking procedure [298]. Subsequently, acrylamide was added to MA-GNP to fabricate macromolecular microsphere composite hydrogels [6,298].
In a recent study by Su, chitosan was utilized as a vehicle for performing a chemical cross-linking technique, resulting in the formation of covalent imide linkages. Proteins with charge characteristics were utilized to fabricate hydrogels that have the ability to encapsulate diverse particles [299]. Huang et al. [217] conducted a significant study where they used a novel methodology to create hydrogels with rubber-like characteristics and specific mechanical properties. This involved integrating expanded silk networks into resilin blocks and employing photochemical cross-linking techniques. Through the manipulation of the silk to resilin ratio, the copolymers exhibited the capability to undergo self-organization, resulting in the formation of fibrous structures within a defined time frame. As a result, this event enabled the intentional formation of fibrils within copolymer solutions using supramolecular methods. The solutions were obtained using the light cross-linking approach implemented on supported hydrogels. Moreover, this served as a demonstration of the impact of the protein-to-polymer ratio on the mechanism of hydrogel formation. Furthermore, the integration of proteins into hydrogels can be utilized to alter and augment their characteristics. Wang et al. (2022) [300] have effectively developed a polymeric biomaterial hydrogel (PBH) that possesses noteworthy attributes such as anti-freezing capabilities, biocompatibility, and tunability. The achievement involved a two-step modification process. The first focus of the study involved the incorporation of anti-freezing proteins obtained from native fish species. Additionally, a chemical cross-linking technique was used to construct the desired polymeric hydrogel system [300].
The properties of hydrogels can be enhanced by integrating polymers with cross-linking or modifying qualities into the composition of polybutylene succinate-co-adipate structures. The polymers under consideration establish chemical interactions with proteins. Polymers have the capacity to serve as cross-linking agents enabling the modification of the amine and carboxyl termini of proteins. The aforementioned procedure results in the development of hydrogels that demonstrate significant resistance to fatigue, as supported by prior investigations [301,302]. Enzymes, serving as biocatalysts, offer notable benefits in facilitating and directing the hydrogel synthesis process. The characteristics indicated above encompass the performance of cross-linking reactions in mild reaction circumstances, demonstrating selectivity in terms of chemical, spatial, and stereochemical preferences, and decreasing the dependence on hazardous cross-linking agents [303]. In their latest study, Le Thi et al. [304] used enzymatic cross-linking as a supplementary strategy for cross-linking. The researchers developed a dual-enzymatic cross-linking approach using tyrosinase and horseradish peroxidase (HRP) in order to create a gelatin-based adhesive PBH [304]. The reactivity of gelatin towards nucleophilic molecules, such as amines or thiols, resulted in the creation of strong tissue adhesion. The reaction being examined involves the conversion of the phenol groups found in gelatin into o-quinone, assisted by the catalytic activity of tyrosinase. The utilization of a dual-enzymatic cross-linking approach has been shown to result in the production of hydrogels with greatly increased adhesive strength. Chirilaet et al. [305] conducted a study wherein silk fibroins were treated to self-crosslinking using the HRP enzyme and H2O2. The results of the study provided evidence supporting the benefits linked to the use of HRP in the process of cross-linking. The benefits that were noticed encompassed a noteworthy decrease in the gelation time, heightened flexibility, and improved compatibility with live cells. Hou et al. [306] performed an evaluation with the objective of developing a biofunctional PBH that is characterized by cost-effectiveness and simplicity. The researchers achieved this by exploiting gelatin and applying enzymatic cross-linking assisted by microbial transglutaminase (mTG) [306]. In the context of this framework, the application of mTG has led to the development of adhesive properties in gelatin microgels and the formation of a large hydrogel with pore sizes that promote cellular migration and proliferation.
7. Fabrication of Protein Fabrics
Multiple fiber-forming production techniques are routinely employed in many industries, such as melt spinning, dry spinning, wet spinning, and electrospinning [307,308,309,310,311,312,313,314]. These techniques utilize high-pressure solution extrusion to produce fibers, which are then subjected to other post-processing operations. The solidification of fibers in melt and dry spinning methods requires the application of cooling gases, but in wet spinning, precipitation is used to achieve the same objective [315]. After the fibers are formed, they undergo a sequence of procedures that involve collection, stretching, and alignment. These activities are facilitated using spindles and reels.
The electrospinning process is characterized by its deviation from conventional extrusion spinning techniques, as it relies on the application of an electric field within a voltage range of 5–20 kV, rather than solely depending on tensile force, to commence the formation of a polymer jet [316]. Electrospun fibers are produced with the use of an electric potential difference to a charged initial solution, with a grounded collector being employed. The solidification of the fibers is a result of the phenomenon of evaporation. The application of electrospinning in polymer fiber manufacturing is characterized by its versatility. However, it is important to acknowledge the existence of some constraints that restrict its effectiveness. These limits include a comparatively low production rate of 0.5 g per hour per spinneret as well as challenges in maintaining precise control over fiber diameter and orientation [315]. Previous studies have reported the impact of solvent conductivity on the trajectory of the polymer jet towards the collector substrate [317,318].
8. Perspectives
Most petroleum-based plastics are non-biodegradable, resulting in continuous accumulation in both terrestrial ecosystems and aquatic ecosystems. Plastic pollution causes severe environmental damage and threatens human health [1,5]. It is more pressing than ever to accelerate the development of biodegradable plastics to replace petroleum-based, non-degradable plastics, thereby reducing plastic pollution [319,320].
There are two major types of proteins in nature: fibrous proteins and globular proteins. Fibrous proteins offer a compelling prospect for making protein-based plastics to replace petroleum-based plastics. Fibrous proteins include keratin (hair, nails, feathers, horns), collagen (connective tissue), fibroin (silk), myosin (muscle), and novel filament-forming proteins (Fm7001 and All4981) recently identified in cyanobacteria [321]. The N2-fixing cyanobacteria are capable of photosynthetically producing high amounts of protein using only air (N2, CO2), mineralized H2O and sunlight. Cyanobacterial protein contents range from 46–71% in dry biomass [322,323], which is higher than conventional food protein sources, such as meat (43%), milk (26%), and soybean (37%) [322,323,324]. Although plant systems are now gaining acceptance as a platform for production of recombinant proteins, there is still resistance to commercial uptake. This is partly due to the relatively low yields achieved in plants, as well as being time-consuming and posing difficulties with downstream processing. Surprisingly, humans almost ignored the use of N2-fixing cyanobacteria, the “protein-making machinery” [322,323], having both the advantages of plant systems and prokaryotic systems, plus being capable of using N2 gas as sole nitrogen source to produce high yield of foreign proteins. Dinitrogen (N2 gas) resource is nearly 2000-fold more abundant (78%) than CO2 resources (0.04%) in the atmosphere and can be directly converted into amino acids by N2-fixing cyanobacteria, and fibrous proteins for making bioplastics can subsequently be synthesized through a synthetic biology approach.
Recently we also discovered in the genomic database of cyanobacteria that N2-fixing cyanobacteria may natively produce more than 11 super large proteins, the sizes ranging from 3883aa to 11342aa (Zhou R et al. unpublished data), much larger than the spider dragline silk proteins (3779aa). There has been no report of the use of N2-fixing cyanobacteria, a potentially commercially viable system, to produce full length spider silk proteins and any protein-based polymers mentioned above. Recently, research on photosynthetic production of recombinant proteins by cyanobacteria is underway [321,325,326,327]. Therefore, the N2-fixing cyanobacteria are the most attractive organisms chosen to overproduce N-rich polymers, the polymerized proteins for bioplastics. Using synthetic biology combined with machine-learning-guided protein engineering [328,329], we are very confident that we can genetically engineer N2-fixing cyanobacteria to produce all protein-based polymers mentioned in this review.
9. Conclusions
Synthetic biology combined with machine learning-guided protein engineering [328] has the great potential to create solutions for complex problems, subsequently revolutionizing our way of living, working, and understanding of nature. Synthetic biology designs and constructs new biological parts, devices, and systems that do not exist in nature. It is focused on engineering biology to produce commodities to benefit society, while biomimicry is about innovation inspired by nature to mimic nature’s genius to solve human problems. This review explored the promising future for protein-based biomaterials. The review targets protein-based polymers, focusing on their classification, application, structural arrangements, and integration into advanced patterning techniques. The classification of proteins based on their structural characteristics offers valuable insights into the diverse array of protein-based biomaterials, each possessing unique properties and potential applications.
The investigation and use of protein-based materials presents a promising domain within the realm of biomaterial research. This entails the innate natural design to generate innovative and sophisticated solutions. The qualities and activities of native proteins are largely determined by their various structural arrangements, which also provide the basis for improvement. Moreover, structural modifications present an intriguing prospect for creating protein-based novel materials with precision and control.
Generating patterns on photoresists by chemically modified proteins showed a significant potential of biomaterials in contemporary manufacturing methodologies. The use of a composite film derived from soy protein serves as an illustration of the wide array of protein sources that can be employed for the advancement of functional materials. The development of biodegradable wheat-glutenin-based polymers serving as environmentally friendly alternatives to petroleum-based plastics; the variety of applications for sponge in extreme biomimetics, bioinspired materials; and the promising future of conchiolin proteins in cosmetics and wound healing are just a few examples of the innovative and sustainable advancements being made in the field of protein-based materials.
A wide range of potential applications with protein-based materials is emerging from research into the interface of protein science, materials engineering, and advanced technologies. The entities possess unique structural configurations that, in conjunction with their ability to facilitate intentional modifications, play a central role in the advancement of patterned biomaterials. The application of a diverse array of proteins and their intricate structural properties enables researchers and engineers to propel the improvement of novel biomaterials, with potential implications across various domains including biotechnology and nanotechnology, among others. This review underscores the considerable potential of protein-based materials as a compelling avenue for scientific inquiry and technological advancement.
Author Contributions
Conceptualization, T.N., L.G. and Z.H.; writing—original draft preparation, T.N. and L.G.; writing—review and editing, Z.H., J.G. and R.Z.; supervision, R.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research is partially funded by USDA-NIFA grant award number: 2023-67022-39594-Genetic engineering of cyanobacteria to produce high-value proteins using atmospheric N2 gas (to L.G.).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable. This is a review article that does not generate substantially new data.
Acknowledgments
The authors thank the reviewers very much for their constructive comments for significant improvement of this review article. The authors appreciate the USDA Supported Hatch Project SD00H691-20 titled Engineering Solar-powered N2-fixing Cyanobacteria for Agricultural and Industrial Applications (to R.Z.).
Conflicts of Interest
Author Jaimie Gibbons was employed by the company Houdek. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Lau, W.W.; Shiran, Y.; Bailey, R.M.; Cook, E.; Stuchtey, M.R.; Koskella, J.; Velis, C.A.; Godfrey, L.; Boucher, J.; Murphy, M.B. Evaluating scenarios toward zero plastic pollution. Science 2020, 369, 1455–1461. [Google Scholar] [CrossRef]
- He, Y.X.; Zhang, N.N.; Li, W.F.; Jia, N.; Chen, B.Y.; Zhou, K.; Zhang, J.; Chen, Y.; Zhou, C.Z. N-Terminal domain of Bombyx mori fibroin mediates the assembly of silk in response to pH decrease. J. Mol. Biol. 2012, 418, 197–207. [Google Scholar] [CrossRef]
- Kane, I.A.; Clare, M.A.; Miramontes, E.; Wogelius, R.; Rothwell, J.J.; Garreau, P.; Pohl, F. Seafloor microplastic hotspots controlled by deep-sea circulation. Science 2020, 368, 1140–1145. [Google Scholar] [CrossRef]
- Jambeck, J.R.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A.; Narayan, R.; Law, K.L. Plastic waste inputs from land into the ocean. Science 2015, 347, 768–771. [Google Scholar] [CrossRef]
- Borrelle, S.B.; Ringma, J.; Law, K.L.; Monnahan, C.C.; Lebreton, L.; McGivern, A.; Murphy, E.; Jambeck, J.; Leonard, G.H.; Hilleary, M.A. Predicted growth in plastic waste exceeds efforts to mitigate plastic pollution. Science 2020, 369, 1515–1518. [Google Scholar] [CrossRef]
- Kadumudi, F.B.; Hasany, M.; Pierchala, M.K.; Jahanshahi, M.; Taebnia, N.; Mehrali, M.; Mitu, C.F.; Shahbazi, M.A.; Zsurzsan, T.G.; Knott, A. The Manufacture of Unbreakable Bionics via Multifunctional and Self-Healing Silk–Graphene Hydrogels. Adv. Mater. 2021, 33, 2170276. [Google Scholar] [CrossRef]
- Gorrasi, G.; Sorrentino, A.; Lichtfouse, E. Back to plastic pollution in COVID times. Environ. Chem. Lett. 2021, 19, 1–4. [Google Scholar] [CrossRef]
- Haider, T.P.; Völker, C.; Kramm, J.; Landfester, K.; Wurm, F.R. Plastics of the future? The impact of biodegradable polymers on the environment and on society. Angew. Chem. Int. Ed. 2019, 58, 50–62. [Google Scholar] [CrossRef]
- Chiellini, E.; Solaro, R. Biodegradable polymeric materials. Adv. Mater. 1996, 8, 305–313. [Google Scholar] [CrossRef]
- Li, K.; Jin, S.; Jiang, S.; Li, X.; Li, J.; Shi, S.Q.; Li, J. Bioinspired mineral–organic strategy for fabricating a high-strength, antibacterial, flame-retardant soy protein bioplastic via internal boron–nitrogen coordination. Chem. Eng. J. 2022, 428, 132616. [Google Scholar] [CrossRef]
- Chen, L.; Qiang, T.; Chen, X.; Ren, W.; Zhang, H.J. Fabrication and evaluation of biodegradable multi-cross-linked mulch film based on waste gelatin. Chem. Eng. J. 2021, 419, 129639. [Google Scholar] [CrossRef]
- Pang, X.; Zhuang, X.; Tang, Z.; Chen, X. Polylactic acid (PLA): Research, development and industrialization. Biotechnol. J. 2010, 5, 1125–1136. [Google Scholar] [CrossRef]
- Ghorpade, V.M.; Gennadios, A.; Hanna, M.A. Laboratory composting of extruded poly (lactic acid) sheets. Bioresour. Technol. 2001, 76, 57–61. [Google Scholar] [CrossRef]
- Sun, C.; Wei, S.; Tan, H.; Huang, Y.; Zhang, Y. Progress in upcycling polylactic acid waste as an alternative carbon source: A review. Chem. Eng. J. 2022, 446, 136881. [Google Scholar] [CrossRef]
- Li, F.; Liu, T.; Gu, W.; Gao, Q.; Li, J.; Shi, S.Q. Bioinspired super-tough and multifunctional soy protein-based material via a facile approach. Chem. Eng. J. 2021, 405, 126700. [Google Scholar] [CrossRef]
- Liu, Y.; Shao, Z.; Vollrath, F. Elasticity of spider silks. Biomacromolecules 2008, 9, 1782–1786. [Google Scholar] [CrossRef]
- Rising, A.; Widhe, M.; Johansson, J.; Hedhammar, M. Spider silk proteins: Recent advances in recombinant production, structure-function relationships and biomedical applications. Cell Mol. Life Sci. 2011, 68, 169–184. [Google Scholar] [CrossRef]
- Foo, C.W.P.; Bini, E.; Huang, J.; Lee, S.Y.; Kaplan, D.l. Solution behavior of synthetic silk peptides and modified recombinant silk proteins. Appl. Phys. A 2006, 82, 193–203. [Google Scholar] [CrossRef]
- Rising, A.; Johansson, J. Toward spinning artificial spider silk. Nat. Chem. Biol. 2015, 11, 309–315. [Google Scholar] [CrossRef]
- Sun, J.; Su, J.; Ma, C.; Göstl, R.; Herrmann, A.; Liu, K.; Zhang, H. Fabrication and mechanical properties of engineered protein-based adhesives and fibers. Adv. Mater. 2020, 32, 1906360. [Google Scholar] [CrossRef]
- Xiao, T.; Wang, Y.; Wei, H.; Yu, P.; Jiang, Y.; Mao, L. Electrochemical monitoring of propagative fluctuation of ascorbate in the live rat brain during spreading depolarization. Angew. Chem. Int. Ed. 2019, 58, 6616–6619. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Yu, Q.; Wen, J.; Li, C.; Guo, Z.; Wang, X.; Wang, N. Ultrafast and highly selective uranium extraction from seawater by hydrogel-like spidroin-based protein fiber. Angew. Chem. 2019, 131, 11911–11916. [Google Scholar] [CrossRef]
- Lewis, R.V. Spider silk: Ancient ideas for new biomaterials. Chem. Rev. 2006, 106, 3762–3774. [Google Scholar] [CrossRef] [PubMed]
- Kluge, J.A.; Rabotyagova, O.; Leisk, G.G.; Kaplan, D.L. Spider silks and their applications. Trends Biotechnol. 2008, 26, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.G.; Rőmer, L.M.; Scheibel, T.R. Polymeric materials based on silk proteins. Polymer 2008, 49, 4309–4327. [Google Scholar] [CrossRef]
- Wang, X.; Pang, Z.; Chen, C.; Xia, Q.; Zhou, Y.; Jing, S.; Wang, R.; Ray, U.; Gan, W.; Li, C. All-natural, degradable, rolled-up straws based on cellulose micro-and nano-hybrid fibers. Adv. Funct. Mater. 2020, 30, 1910417. [Google Scholar] [CrossRef]
- Song, P.; Wang, H. High-performance polymeric materials through hydrogen-bond cross-linking. Adv. Mater. 2020, 32, 1901244. [Google Scholar] [CrossRef] [PubMed]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef] [PubMed]
- Myllyharju, J.; Kivirikko, K.I. Collagens, modifying enzymes and their mutations in humans, flies and worms. Trends Genet. 2004, 20, 33–43. [Google Scholar] [CrossRef]
- Gordon, M.K.; Hahn, R.A. Collagens. Cell Tissue Res. 2010, 339, 247–257. [Google Scholar] [CrossRef]
- Wess, T.J. Collagen fibril form and function. Adv. Protein Chem. 2005, 70, 341–374. [Google Scholar] [PubMed]
- Kadler, K.E.; Baldock, C.; Bella, J.; Boot-Handford, R.P. Collagens at a glance. J. Cell Sci. 2007, 120, 1955–1958. [Google Scholar] [CrossRef]
- Heim, M.; Keerl, D.; Scheibel, T. Spinnenseide: Vom löslichen Protein zur außergewöhnlichen Faser. Angew. Chem. 2009, 121, 3638–3650. [Google Scholar] [CrossRef]
- Omenetto, F.G.; Kaplan, D.L. New opportunities for an ancient material. Science 2010, 329, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Hearle, J.W. Protein fibers: Structural mechanics and future opportunities. J. Mater. Sci. 2007, 42, 8010–8019. [Google Scholar] [CrossRef]
- Scheibel, T. Protein fibers as performance proteins: New technologies and applications. Curr. Opin. Biotechnol. 2005, 16, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Sponner, A.; Unger, E.; Grosse, F.; Weisshart, K. Differential polymerization of the two main protein components of dragline silk during fibre spinning. Nat. Mater. 2005, 4, 772–775. [Google Scholar] [CrossRef] [PubMed]
- Motta, A.; Migliaresi, C.; Faccioni, F.; Torricelli, P.; Fini, M.; Giardino, R. Fibroin hydrogels for biomedical applications: Preparation, characterization and in vitro cell culture studies. J. Biomater. Sci. Polym. Ed. 2004, 15, 851–864. [Google Scholar] [CrossRef]
- Sah, M.; Pramanik, K. Preparation, characterization and in vitro study of biocompatible fibroin hydrogel. Afr. J. Biotechnol. 2011, 10, 7878–7892. [Google Scholar]
- Sabatier, L.; Miosge, N.; Hubmacher, D.; Lin, G.; Davis, E.C.; Reinhardt, D.P. Fibrillin-3 expression in human development. Matrix Biol. 2011, 30, 43–52. [Google Scholar] [CrossRef]
- Corson, G.M.; Charbonneau, N.L.; Keene, D.R.; Sakai, L.Y. Differential expression of fibrillin-3 adds to microfibril variety in human and avian, but not rodent, connective tissues. Genomics 2004, 83, 461–472. [Google Scholar] [CrossRef]
- Zhang, H.; Apfelroth, S.D.; Hu, W.; Davis, E.C.; Sanguineti, C.; Bonadio, J.; Mecham, R.P.; Ramirez, F. Structure and expression of fibrillin-2, a novel microfibrillar component preferentially located in elastic matrices. J. Cell Biol. 1994, 124, 855–863. [Google Scholar] [CrossRef]
- Hubmacher, D.; Tiedemann, K.; Reinhardt, D.P. Fibrillins: From biogenesis of microfibrils to signaling functions. Curr. Top. Dev. Biol. 2006, 75, 93–123. [Google Scholar]
- Baldock, C.; Koster, A.J.; Ziese, U.; Rock, M.J.; Sherratt, M.J.; Kadler, K.E.; Shuttleworth, C.A.; Kielty, C.M. The supramolecular organization of fibrillin-rich microfibrils. J. Cell Biol. 2001, 152, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Kielty, C.M.; Sherratt, M.J.; Shuttleworth, C.A. Elastic fibres. J. Cell Sci. 2002, 115, 2817–2828. [Google Scholar] [CrossRef]
- Visconti, R.P.; Barth, J.L.; Keeley, F.W.; Little, C.D. Codistribution analysis of elastin and related fibrillar proteins in early vertebrate development. Matrix Biol. 2003, 22, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Rosenbloom, J.; Abrams, W.R.; Mecham, R. Extracellular matrix 4: The elastic fiber. FASEB J. 1993, 7, 1208–1218. [Google Scholar] [CrossRef]
- Katoh, K.; Shibayama, M.; Tanabe, T.; Yamauchi, K. Preparation and physicochemical properties of compression-molded keratin films. Biomaterials 2004, 25, 2265–2272. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.M.; Rippon, J.A. The Coloration of Wool and Other Keratin Fibres; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Negri, A.P.; Cornell, H.J.; Rivett, D.E. A model for the surface of keratin fibers. Text. Res. J. 1993, 63, 109–115. [Google Scholar] [CrossRef]
- Chen, Q.; Gao, K.; Peng, C.; Xie, H.; Zhao, Z.K.; Bao, M. Preparation of lignin/glycerol-based bis (cyclic carbonate) for the synthesis of polyurethanes. Green. Chem. 2015, 17, 4546–4551. [Google Scholar] [CrossRef]
- Lee, C.H.; Kim, M.S.; Chung, B.M.; Leahy, D.J.; Coulombe, P.A. Structural basis for heteromeric assembly and perinuclear organization of keratin filaments. Nat. Struct. Mol. Biol. 2012, 19, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Gosline, J.; Lillie, M.; Carrington, E.; Guerette, P.; Ortlepp, C.; Savage, K. Elastic proteins: Biological roles and mechanical properties. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2002, 357, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Sage, H. Structure-function relationships in the evolution of elastin. J. Investig. Dermatol. 1982, 79, 146–153. [Google Scholar] [CrossRef][Green Version]
- Faury, G. Function–structure relationship of elastic arteries in evolution: From microfibrils to elastin and elastic fibres. Pathol. Biol. 2001, 49, 310–325. [Google Scholar] [CrossRef]
- Sage, H.; Gray, W. Studies on the evolution of elastin--I. Phylogenetic distribution. Comp. Biochem. Physiol. B Comp. Biochem. 1979, 64, 313–327. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.C. Stability of elastin in the developing mouse aorta: A quantitative radioautographic study. Histochemistry 1993, 100, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, S.; Endicott, S.; Province, M.; Pierce, J.; Campbell, E. Marked longevity of human lung parenchymal elastic fibers deduced from prevalence of D-aspartate and nuclear weapons-related radiocarbon. J. Clin. Investig. 1991, 87, 1828–1834. [Google Scholar] [CrossRef]
- Keeley, F.W.; Bellingham, C.M.; Woodhouse, K.A. Elastin as a self–organizing biomaterial: Use of recombinantly expressed human elastin polypeptides as a model for investigations of structure and self–assembly of elastin. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2002, 357, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Pepe, A.; Guerra, D.; Bochicchio, B.; Quaglino, D.; Gheduzzi, D.; Ronchetti, I.P.; Tamburro, A.M. Dissection of human tropoelastin: Supramolecular organization of polypeptide sequences coded by particular exons. Matrix Biol. 2005, 24, 96–109. [Google Scholar] [CrossRef]
- Tamburro, A.M.; Bochicchio, B.; Pepe, A. Dissection of human tropoelastin: Exon-by-exon chemical synthesis and related conformational studies. Biochemistry 2003, 42, 13347–13362. [Google Scholar] [CrossRef]
- Muiznieks, L.D.; Keeley, F.W. Proline periodicity modulates the self-assembly properties of elastin-like polypeptides. J. Biol. Chem. 2010, 285, 39779–39789. [Google Scholar] [CrossRef] [PubMed]
- Rauscher, S.; Baud, S.; Miao, M.; Keeley, F.W.; Pomes, R. Proline and glycine control protein self-organization into elastomeric or amyloid fibrils. Structure 2006, 14, 1667–1676. [Google Scholar] [CrossRef] [PubMed]
- Weis-Fogh, T. Molecular interpretation of the elasticity of resilin, a rubber-like protein. J. Mol. Biol. 1961, 3, 648–667. [Google Scholar] [CrossRef]
- Weis-Fogh, T. Thermodynamic properties of resilin, a rubber-like protein. J. Mol. Biol. 1961, 3, 520–531. [Google Scholar] [CrossRef]
- Truong, M.Y.; Dutta, N.K.; Choudhury, N.R.; Kim, M.; Elvin, C.M.; Nairn, K.M.; Hill, A.J. The effect of hydration on molecular chain mobility and the viscoelastic behavior of resilin-mimetic protein-based hydrogels. Biomaterials 2011, 32, 8462–8473. [Google Scholar] [CrossRef] [PubMed]
- Renner, J.N.; Cherry, K.M.; Su, R.S.-C.; Liu, J.C. Characterization of resilin-based materials for tissue engineering applications. Biomacromolecules 2012, 13, 3678–3685. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Tong, Z.; Jia, X.; Kiick, K.L. Resilin-like polypeptide hydrogels engineered for versatile biological function. Soft Matter 2013, 9, 665–673. [Google Scholar] [CrossRef] [PubMed]
- McGann, C.L.; Levenson, E.A.; Kiick, K.L. Resilin-based hybrid hydrogels for cardiovascular tissue engineering. Macromol. Chem. Phys. 2013, 214, 203–213. [Google Scholar] [CrossRef]
- Johansson, E.; Malik, A.H.; Hussain, A.; Rasheed, F.; Newson, W.R.; Plivelic, T.; Hedenqvist, M.S.; Gällstedt, M.; Kuktaite, R. Wheat gluten polymer structures: The impact of genotype, environment, and processing on their functionality in various applications. Cereal Chem. 2013, 90, 367–376. [Google Scholar] [CrossRef]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24, 115–119. [Google Scholar] [CrossRef]
- Wrigley, C.W. Giant proteins with flour power. Nature 1996, 381, 738–739. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Rabu, J.; Goulin, K.; Sainlaud, C.; Chen, F.; Johansson, E.; Olsson, R.T.; Hedenqvist, M.S. Flexible strength-improved and crack-resistant biocomposites based on plasticised wheat gluten reinforced with a flax-fibre-weave. Compos. Part. A Appl. Sci. Manuf. 2017, 94, 61–69. [Google Scholar] [CrossRef]
- Richter, R.; Hejazi, M.; Kraft, R.; Ziegler, K.; Lockau, W. Cyanophycinase, a peptidase degrading the cyanobacterial reserve material multi-L-arginyl-poly-L-aspartic acid (cyanophycin) Molecular cloning of the gene of Synechocystis sp. PCC 6803, expression in Escherichia coli, and biochemical characterization of the purified enzyme. Eur. J. Biochem. 1999, 263, 163–169. [Google Scholar] [PubMed]
- Brahney, J.; Hallerud, M.; Heim, E.; Hahnenberger, M.; Sukumaran, S. Plastic rain in protected areas of the United States. Science 2020, 368, 1257–1260. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H.; Wysokowski, M.; Żółtowska-Aksamitowska, S.; Petrenko, I.; Jesionowski, T. Collagens of poriferan origin. Mar. Drugs 2018, 16, 79. [Google Scholar] [CrossRef] [PubMed]
- Szatkowski, T.; Wysokowski, M.; Lota, G.; Pęziak, D.; Bazhenov, V.V.; Nowaczyk, G.; Walter, J.; Molodtsov, S.L.; Stöcker, H.; Himcinschi, C. Novel nanostructured hematite–spongin composite developed using an extreme biomimetic approach. RSC Adv. 2015, 5, 79031–79040. [Google Scholar] [CrossRef]
- Szatkowski, T.; Kopczyński, K.; Motylenko, M.; Borrmann, H.; Mania, B.; Graś, M.; Lota, G.; Bazhenov, V.V.; Rafaja, D.; Roth, F. Extreme biomimetics: A carbonized 3D spongin scaffold as a novel support for nanostructured manganese oxide (IV) and its electrochemical applications. Nano Res. 2018, 11, 4199–4214. [Google Scholar] [CrossRef]
- Zdarta, J.; Norman, M.; Smułek, W.; Moszyński, D.; Kaczorek, E.; Stelling, A.L.; Ehrlich, H.; Jesionowski, T. Spongin-based scaffolds from Hippospongia communis demosponge as an effective support for lipase immobilization. Catalysts 2017, 7, 147. [Google Scholar] [CrossRef]
- Zdarta, J.; Antecka, K.; Frankowski, R.; Zgoła-Grześkowiak, A.; Ehrlich, H.; Jesionowski, T. The effect of operational parameters on the biodegradation of bisphenols by Trametes versicolor laccase immobilized on Hippospongia communis spongin scaffolds. Sci. Total Environ. 2018, 615, 784–795. [Google Scholar] [CrossRef]
- Norman, M.; Żółtowska-Aksamitowska, S.; Zgoła-Grześkowiak, A.; Ehrlich, H.; Jesionowski, T. Iron (III) phthalocyanine supported on a spongin scaffold as an advanced photocatalyst in a highly efficient removal process of halophenols and bisphenol A. J. Hazard. Mater. 2018, 347, 78–88. [Google Scholar] [CrossRef]
- Ashouri, V.; Adib, K.; Rahimi-Nasrabadi, M. Pre-concentration and extraction of fenitrothion using a prefabricated 3D spongin-based skeleton of marine demosponge: Optimization by experimental design. Appl. Phys. A 2020, 126, 860. [Google Scholar] [CrossRef]
- Zarrintaj, P.; Manouchehri, S.; Ahmadi, Z.; Saeb, M.R.; Urbanska, A.M.; Kaplan, D.L.; Mozafari, M. Agarose-based biomaterials for tissue engineering. Carbohydr. Polym. 2018, 187, 66–84. [Google Scholar] [CrossRef] [PubMed]
- Campos, F.; Bonhome-Espinosa, A.B.; Chato-Astrain, J.; Sánchez-Porras, D.; García-García, Ó.D.; Carmona, R.; López-López, M.T.; Alaminos, M.; Carriel, V.; Rodriguez, I.A. Evaluation of fibrin-agarose tissue-like hydrogels biocompatibility for tissue engineering applications. Front. Bioeng. Biotechnol. 2020, 8, 596. [Google Scholar] [CrossRef] [PubMed]
- Yazdi, M.K.; Taghizadeh, A.; Taghizadeh, M.; Stadler, F.J.; Farokhi, M.; Mottaghitalab, F.; Zarrintaj, P.; Ramsey, J.D.; Seidi, F.; Saeb, M.R. Agarose-based biomaterials for advanced drug delivery. J. Control. Release 2020, 326, 523–543. [Google Scholar] [CrossRef] [PubMed]
- López-Marcial, G.R.; Zeng, A.Y.; Osuna, C.; Dennis, J.; García, J.M.; O’Connell, G.D. Agarose-based hydrogels as suitable bioprinting materials for tissue engineering. ACS Biomater. Sci. Eng. 2018, 4, 3610–3616. [Google Scholar] [CrossRef] [PubMed]
- Numata, K. How to define and study structural proteins as biopolymer materials. Polym. J. 2020, 52, 1043–1056. [Google Scholar] [CrossRef]
- Jesionowski, T.; Norman, M.; Żółtowska-Aksamitowska, S.; Petrenko, I.; Joseph, Y.; Ehrlich, H. Marine spongin: Naturally prefabricated 3D scaffold-based biomaterial. Mar. Drugs 2018, 16, 88. [Google Scholar] [CrossRef] [PubMed]
- Andrade, P.; Portugal, L.; Rondon, E.; Toffoli-Kadri, M.; Cepa Matos, M.d.F. Effect of powdered shells treatment of the snail Megalobulimus lopesi on wounds of diabetic rats. Acta Cir. Bras. 2018, 33, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H.; Martinović, R.; Joksimović, D.; Petrenko, I.; Schiaparelli, S.; Wysokowski, M.; Tsurkan, D.; Stelling, A.L.; Springer, A.; Gelinsky, M.; et al. Conchixes: Organic scaffolds which resemble the size and shapes of mollusks shells, their isolation and potential multifunctional applications. Appl. Phys. A 2020, 126, 562. [Google Scholar] [CrossRef]
- Heldreth, B. Hydrolyzed Source Proteins as Used in Cosmetics. 2012. Available online: http://www.cir-safety.org/sites/default/files/hprtns052012slr.pdf (accessed on 9 December 2023).
- Latire, T.; Legendre, F.; Bigot, N.; Carduner, L.; Kellouche, S.; Bouyoucef, M.; Carreiras, F.; Marin, F.; Lebel, J.M.; Galéra, P.; et al. Shell extracts from the marine bivalve Pecten maximus regulate the synthesis of extracellular matrix in primary cultured human skin fibroblasts. PLoS ONE 2014, 9, e99931. [Google Scholar] [CrossRef]
- Latire, T.; Legendre, F.; Bouyoucef, M.; Marin, F.; Carreiras, F.; Rigot-Jolivet, M.; Lebel, J.-M.; Galéra, P.; Serpentini, A. Shell extracts of the edible mussel and oyster induce an enhancement of the catabolic pathway of human skin fibroblasts, in vitro. Cytotechnology 2017, 69, 815–829. [Google Scholar] [CrossRef]
- Yaqub, N.; Bullen, R.; Atkinson, H. Cleansing Composition. U.S. Patent US20040258650A1, 19 May 2015. [Google Scholar]
- Su, J.; Zhao, K.; Ren, Y.; Zhao, L.; Wei, B.; Liu, B.; Zhang, Y.; Wang, F.; Li, J.; Liu, Y. Biosynthetic structural proteins with super plasticity, extraordinary mechanical performance, biodegradability, biocompatibility and information storage ability. Angew. Chem. Int. Ed. 2022, 61, e202117538. [Google Scholar] [CrossRef]
- Rasheed, F. Production of Sustainable Bioplastic Materials from Wheat Gluten Proteins; Faculty of Landscape Planning, Horticulture and Agriculture Science: Uppsala, Sweden, 2011. [Google Scholar]
- Rasel, H.; Johansson, T.; Gällstedt, M.; Newson, W.; Johansson, E.; Hedenqvist, M. Development of bioplastics based on agricultural side-stream products: Film extrusion of Crambe abyssinica/wheat gluten blends for packaging purposes. J. Appl. Polym. Sci. 2016, 133, 42442. [Google Scholar] [CrossRef]
- Guerette, P.A.; Hoon, S.; Seow, Y.; Raida, M.; Masic, A.; Wong, F.T.; Ho, V.H.; Kong, K.W.; Demirel, M.C.; Pena-Francesch, A. Accelerating the design of biomimetic materials by integrating RNA-seq with proteomics and materials science. Nat. Biotechnol. 2013, 31, 908–915. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Ebrahimi, D.; Dinjaski, N.; Tarakanova, A.; Buehler, M.J.; Wong, J.Y.; Kaplan, D.L. Synergistic integration of experimental and simulation approaches for the de novo design of silk-based materials. Acc. Chem. Res. 2017, 50, 866–876. [Google Scholar] [CrossRef]
- Choi, K.R.; Jang, W.D.; Yang, D.; Cho, J.S.; Park, D.; Lee, S.Y. Systems metabolic engineering strategies: Integrating systems and synthetic biology with metabolic engineering. Trends Biotechnol. 2019, 37, 817–837. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.-X.; Qian, Z.-G.; Ki, C.S.; Park, Y.H.; Kaplan, D.L.; Lee, S.Y. Native-sized recombinant spider silk protein produced in metabolically engineered Escherichia coli results in a strong fiber. Proc. Natl. Acad. Sci. USA 2010, 107, 14059–14063. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Li, L.; Zhou, S. Microbial production of cyanophycin: From enzymes to biopolymers. Biotechnol. Adv. 2019, 37, 107400. [Google Scholar] [CrossRef]
- Simon, R.D.; Weathers, P. Determination of the structure of the novel polypeptide containing aspartic acid and arginine which is found in cyanobacteria. Biochim. Biophys. Acta (BBA)-Protein Struct. 1976, 420, 165–176. [Google Scholar] [CrossRef]
- Conrad, U. Polymers from plants to develop biodegradable plastics. Trends Plant Sci. 2005, 10, 511–512. [Google Scholar] [CrossRef]
- Gupta, M.; Carr, N. Enzyme activities related to cyanophycin metabolism in heterocysts and vegetative cells of Anabaena spp. Microbiology 1981, 125, 17–23. [Google Scholar] [CrossRef][Green Version]
- Obst, M.; Oppermann-Sanio, F.B.; Luftmann, H.; Steinbüchel, A. Isolation of cyanophycin-degrading bacteria, cloning and characterization of an extracellular cyanophycinase gene (cphE) from Pseudomonas anguilliseptica strain BI: The cphE gene from P. anguilliseptica BI encodes a cyanophycin-hydrolyzing enzyme. J. Biol. Chem. 2002, 277, 25096–25105. [Google Scholar] [CrossRef]
- Hellier, M.; Holdsworth, C.; McColl, I.; Perrett, D. Dipeptide absorption in man. Gut 1972, 13, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Bankar, S.B.; Nimbalkar, P.R.; Chavan, P.V.; Singhal, R.S. Microbial polyamino acids: An overview for commercial attention. In Role of Materials Science in Food Bioengineering, Handbook of Food Bioengineering; Academic Press: Cambridge, MA, USA, 2018; pp. 381–412. [Google Scholar]
- Hai, T.; Ahlers, H.; Gorenflo, V.; Steinbüchel, A. Axenic cultivation of anoxygenic phototrophic bacteria, cyanobacteria, and microalgae in a new closed tubular glass photobioreactor. Appl. Microbiol. Biotechnol. 2000, 53, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Sallam, A.; Steinbüchel, A. Dipeptides in nutrition and therapy: Cyanophycin-derived dipeptides as natural alternatives and their biotechnological production. Appl. Microbiol. Biotechnol. 2010, 87, 815–828. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.; Torcato, I.; Castanho, M.A. Biomedical applications of dipeptides and tripeptides. Pept. Sci. 2012, 98, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Frommeyer, M.; Wiefel, L.; Steinbüchel, A. Features of the biotechnologically relevant polyamide family “cyanophycins” and their biosynthesis in prokaryotes and eukaryotes. Crit. Rev. Biotechnol. 2016, 36, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Mooibroek, H.; Oosterhuis, N.; Giuseppin, M.; Toonen, M.; Franssen, H.; Scott, E.; Sanders, J.; Steinbüchel, A. Assessment of technological options and economical feasibility for cyanophycin biopolymer and high-value amino acid production. Appl. Microbiol. Biotechnol. 2007, 77, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Neumann, K.; Stephan, D.P.; Ziegler, K.; Hühns, M.; Broer, I.; Lockau, W.; Pistorius, E.K. Production of cyanophycin, a suitable source for the biodegradable polymer polyaspartate, in transgenic plants. Plant Biotechnol. J. 2005, 3, 249–258. [Google Scholar] [CrossRef]
- Andreeva, A.; Howorth, D.; Chothia, C.; Kulesha, E.; Murzin, A.G. SCOP2 prototype: A new approach to protein structure mining. Nucleic Acids Res. 2014, 42, 11847. [Google Scholar] [CrossRef]
- Shadwick, R.E. Elastic energy storage in tendons: Mechanical differences related to function and age. J. Appl. Physiol. 1990, 68, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Owczarzy, A.; Kurasiński, R.; Kulig, K.; Rogóż, W.; Szkudlarek, A.; Maciążek-Jurczyk, M. Collagen—Structure, properties and application. Eng. Biomater. 2020, 23, 17–23. [Google Scholar] [CrossRef]
- Safiya, N.; Casparus, J.R.V. Collagen: From Waste to Gold. In Biotechnological Applications of Biomass; Thalita Peixoto, B., Thiago Olitta, B., Luiz Carlos, B., Eds.; IntechOpen: Rijeka, Croatia, 2020. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2021, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Thilagavathi, G.; Viju, S. Silk as a suture material. In Advances in Silk Science and Technology; Woodhead Publishing: Cambridge, UK, 2015; pp. 219–232. [Google Scholar]
- Yu, H.-S.; Shen, Y.-H.; Yuan, G.-X.; Hu, Y.-G.; Xu, H.-E.; Xiang, Z.-H.; Zhang, Z. Evidence of selection at melanin synthesis pathway loci during silkworm domestication. Mol. Biol. Evol. 2011, 28, 1785–1799. [Google Scholar] [CrossRef] [PubMed]
- Babu, K.M. Silk: Processing, Properties and Applications; Woodhead Publishing: Cambridge, UK, 2018. [Google Scholar]
- Guo, Y.; Shen, Y.-H.; Sun, W.; Kishino, H.; Xiang, Z.-H.; Zhang, Z. Nucleotide diversity and selection signature in the domesticated silkworm, Bombyx mori, and wild silkworm, Bombyx mandarina. J. Insect Sci. 2011, 11, 155. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pereira, R.F.; Silva, M.M.; de Zea Bermudez, V. Bombyx mori silk fibers: An outstanding family of materials. Macromol. Mater. Eng. 2015, 300, 1171–1198. [Google Scholar] [CrossRef]
- Heim, M.; Keerl, D.; Scheibel, T. Spider Silk: From Soluble Protein to Extraordinary Fiber. Angew. Chem. Int. Ed. 2009, 48, 3584–3596. [Google Scholar] [CrossRef]
- Lu, W.; Shi, R.; Li, X.; Ma, S.; Yang, D.; Shang, D.; Xia, Q. A review on complete silk gene sequencing and de novo assembly of artificial silk. Int. J. Biol. Macromol. 2024, 264, 130444. [Google Scholar] [CrossRef]
- Hsia, Y.; Gnesa, E.; Jeffery, F.; Tang, S.; Vierra, C. Spider silk composites and applications. Met. Ceram. Polym. Compos. Var. Uses 2011, 2, 303–324. [Google Scholar]
- Kuhbier, J.W.; Reimers, K.; Kasper, C.; Allmeling, C.; Hillmer, A.; Menger, B.; Vogt, P.M.; Radtke, C. First investigation of spider silk as a braided microsurgical suture. J. Biomed. Mater. Res. Part. B Appl. Biomater. 2011, 97, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Kim, T.Y.; Lee, S.Y. Recent advances in production of recombinant spider silk proteins. Curr. Opin. Biotechnol. 2012, 23, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Keten, S.; Xu, Z.; Ihle, B.; Buehler, M.J. Nanoconfinement controls stiffness, strength and mechanical toughness of β-sheet crystals in silk. Nat. Mater. 2010, 9, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Yarger, J.L.; Cherry, B.R.; Van Der Vaart, A. Uncovering the structure–function relationship in spider silk. Nat. Rev. Mater. 2018, 3, 1–11. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, F.; Jiang, X.; Cao, M.; Wang, S.; Zou, H.; Cao, Y.; Xian, M.; Liu, H. Microbial production of amino acid-modified spider dragline silk protein with intensively improved mechanical properties. Prep. Biochem. Biotechnol. 2016, 46, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Copeland, C.G.; Bell, B.E.; Christensen, C.D.; Lewis, R.V. Development of a process for the spinning of synthetic spider silk. ACS Biomater. Sci. Eng. 2015, 1, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Deng, Q.; Liu, X.Y.; Yang, D. Engineered large spider eggcase silk protein for strong artificial fibers. Adv. Mater. 2013, 25, 1216–1220. [Google Scholar] [CrossRef] [PubMed]
- Adrianos, S.L.; Teulé, F.; Hinman, M.B.; Jones, J.A.; Weber, W.S.; Yarger, J.L.; Lewis, R.V. Nephila clavipes flagelliform silk-like GGX motifs contribute to extensibility and spacer motifs contribute to strength in synthetic spider silk fibers. Biomacromolecules 2013, 14, 1751–1760. [Google Scholar] [CrossRef]
- Andersson, M.; Jia, Q.; Abella, A.; Lee, X.-Y.; Landreh, M.; Purhonen, P.; Hebert, H.; Tenje, M.; Robinson, C.V.; Meng, Q. Biomimetic spinning of artificial spider silk from a chimeric minispidroin. Nat. Chem. Biol. 2017, 13, 262–264. [Google Scholar] [CrossRef]
- Cui, Y.; Gong, H.; Wang, Y.; Li, D.; Bai, H. A thermally insulating textile inspired by polar bear hair. Adv. Mater. 2018, 30, 1706807. [Google Scholar] [CrossRef]
- Yu, J.; Wei, W.; Danner, E.; Ashley, R.K.; Israelachvili, J.N.; Waite, J.H. Mussel protein adhesion depends on interprotein thiol-mediated redox modulation. Nat. Chem. Biol. 2011, 7, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Jin, S.; Park, Y.; Jung, Y.M.; Cha, H.J. Coacervation of interfacial adhesive proteins for initial mussel adhesion to a wet surface. Small 2018, 14, 1803377. [Google Scholar] [CrossRef] [PubMed]
- Kono, N.; Nakamura, H.; Ohtoshi, R.; Tomita, M.; Numata, K.; Arakawa, K. The bagworm genome reveals a unique fibroin gene that provides high tensile strength. Commun. Biol. 2019, 2, 148. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, T.; Tsubota, T.; Tashiro, K.; Jouraku, A.; Kameda, T. A study of the extraordinarily strong and tough silk produced by bagworms. Nat. Commun. 2019, 10, 1469. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Bachus, K.N.; Stewart, R.J. A water-borne adhesive modeled after the sandcastle glue of P. californica. Macromol. Biosci. 2009, 9, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Pena-Francesch, A.; Demirel, M.C. Squid-inspired tandem repeat proteins: Functional fibers and films. Front. Chem. 2019, 7, 69. [Google Scholar] [CrossRef] [PubMed]
- Tomko, J.A.; Pena-Francesch, A.; Jung, H.; Tyagi, M.; Allen, B.D.; Demirel, M.C.; Hopkins, P.E. Tunable thermal transport and reversible thermal conductivity switching in topologically networked bio-inspired materials. Nat. Nanotechnol. 2018, 13, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Sponner, A.; Vater, W.; Monajembashi, S.; Unger, E.; Grosse, F.; Weisshart, K. Composition and hierarchical organisation of a spider silk. PLoS ONE 2007, 2, e998. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sponner, A.; Porter, D.; Vollrath, F. Proline and processing of spider silks. Biomacromolecules 2008, 9, 116–121. [Google Scholar] [CrossRef]
- Nguyen, T.P.; Nguyen, Q.; Nguyen, V.-H.; Le, T.-H.; Huynh, V.; Vo, D.-V.; Trinh, Q.T.; Kim, S.Y.; Le, Q. Silk Fibroin-Based Biomaterials for Biomedical Applications: A Review. Polymers 2019, 11, 1933. [Google Scholar] [CrossRef]
- Shao, Z.; Vollrath, F. Surprising strength of silkworm silk. Nature 2002, 418, 741. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, D.; Adams, W.W.; Farmer, B.; Viney, C. Silk: Biology, Structure, Properties, and Genetics; ACS Publications: Washington, DC. USA, 1994. [Google Scholar]
- Vollrath, F.; Knight, D.P. Liquid crystalline spinning of spider silk. Nature 2001, 410, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Winkler, S.; Kaplan, D.L. Molecular biology of spider silk. Rev. Mol. Biotechnol. 2000, 74, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Foo, C.W.P.; Kaplan, D.L. Genetic engineering of fibrous proteins: Spider dragline silk and collagen. Adv. Drug Deliv. Rev. 2002, 54, 1131–1143. [Google Scholar]
- Altman, G.H.; Diaz, F.; Jakuba, C.; Calabro, T.; Horan, R.L.; Chen, J.; Lu, H.; Richmond, J.; Kaplan, D.L. Silk-based biomaterials. Biomaterials 2003, 24, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.Z.; Confalonieri, F.; Jacquet, M.; Perasso, R.; Li, Z.G.; Janin, J. Silk fibroin: Structural implications of a remarkable amino acid sequence. Proteins Struct. Funct. Bioinform. 2001, 44, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.-Z.; Confalonieri, F.; Medina, N.; Zivanovic, Y.; Esnault, C.; Yang, T.; Jacquet, M.; Janin, J.; Duguet, M.; Perasso, R. Fine organization of Bombyx mori fibroin heavy chain gene. Nucleic Acids Res. 2000, 28, 2413–2419. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, K.; Kikuchi, Y.; Takagi, T.; Kikuchi, A.; Oyama, F.; Shimura, K.; Mizuno, S. Primary structure of the silk fibroin light chain determined by cDNA sequencing and peptide analysis. J. Mol. Biol. 1989, 210, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.; Tanaka, K.; Arisaka, F.; Kimura, S.; Ohtomo, K.; Mizuno, S. Silk fibroin of Bombyx mori is secreted, assembling a high molecular mass elementary unit consisting of H-chain, L-chain, and P25, with a 6: 6: 1 molar ratio. J. Biol. Chem. 2000, 275, 40517–40528. [Google Scholar] [CrossRef]
- Vasconcelos, A.; Freddi, G.; Cavaco-Paulo, A. Biodegradable materials based on silk fibroin and keratin. Biomacromolecules 2008, 9, 1299–1305. [Google Scholar] [CrossRef]
- Bowen, C.H.; Dai, B.; Sargent, C.J.; Bai, W.; Ladiwala, P.; Feng, H.; Huang, W.; Kaplan, D.L.; Galazka, J.M.; Zhang, F. Recombinant Spidroins Fully Replicate Primary Mechanical Properties of Natural Spider Silk. Biomacromolecules 2018, 19, 3853–3860. [Google Scholar] [CrossRef] [PubMed]
- Bitar, L.; Isella, B.; Bertella, F.; Bettker Vasconcelos, C.; Harings, J.; Kopp, A.; van der Meer, Y.; Vaughan, T.J.; Bortesi, L. Sustainable Bombyx mori’s silk fibroin for biomedical applications as a molecular biotechnology challenge: A review. Int. J. Biol. Macromol. 2024, 264, 130374. [Google Scholar] [CrossRef] [PubMed]
- Bostan, F.; Surmeli, N.B. Cloning, expression, and characterization of a novel sericin-like protein. Biotechnol. Appl. Biochem. 2022, 69, 136–144. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Wang, Y.; Li, Q.; Liang, X.; Wang, G.; Li, J.; Peng, R.; Sima, Y.; Xu, S. Ectopic expression of sericin enables efficient production of ancient silk with structural changes in silkworm. Nat. Commun. 2022, 13, 6295. [Google Scholar] [CrossRef]
- Jin, H.-J.; Kaplan, D.L. Mechanism of silk processing in insects and spiders. Nature 2003, 424, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- Motta, A.; Fambri, L.; Migliaresi, C. Regenerated silk fibroin films: Thermal and dynamic mechanical analysis. Macromol. Chem. Phys. 2002, 203, 1658–1665. [Google Scholar] [CrossRef]
- Hümmerich, D.; Slotta, U.; Scheibel, T. Processing and modification of films made from recombinant spider silk proteins. Appl. Phys. A 2006, 82, 219–222. [Google Scholar] [CrossRef]
- Zhang, Y.-Q. Applications of natural silk protein sericin in biomaterials. Biotechnol. Adv. 2002, 20, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.R.; Kaplan, D.L. Biomedical applications of chemically-modified silk fibroin. J. Mater. Chem. 2009, 19, 6443–6450. [Google Scholar] [CrossRef]
- Matsumoto, A.; Chen, J.; Collette, A.L.; Kim, U.-J.; Altman, G.H.; Cebe, P.; Kaplan, D.L. Mechanisms of silk fibroin sol−gel transitions. J. Phys. Chem. B 2006, 110, 21630–21638. [Google Scholar] [CrossRef]
- Rockwood, D.N.; Preda, R.C.; Yücel, T.; Wang, X.; Lovett, M.L.; Kaplan, D.L. Materials fabrication from Bombyx mori silk fibroin. Nat. Protoc. 2011, 6, 1612–1631. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wang, X.; Rnjak, J.; Weiss, A.S.; Kaplan, D.L. Biomaterials derived from silk–tropoelastin protein systems. Biomaterials 2010, 31, 8121–8131. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-C.; Wang, S.-S.; Chou, N.-K.; Chi, N.-H.; Huang, Y.-Y.; Chang, Y.-L.; Shieh, M.-J.; Chung, T.-W. The cardiomyogenic differentiation of rat mesenchymal stem cells on silk fibroin–polysaccharide cardiac patches in vitro. Biomaterials 2009, 30, 3757–3765. [Google Scholar] [CrossRef] [PubMed]
- Unger, R.E.; Wolf, M.; Peters, K.; Motta, A.; Migliaresi, C.; Kirkpatrick, C.J. Growth of human cells on a non-woven silk fibroin net: A potential for use in tissue engineering. Biomaterials 2004, 25, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Niamsa, N.; Srisuwan, Y.; Baimark, Y.; Phinyocheep, P.; Kittipoom, S. Preparation of nanocomposite chitosan/silk fibroin blend films containing nanopore structures. Carbohydr. Polym. 2009, 78, 60–65. [Google Scholar] [CrossRef]
- Um, I.C.; Kweon, H.; Park, Y.H.; Hudson, S. Structural characteristics and properties of the regenerated silk fibroin prepared from formic acid. Int. J. Biol. Macromol. 2001, 29, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Um, I.C.; Kweon, H.Y.; Lee, K.G.; Park, Y.H. The role of formic acid in solution stability and crystallization of silk protein polymer. Int. J. Biol. Macromol. 2003, 33, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Min, B.-M.; Jeong, L.; Nam, Y.S.; Kim, J.-M.; Kim, J.Y.; Park, W.H. Formation of silk fibroin matrices with different texture and its cellular response to normal human keratinocytes. Int. J. Biol. Macromol. 2004, 34, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kim, H.-J.; Vunjak-Novakovic, G.; Kaplan, D.L. Stem cell-based tissue engineering with silk biomaterials. Biomaterials 2006, 27, 6064–6082. [Google Scholar] [CrossRef]
- Kundu, B.; Rajkhowa, R.; Kundu, S.C.; Wang, X. Silk fibroin biomaterials for tissue regenerations. Adv. Drug Deliv. Rev. 2013, 65, 457–470. [Google Scholar] [CrossRef]
- Chen, X.; Li, W.; Zhong, W.; Lu, Y.; Yu, T. pH sensitivity and ion sensitivity of hydrogels based on complex-forming chitosan/silk fibroin interpenetrating polymer network. J. Appl. Polym. Sci. 1997, 65, 2257–2262. [Google Scholar] [CrossRef]
- Haider, Z.A.; Arai, M.; Hirabayashi, K. Mechanism of the gelation of fibroin solution. Biosci. Biotechnol. Biochem. 1993, 57, 1910–1912. [Google Scholar]
- Hanawa, T.; Watanabe, A.; Tsuchiya, T.; Ikoma, R.; Hidaka, M.; Sugihara, M. New oral dosage form for elderly patients: Preparation and characterization of silk fibroin gel. Chem. Pharm. Bull. 1995, 43, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Kim, U.-J.; Park, J.; Li, C.; Jin, H.-J.; Valluzzi, R.; Kaplan, D.L. Structure and properties of silk hydrogels. Biomacromolecules 2004, 5, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.; Yoo, M.; Seo, J.; Park, S.; Na, H.; Lee, H.; Kim, S.; Cho, C. Evaluation of semi-interpenetrating polymer networks composed of chitosan and poloxamer for wound dressing application. Int. J. Pharm. 2007, 341, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Brinckmann, J.; Hunzelmann, N.; Kahle, B.; Rohwedel, J.; Kramer, J.; Gibson, M.A.; Hubmacher, D.; Reinhardt, D.P. Enhanced fibrillin-2 expression is a general feature of wound healing and sclerosis: Potential alteration of cell attachment and storage of TGF-β. Lab. Investig. 2010, 90, 739–752. [Google Scholar] [CrossRef] [PubMed]
- Mariencheck, M.C.; Davis, E.C.; Zhang, H.; Ramirez, F.; Rosenbloom, J.; Gibson, M.A.; Parks, W.C.; Mecham, R.P. Fibrillin-1 and Fibrillin-2 Show Temporal and Tissue-Specific Regulation of Expression in Developing Elastic Tissues. Connect. Tissue Res. 1995, 31, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Handford, P.; Downing, A.K.; Rao, Z.; Hewett, D.R.; Sykes, B.C.; Kielty, C.M. The calcium binding properties and molecular organization of epidermal growth factor-like domains in human fibrillin-1. J. Biol. Chem. 1995, 270, 6751–6756. [Google Scholar] [CrossRef] [PubMed]
- Bragulla, H.H.; Homberger, D.G. Structure and functions of keratin proteins in simple, stratified, keratinized and cornified epithelia. J. Anat. 2009, 214, 516–559. [Google Scholar] [CrossRef]
- Wang, L.; Shang, Y.; Zhang, J.; Yuan, J.; Shen, J. Recent advances in keratin for biomedical applications. Adv. Colloid. Interface Sci. 2023, 321, 103012. [Google Scholar] [CrossRef]
- Schweizer, J.; Bowden, P.E.; Coulombe, P.A.; Langbein, L.; Lane, E.B.; Magin, T.M.; Maltais, L.; Omary, M.B.; Parry, D.A.; Rogers, M.A.; et al. New consensus nomenclature for mammalian keratins. J. Cell Biol. 2006, 174, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Hardy, M.H. The secret life of the hair follicle. Trends Genet. 1992, 8, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Panteleyev, A.A.; Jahoda, C.A.; Christiano, A.M. Hair follicle predetermination. J. Cell Sci. 2001, 114, 3419–3431. [Google Scholar] [CrossRef] [PubMed]
- Popescu, C.; Höcker, H. Hair—The most sophisticated biological composite material. Chem. Soc. Rev. 2007, 36, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Parry, D.A.; Steinert, P.M. Intermediate filament structure. Curr. Opin. Cell Biol. 1992, 4, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Hill, P.; Brantley, H.; Van Dyke, M. Some properties of keratin biomaterials: Kerateines. Biomaterials 2010, 31, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Plowman, J.E. Proteomic database of wool components. J. Chromatogr. B 2003, 787, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, A.; Cavaco-Paulo, A. Wound dressings for a proteolytic-rich environment. Appl. Microbiol. Biotechnol. 2011, 90, 445–460. [Google Scholar] [CrossRef] [PubMed]
- Smack, D.P.; Korge, B.P.; James, W.D. Keratin and keratinization. J. Am. Acad. Dermatol. 1994, 30, 85–102. [Google Scholar] [CrossRef]
- Maclaren, J. The extent of reduction of wool proteins by thiols. Aust. J. Chem. 1962, 15, 824–831. [Google Scholar] [CrossRef]
- O’donnell, I.; Thompson, E. Studies on reduced wool. Aust. J. Biol. Sci. 1964, 17, 973–978. [Google Scholar] [CrossRef]
- Zahn, H. Progress report on hair keratin research. Int. J. Cosmet. Sci. 2002, 24, 163. [Google Scholar] [CrossRef] [PubMed]
- Tonin, C.; Aluigi, A.; Vineis, C.; Varesano, A.; Montarsolo, A.; Ferrero, F. Thermal and structural characterization of poly (ethylene-oxide)/keratin blend films. J. Therm. Anal. Calorim. 2007, 89, 601–608. [Google Scholar] [CrossRef]
- Coulombe, P.A.; Chan, Y.; Albers, K.; Fuchs, E. Deletions in epidermal keratins leading to alterations in filament organization in vivo and in intermediate filament assembly in vitro. J. Cell Biol. 1990, 111, 3049–3064. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.; Conrads, A.; Phan, K.-H.; van de Löcht, M.; Zahn, H. In vitro reconstitution of wool intermediate filaments. Int. J. Biol. Macromol. 1986, 8, 258–264. [Google Scholar] [CrossRef]
- Ikkai, F.; Naito, S. Dynamic light scattering and circular dichroism studies on heat-induced gelation of hard-keratin protein aqueous solutions. Biomacromolecules 2002, 3, 482–487. [Google Scholar] [CrossRef]
- Rouse, J.G.; Van Dyke, M.E. A review of keratin-based biomaterials for biomedical applications. Materials 2010, 3, 999–1014. [Google Scholar] [CrossRef]
- Kawano, Y.; Okamoto, S. Film and gel of keratins. Kagaku Seibutsu 1975, 13, 291–292. [Google Scholar]
- Yeo, J.; Jung, G.; Tarakanova, A.; Martín-Martínez, F.J.; Qin, Z.; Cheng, Y.; Zhang, Y.-W.; Buehler, M.J. Multiscale modeling of keratin, collagen, elastin and related human diseases: Perspectives from atomistic to coarse-grained molecular dynamics simulations. Extrem. Mech. Lett. 2018, 20, 112–124. [Google Scholar] [CrossRef]
- Costa, F.; Silva, R.; Boccaccini, A. Fibrous protein-based biomaterials (silk, keratin, elastin, and resilin proteins) for tissue regeneration and repair. In Peptides and Proteins as Biomaterials for Tissue Regeneration and Repair; Woodhead Publishing: Cambridge, UK, 2018; pp. 175–204. [Google Scholar]
- Zhou, Z.; Zhang, S.; Cao, Y.; Marelli, B.; Xia, X.; Tao, T.H. Engineering the future of silk materials through advanced manufacturing. Adv. Mater. 2018, 30, 1706983. [Google Scholar] [CrossRef]
- Zhu, B.; Wang, H.; Leow, W.R.; Cai, Y.; Loh, X.J.; Han, M.Y.; Chen, X. Silk fibroin for flexible electronic devices. Adv. Mater. 2016, 28, 4250–4265. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Ling, S.; Li, C.; Omenetto, F.G.; Kaplan, D.L. Silkworm silk-based materials and devices generated using bio-nanotechnology. Chem. Soc. Rev. 2018, 47, 6486–6504. [Google Scholar] [CrossRef] [PubMed]
- Koh, L.-D.; Yeo, J.; Lee, Y.Y.; Ong, Q.; Han, M.; Tee, B.C. Advancing the frontiers of silk fibroin protein-based materials for futuristic electronics and clinical wound-healing (Invited review). Mater. Sci. Eng. C 2018, 86, 151–172. [Google Scholar] [CrossRef] [PubMed]
- DeFrates, K.G.; Moore, R.; Borgesi, J.; Lin, G.; Mulderig, T.; Beachley, V.; Hu, X. Protein-Based Fiber Materials in Medicine: A Review. Nanomaterials 2018, 8, 457. [Google Scholar] [CrossRef] [PubMed]
- Grewell, D.; Schrader, J.; Srinivasan, G. Protein-based Plastics. In Bioplastics and Biocomposites: A Practical Introduction; Grewell, D., Ed.; Royal Society of Chemistry: Cambridge, UK, 2023; Volume 79. [Google Scholar]
- Kenneth, B.; Torkel, W.-F. Amino acid composition of a new rubber-like protein, resilin. Biochim. Biophys. Acta 1961, 48, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-C.; Fan, R.-X.; Tian, K.-K.; Xia, X.-X.; Qian, Z.-G. Controllable fibrillization reinforces genetically engineered rubberlike protein hydrogels. Biomacromolecules 2021, 22, 961–970. [Google Scholar] [CrossRef]
- Balu, R.; Dutta, N.K.; Dutta, A.K.; Choudhury, N.R. Resilin-mimetics as a smart biomaterial platform for biomedical applications. Nat. Commun. 2021, 12, 149. [Google Scholar] [CrossRef] [PubMed]
- Sage, E.; Gray, W. Evolution of elastin structure. In Elastin and Elastic Tissue; Springer: Berlin/Heidelberg, Germany, 1977; pp. 291–312. [Google Scholar]
- Mithieux, S.M.; Rasko, J.E.; Weiss, A.S. Synthetic elastin hydrogels derived from massive elastic assemblies of self-organized human protein monomers. Biomaterials 2004, 25, 4921–4927. [Google Scholar] [CrossRef] [PubMed]
- Lillie, M.; Gosline, J. The viscoelastic basis for the tensile strength of elastin. Int. J. Biol. Macromol. 2002, 30, 119–127. [Google Scholar] [CrossRef]
- Rauscher, S.; Pomès, R. Structural disorder and protein elasticity. In Fuzziness. Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2012; pp. 159–183. [Google Scholar]
- Muiznieks, L.D.; Weiss, A.S.; Keeley, F.W. Structural disorder and dynamics of elastin. Biochem. Cell Biol. 2010, 88, 239–250. [Google Scholar] [CrossRef]
- Muiznieks, L.D.; Weiss, A.S. Flexibility in the solution structure of human tropoelastin. Biochemistry 2007, 46, 8196–8205. [Google Scholar] [CrossRef] [PubMed]
- Toonkool, P.; Regan, D.G.; Kuchel, P.W.; Morris, M.B.; Weiss, A.S. Thermodynamic and hydrodynamic properties of human tropoelastin: Analytical ultracentrifuge and pulsed field-gradient spin-echo NMR studies. J. Biol. Chem. 2001, 276, 28042–28050. [Google Scholar] [CrossRef] [PubMed]
- Baldock, C.; Oberhauser, A.F.; Ma, L.; Lammie, D.; Siegler, V.; Mithieux, S.M.; Tu, Y.; Chow, J.Y.H.; Suleman, F.; Malfois, M. Shape of tropoelastin, the highly extensible protein that controls human tissue elasticity. Proc. Natl. Acad. Sci. USA 2011, 108, 4322–4327. [Google Scholar] [CrossRef] [PubMed]
- Pometun, M.S.; Chekmenev, E.Y.; Wittebort, R.J. Quantitative observation of backbone disorder in native elastin. J. Biol. Chem. 2004, 279, 7982–7987. [Google Scholar] [CrossRef]
- Kozel, B.A.; Rongish, B.J.; Czirok, A.; Zach, J.; Little, C.D.; Davis, E.C.; Knutsen, R.H.; Wagenseil, J.E.; Levy, M.A.; Mecham, R.P. Elastic fiber formation: A dynamic view of extracellular matrix assembly using timer reporters. J. Cell. Physiol. 2006, 207, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Tamburro, A.M.; Pepe, A.; Bochicchio, B.; Quaglino, D.; Ronchetti, I.P. Supramolecular amyloid-like assembly of the polypeptide sequence coded by exon 30 of human tropoelastin. J. Biol. Chem. 2005, 280, 2682–2690. [Google Scholar] [CrossRef]
- Miao, M.; Bellingham, C.M.; Stahl, R.J.; Sitarz, E.E.; Lane, C.J.; Keeley, F.W. Sequence and structure determinants for the self-aggregation of recombinant polypeptides modeled after human elastin. J. Biol. Chem. 2003, 278, 48553–48562. [Google Scholar] [CrossRef]
- Hoeve, C.; Flory, P. The elastic properties of elastin. Biopolym. Orig. Res. Biomol. 1974, 13, 677–686. [Google Scholar] [CrossRef]
- Burrows, M.; Shaw, S.R.; Sutton, G.P. Resilin and chitinous cuticle form a composite structure for energy storage in jumping by froghopper insects. BMC Biol. 2008, 6, 41. [Google Scholar] [CrossRef]
- Bennet-Clark, H.; Lucey, E. The jump of the flea: A study of the energetics and a model of the mechanism. J. Exp. Biol. 1967, 47, 59–76. [Google Scholar] [CrossRef]
- Weis-Fogh, T. A rubber-like protein in insect cuticle. J. Exp. Biol. 1960, 37, 889–907. [Google Scholar] [CrossRef]
- Tamburro, A.M.; Panariello, S.; Santopietro, V.; Bracalello, A.; Bochicchio, B.; Pepe, A. Molecular and Supramolecular Structural Studies on Significant Repetitive Sequences of Resilin. ChemBioChem 2010, 11, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Song, J.H.; Murphy, R.J.; Narayan, R.; Davies, G.B. Biodegradable and compostable alternatives to conventional plastics. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2127–2139. [Google Scholar] [CrossRef] [PubMed]
- Grewell, D.; Schrader, J.; Srinivasan, G. Developing Protein-Based Plastics. In Soy-Based Chemicals and Materials, 1st ed.; Brentin, R.P., Ed.; American Chemical Society: Washington, DC, USA, 2014; Volume 1178. [Google Scholar]
- Sugiyama, T.; Rafalski, A.; Peterson, D.; Söll, D. A wheat HMW glutenin subunit gene reveals a highly repeated structure. Nucleic Acids Res. 1985, 13, 8729–8737. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Halfmann, C.; Gu, L.; Zhou, R. Engineering cyanobacteria for the production of a cyclic hydrocarbon fuel from CO2 and H2O. Green. Chem. 2014, 16, 3175–3185. [Google Scholar] [CrossRef]
- Elhai, J.; Wolk, C.P. Conjugal transfer of DNA to cyanobacteria. Methods Enzymol. 1988, 167, 747–754. [Google Scholar] [PubMed]
- Kubiak, A.; Pajewska-Szmyt, M.; Kotula, M.; Leśniewski, B.; Voronkina, A.; Rahimi, P.; Falahi, S.; Heimler, K.; Rogoll, A.; Vogt, C.; et al. Spongin as a Unique 3D Template for the Development of Functional Iron-Based Composites Using Biomimetic Approach In Vitro. Mar. Drugs 2023, 21, 460. [Google Scholar] [CrossRef]
- Ehrlich, H. Biological Materials of Marine Origin; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Petrenko, I.; Summers, A.P.; Simon, P.; Żółtowska-Aksamitowska, S.; Motylenko, M.; Schimpf, C.; Rafaja, D.; Roth, F.; Kummer, K.; Brendler, E.; et al. Extreme biomimetics: Preservation of molecular detail in centimeter-scale samples of biological meshes laid down by sponges. Sci. Adv. 2019, 5, eaax2805. [Google Scholar] [CrossRef]
- Ashouri, V.; Rahimi-Nasrabadi, M.; Attaran, G.; Adib, K.; Zahedi, M.; Ganjali, M.; Marzi, E. Extraction and pre-concentration of ketamine by using a three-dimensional spongin-based scaffold of the Haliclona sp. marine demosponge origin. Appl. Phys. A 2020, 126, 421. [Google Scholar] [CrossRef]
- Yang, S.; Wang, Z.; Hu, Y.; Zong, K.; Zhang, X.; Ke, H.; Wang, P.; Go, Y.; Chan, X.H.F.; Wu, J.; et al. Hydrolyzed Conchiolin Protein (HCP) Extracted from Pearls Antagonizes both ET-1 and α-MSH for Skin Whitening. Int. J. Mol. Sci. 2023, 24, 7471. [Google Scholar] [CrossRef]
- Chen, S.; Huang, S.; Li, Y.; Zhou, C. Recent Advances in Epsilon-Poly-L-Lysine and L-Lysine-Based Dendrimer Synthesis, Modification, and Biomedical Applications. Front. Chem. 2021, 9, 659304. [Google Scholar] [CrossRef] [PubMed]
- Björn, W.; Karl, F. Cyanophycin: A Nitrogen-Rich Reserve Polymer; IntechOpen: Winchester, UK, 2018. [Google Scholar] [CrossRef]
- Ziegler, K.; Diener, A.; Herpin, C.; Richter, R.; Deutzmann, R.; Lockau, W. Molecular characterization of cyanophycin synthetase, the enzyme catalyzing the biosynthesis of the cyanobacterial reserve material multi-L-arginyl-poly-L-aspartate (cyanophycin). Eur. J. Biochem. 1998, 254, 154–159. [Google Scholar] [CrossRef]
- Sharon, I.; Haque, A.S.; Grogg, M.; Lahiri, I.; Seebach, D.; Leschziner, A.E.; Hilvert, D.; Schmeing, T.M. Structures and function of the amino acid polymerase cyanophycin synthetase. Nat. Chem. Biol. 2021, 17, 1101–1110. [Google Scholar] [CrossRef] [PubMed]
- Steinle, A.; Steinbüchel, A. Establishment of a simple and effective isolation method for cyanophycin from recombinant Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2010, 85, 1393–1399. [Google Scholar] [CrossRef]
- Abd-El-Karem, Y.; Elbers, T.; Reichelt, R.; Steinbüchel, A. Heterologous expression of Anabaena sp. PCC7120 cyanophycin metabolism genes cphA1 and cphB1 in Sinorhizobium (Ensifer) meliloti 1021. Appl. Microbiol. Biotechnol. 2011, 89, 1177–1192. [Google Scholar] [CrossRef]
- Fokina, O.; Chellamuthu, V.R.; Zeth, K.; Forchhammer, K. A Novel Signal Transduction Protein P-II Variant from Synechococcus elongatus PCC 7942 Indicates a Two-Step Process for NAGK-P-II Complex Formation. J. Mol. Biol. 2010, 399, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Watzer, B.; Engelbrecht, A.; Hauf, W.; Stahl, M.; Maldener, I.; Forchhammer, K. Metabolic pathway engineering using the central signal processor P-II. Microb. Cell Fact. 2015, 14, 192. [Google Scholar] [CrossRef] [PubMed]
- Trautmann, A.; Watzer, B.; Wilde, A.; Forchhammer, K.; Posten, C. Effect of phosphate availability on cyanophycin accumulation in Synechocystis sp PCC 6803 and the production strain BW86. Algal. Res. 2016, 20, 189–196. [Google Scholar] [CrossRef]
- Khlystov, N.A.; Chan, W.Y.; Kunjapur, A.M.; Shi, W.C.; Prather, K.U.; Olsen, B.D. Material properties of the cyanobacterial reserve polymer multi-L- arginyl-poly-L-aspartate (cyanophycin). Polymer 2017, 109, 238–245. [Google Scholar] [CrossRef]
- Kwiatos, N.; Steinbüchel, A. Cyanophycin Modifications—Widening the Application Potential. Front. Bioeng. Biotechnol. 2021, 9, 763804. [Google Scholar] [CrossRef]
- Frey, K.M.; Oppermann-Sanio, F.B.; Schmidt, H.; Steinbüchel, A. Technical-scale production of cyanophycin with recombinant strains of Escherichia coli. Appl. Environ. Microbiol. 2002, 68, 3377–3384. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Koch, M.; Forchhammer, K. Cyanobacterial Biopolymers. In Cyanobacteria Biotechnology; John Wiley & Sons: Hoboken, NJ, USA, 2021; pp. 301–330. [Google Scholar] [CrossRef]
- Halfmann, C.; Gu, L.; Gibbons, W.; Zhou, R. Genetically engineering cyanobacteria to convert CO2, water, and light into the long-chain hydrocarbon farnesene. Appl. Microbiol. Biotechnol. 2014, 98, 9869–9877. [Google Scholar] [CrossRef] [PubMed]
- Halfmann, C.T. The Synthetic Biology of N 2-Fixing Cyanobacteria for Photosynthetic Terpenoid Production. Ph.D. Dissertation, South Dakota State University, Brookings, SD, USA, 2017. [Google Scholar]
- Gu, L.; Xiang, X.; Raynie, D.; Gibbons, W.; Zhou, R. Biosolar Conversion of CO2 and H2O into Long-chain alcohol. In Proceedings of the Sun Grant National Conference, Science for Biomass Feedstock Production and Utilization, New Orleans, LA, USA, 2–5 October 2012. [Google Scholar]
- Chen, K.; Zhu, H.; Gu, L.; Tian, S.; Zhou, R. Target Gene Inactivation in Cyanobacterium Anabaena sp. PCC 7120. Bio-Protocol 2016, 6, e1890. [Google Scholar] [CrossRef]
- Xu, X.; Gu, L.; He, P.; Zhou, R. Characterization of five putative aspartate aminotransferase genes in the N2-fixing heterocystous cyanobacterium Anabaena sp. strain PCC 7120. Microbiology 2015, 161, 1219–1230. [Google Scholar] [CrossRef]
- Zhou, R.B.; Wolk, C.P. A two-component system mediates developmental regulation of biosynthesis of a heterocyst polysaccharide. J. Biol. Chem. 2003, 278, 19939–19946. [Google Scholar] [CrossRef]
- Chen, K.; Xu, X.; Gu, L.; Hildreth, M.; Zhou, R. Simultaneous gene inactivation and promoter reporting in cyanobacteria. Appl. Microbiol. Biotechnol. 2015, 99, 1779–1793. [Google Scholar] [CrossRef] [PubMed]
- Oppermann-Sanio, F.B.; Steinbüchel, A. Occurrence, functions and biosynthesis of polyamides in microorganisms and biotechnological production. Naturwissenschaften 2002, 89, 11–22. [Google Scholar] [CrossRef]
- Obst, M.; Sallam, A.; Luftmann, H.; Steinbüchel, A. Isolation and Characterization of Gram-Positive Cyanophycin-Degrading Bacteria Kinetic Studies on Cyanophycin Depolymerase Activity in Aerobic Bacteria. Biomacromolecules 2004, 5, 153–161. [Google Scholar] [CrossRef]
- Sallam, A.; Kast, A.; Przybilla, S.; Meiswinkel, T.; Steinbüchel, A. Biotechnological process for production of β-dipeptides from cyanophycin on a technical scale and its optimization. Appl. Environ. Microbiol. 2009, 75, 29–38. [Google Scholar] [CrossRef]
- Olechnovič, K.; Venclovas, Č. VoroMQA web server for assessing three-dimensional structures of proteins and protein complexes. Nucleic Acids Res. 2019, 47, W437–W442. [Google Scholar] [CrossRef]
- Pickel, B.; Schaller, A. Dirigent proteins: Molecular characteristics and potential biotechnological applications. Appl. Microbiol. Biotechnol. 2013, 97, 8427–8438. [Google Scholar] [CrossRef]
- Satarker, S.; Nampoothiri, M. Structural proteins in severe acute respiratory syndrome coronavirus-2. Arch. Med. Res. 2020, 51, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Z.; Jeon, J.; Jiang, B.; Subramani, S.V.; Li, J.; Zhang, F. Protein-Based Hydrogels and Their Biomedical Applications. Molecules 2023, 28, 4988. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Qi, X.; Chen, Y.; Wu, Z. Thermo-sensitive hydrogels for delivering biotherapeutic molecules: A review. Saudi Pharm. J. 2019, 27, 990–999. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Jia, X.; Zhang, Q.; Chen, H.; Zhu, Q.; Yin, L. Interpenetrating polymer network hydrogels of soy protein isolate and sugar beet pectin as a potential carrier for probiotics. Food Hydrocoll. 2021, 113, 106453. [Google Scholar] [CrossRef]
- Joseph, C.A.; McCarthy, C.W.; Tyo, A.G.; Hubbard, K.R.; Fisher, H.C.; Altscheffel, J.A.; He, W.; Pinnaratip, R.; Liu, Y.; Lee, B.P. Development of an injectable nitric oxide releasing poly (ethylene) glycol-Fibrin adhesive hydrogel. ACS Biomater. Sci. Eng. 2018, 5, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Zhang, B.; Yadav, M.P.; Feng, L.; Yan, J.; Jia, X.; Yin, L. Corn fiber gum-soybean protein isolate double network hydrogel as oral delivery vehicles for thermosensitive bioactive compounds. Food Hydrocoll. 2020, 107, 105865. [Google Scholar] [CrossRef]
- Li, Z.; Xu, W.; Wang, X.; Jiang, W.; Ma, X.; Wang, F.; Zhang, C.; Ren, C. Fabrication of PVA/PAAm IPN hydrogel with high adhesion and enhanced mechanical properties for body sensors and antibacterial activity. Eur. Polym. J. 2021, 146, 110253. [Google Scholar] [CrossRef]
- Li, Z.; Meng, X.; Xu, W.; Zhang, S.; Ouyang, J.; Zhang, Z.; Liu, Y.; Niu, Y.; Ma, S.; Xue, Z. Single network double cross-linker (SNDCL) hydrogels with excellent stretchability, self-recovery, adhesion strength, and conductivity for human motion monitoring. Soft Matter 2020, 16, 7323–7331. [Google Scholar] [CrossRef]
- Huang, S.; Kong, X.; Xiong, Y.; Zhang, X.; Chen, H.; Jiang, W.; Niu, Y.; Xu, W.; Ren, C. An overview of dynamic covalent bonds in polymer material and their applications. Eur. Polym. J. 2020, 141, 110094. [Google Scholar] [CrossRef]
- Lim, S.; Jung, G.A.; Muckom, R.J.; Glover, D.J.; Clark, D.S. Engineering bioorthogonal protein–polymer hybrid hydrogel as a functional protein immobilization platform. Chem. Commun. 2019, 55, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Najmi, L.; Hu, Z. Review on Molecular Dynamics Simulations of Effects of Carbon Nanotubes (CNTs) on Electrical and Thermal Conductivities of CNT-Modified Polymeric Composites. J. Compos. Sci. 2023, 7, 165. [Google Scholar] [CrossRef]
- Hu, Z.; Hong, H. Review on Material Performance of Carbon Nanotube-Modified Polymeric Nanocomposites. Recent. Prog. Mater. 2023, 5, 1–20. [Google Scholar] [CrossRef]
- Berger, J.; Reist, M.; Mayer, J.; Felt, O.; Peppas, N.; Gurny, R. Structure and interactions in covalently and ionically crosslinked chitosan hydrogels for biomedical applications. Eur. J. Pharm. Biopharm. 2004, 57, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Xia, X.-X.; Huang, S.-C.; Qian, Z.-G. Development of adhesive and conductive resilin-based hydrogels for wearable sensors. Biomacromolecules 2019, 20, 3283–3293. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Han, G.; Wang, Q.; Zhang, S.; You, R.; Luo, Z.; Xu, A.; Li, X.; Li, M.; Zhang, Q. Directed assembly of robust and biocompatible silk fibroin/hyaluronic acid composite hydrogels. Compos. Part. B Eng. 2019, 176, 107204. [Google Scholar] [CrossRef]
- Hu, X.; Lu, Q.; Sun, L.; Cebe, P.; Wang, X.; Zhang, X.; Kaplan, D.L. Biomaterials from ultrasonication-induced silk fibroin− hyaluronic acid hydrogels. Biomacromolecules 2010, 11, 3178–3188. [Google Scholar] [CrossRef]
- Tang, Z.; Chen, Q.; Chen, F.; Zhu, L.; Lu, S.; Ren, B.; Zhang, Y.; Yang, J.; Zheng, J. General principle for fabricating natural globular protein-based double-network hydrogels with integrated highly mechanical properties and surface adhesion on solid surfaces. Chem. Mater. 2018, 31, 179–189. [Google Scholar] [CrossRef]
- Xu, R.; Ma, S.; Lin, P.; Yu, B.; Zhou, F.; Liu, W. High strength astringent hydrogels using protein as the building block for physically cross-linked multi-network. ACS Appl. Mater. Interfaces 2017, 10, 7593–7601. [Google Scholar] [CrossRef]
- Dai, X.; Zhang, Y.; Gao, L.; Bai, T.; Wang, W.; Cui, Y.; Liu, W. A Mechanically Strong, Highly Stable, Thermoplastic, and self-healable supramolecular polymer hydrogel. Adv. Mater. 2015, 27, 3566–3571. [Google Scholar] [CrossRef]
- Hoff, S.E.; Liu, J.; Heinz, H. Binding mechanism and binding free energy of amino acids and citrate to hydroxyapatite surfaces as a function of crystallographic facet, pH, and electrolytes. J. Colloid. Interface Sci. 2022, 605, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wu, T.; Wang, M.; Shi, S.; Yuan, G.; Li, X.; Chong, H.; Wu, B.; Zheng, P. Enzymatic biosynthesis and immobilization of polyprotein verified at the single-molecule level. Nat. Commun. 2019, 10, 2775. [Google Scholar] [CrossRef] [PubMed]
- Fisher, S.A.; Baker, A.E.; Shoichet, M.S. Designing peptide and protein modified hydrogels: Selecting the optimal conjugation strategy. J. Am. Chem. Soc. 2017, 139, 7416–7427. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gao, W. Precision conjugation: An emerging tool for generating protein–polymer conjugates. Angew. Chem. Int. Ed. 2021, 60, 11024–11035. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Zhang, W.-B.; Mahdavi, A.; Arnold, F.H.; Tirrell, D.A. Synthesis of bioactive protein hydrogels by genetically encoded SpyTag-SpyCatcher chemistry. Proc. Natl. Acad. Sci. USA 2014, 111, 11269–11274. [Google Scholar] [CrossRef] [PubMed]
- Lyu, S.; Fang, J.; Duan, T.; Fu, L.; Liu, J.; Li, H. Optically controlled reversible protein hydrogels based on photoswitchable fluorescent protein Dronpa. Chem. Commun. 2017, 53, 13375–13378. [Google Scholar] [CrossRef] [PubMed]
- Fierer, J.O.; Veggiani, G.; Howarth, M. SpyLigase peptide–peptide ligation polymerizes affibodies to enhance magnetic cancer cell capture. Proc. Natl. Acad. Sci. USA 2014, 111, E1176–E1181. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Wang, Z.; Xiao, Y.; Zhang, S.; Wang, J. Advances in crosslinking strategies of biomedical hydrogels. Biomater. Sci.-UK 2019, 7, 843–855. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yang, L.; He, Y.; Xiao, H.; Lin, W. Microsphere-structured hydrogel crosslinked by polymerizable protein-based nanospheres. Polymer 2020, 211, 123114. [Google Scholar] [CrossRef]
- Su, J.; Lu, S.; Jiang, S.; Li, B.; Liu, B.; Sun, Q.; Li, J.; Wang, F.; Wei, Y. Engineered protein photo-thermal hydrogels for outstanding in situ tongue cancer therapy. Adv. Mater. 2021, 33, 2100619. [Google Scholar] [CrossRef]
- Wang, Y.; Xia, Y.; Xiang, P.; Dai, Y.; Gao, Y.; Xu, H.; Yu, J.; Gao, G.; Chen, K. Protein-assisted freeze-tolerant hydrogel with switchable performance toward customizable flexible sensor. Chem. Eng. J. 2022, 428, 131171. [Google Scholar] [CrossRef]
- Boonpavanitchakul, K.; Bast, L.K.; Bruns, N.; Magaraphan, R. Silk sericin-polylactide protein–polymer conjugates as biodegradable amphiphilic materials and their application in drug release systems. Bioconjugate Chem. 2020, 31, 2312–2324. [Google Scholar] [CrossRef]
- Lei, H.; Dong, L.; Li, Y.; Zhang, J.; Chen, H.; Wu, J.; Zhang, Y.; Fan, Q.; Xue, B.; Qin, M. Stretchable hydrogels with low hysteresis and anti-fatigue fracture based on polyprotein cross-linkers. Nat. Commun. 2020, 11, 4032. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Topuz, F.; Hahn, K.; Albrecht, K.; Groll, J. Embedding of Active Proteins and Living Cells in Redox-Sensitive Hydrogels and Nanogels through Enzymatic Cross-Linking. Angew. Chem. Int. Ed. 2013, 52, 3000–3003. [Google Scholar] [CrossRef] [PubMed]
- Le Thi, P.; Lee, Y.; Nguyen, D.H.; Park, K.D. In situ forming gelatin hydrogels by dual-enzymatic cross-linking for enhanced tissue adhesiveness. J. Mater. Chem. B 2017, 5, 757–764. [Google Scholar] [CrossRef]
- Chirila, T.V.; Suzuki, S.; Papolla, C. A comparative investigation of Bombyx mori silk fibroin hydrogels generated by chemical and enzymatic cross-linking. Biotechnol. Appl. Biochem. 2017, 64, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Lake, R.; Park, S.; Edwards, S.; Jones, C.; Jeong, K.J. Injectable macroporous hydrogel formed by enzymatic cross-linking of gelatin microgels. ACS Appl. Bio Mater. 2018, 1, 1430–1439. [Google Scholar] [CrossRef]
- Mezghani, K.; Spruiell, J. High speed melt spinning of poly (L-lactic acid) filaments. J. Polym. Sci. Part B Polym. Phys. 1998, 36, 1005–1012. [Google Scholar] [CrossRef]
- Matabola, K.; De Vries, A.; Moolman, F.; Luyt, A. Single polymer composites: A review. J. Mater. Sci. 2009, 44, 6213–6222. [Google Scholar] [CrossRef]
- Berry, S.M.; Harfenist, S.A.; Cohn, R.W.; Keynton, R.S. Characterization of micromanipulator-controlled dry spinning of micro-and sub-microscale polymer fibers. J. Micromechanics Microengineering 2006, 16, 1825. [Google Scholar] [CrossRef]
- Wei, W.; Zhang, Y.; Zhao, Y.; Luo, J.; Shao, H.; Hu, X. Bio-inspired capillary dry spinning of regenerated silk fibroin aqueous solution. Mater. Sci. Eng. C 2011, 31, 1602–1608. [Google Scholar] [CrossRef]
- Phillips, D.M.; Drummy, L.F.; Naik, R.R.; Hugh, C.; Fox, D.M.; Trulove, P.C.; Mantz, R.A. Regenerated silk fiber wet spinning from an ionic liquid solution. J. Mater. Chem. 2005, 15, 4206–4208. [Google Scholar] [CrossRef]
- Nelson, K.D.; Romero, A.; Waggoner, P.; Crow, B.; Borneman, A.; Smith, G.M. Technique paper for wet-spinning poly (L-lactic acid) and poly (DL-lactide-co-glycolide) monofilament fibers. Tissue Eng. 2003, 9, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.-M.; Zhang, Y.-Z.; Kotaki, M.; Ramakrishna, S. A review on polymer nanofibers by electrospinning and their applications in nanocomposites. Compos. Sci. Technol. 2003, 63, 2223–2253. [Google Scholar] [CrossRef]
- Pham, Q.P.; Sharma, U.; Mikos, A.G. Electrospinning of polymeric nanofibers for tissue engineering applications: A review. Tissue Eng. 2006, 12, 1197–1211. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Stoyanov, S.D.; Stride, E.; Pelan, E.; Edirisinghe, M. Electrospinning versus fibre production methods: From specifics to technological convergence. Chem. Soc. Rev. 2012, 41, 4708–4735. [Google Scholar] [CrossRef] [PubMed]
- Reneker, D.H.; Yarin, A.L. Electrospinning jets and polymer nanofibers. Polymer 2008, 49, 2387–2425. [Google Scholar] [CrossRef]
- Badrossamay, M.R.; McIlwee, H.A.; Goss, J.A.; Parker, K.K. Nanofiber assembly by rotary jet-spinning. Nano Lett. 2010, 10, 2257–2261. [Google Scholar] [CrossRef]
- Plowman, J.E.; Deb-Choudhury, S.; Clerens, S.; Thomas, A.; Cornellison, C.D.; Dyer, J.M. Unravelling the proteome of wool: Towards markers of wool quality traits. J. Proteom. 2012, 75, 4315–4324. [Google Scholar] [CrossRef]
- Service Robert, F. Hunt for renewable plastics clears a hurdle. Science 2021, 371, 873. [Google Scholar] [CrossRef]
- Nanda, S.; Patra, B.R.; Patel, R.; Bakos, J.; Dalai, A.K. Innovations in applications and prospects of bioplastics and biopolymers: A review. Environ. Chem. Lett. 2021, 20, 379–395. [Google Scholar] [CrossRef] [PubMed]
- Springstein, B.L.; Woehle, C.; Weissenbach, J.; Helbig, A.O.; Dagan, T.; Stucken, K. Identification and characterization of novel filament-forming proteins in cyanobacteria. Sci. Rep. 2020, 10, 1894. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.V.; Garcia Mdel, C.; Fernandez, F.G.; Bustos, C.S.; Chisti, Y.; Sevilla, J.M. Protein measurements of microalgal and cyanobacterial biomass. Bioresour. Technol. 2010, 101, 7587–7591. [Google Scholar] [CrossRef]
- Menetrez, M.Y. An overview of algae biofuel production and potential environmental impact. Environ. Sci. Technol. 2012, 46, 7073–7085. [Google Scholar] [CrossRef] [PubMed]
- Lupatini, A.L.; Colla, L.M.; Canan, C.; Colla, E. Potential application of microalga Spirulina platensis as a protein source. J. Sci. Food Agric. 2017, 97, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Betterle, N.; Hidalgo Martinez, D.; Melis, A. Cyanobacterial Production of Biopharmaceutical and Biotherapeutic Proteins. Front. Plant Sci. 2020, 11, 237. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhang, H.; Meng, H.; Zhu, Y.; Bao, G.; Zhang, Y.; Li, Y.; Ma, Y. Discovery of a super-strong promoter enables efficient production of heterologous proteins in cyanobacteria. Sci. Rep. 2014, 4, 4500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Betterle, N.; Hidalgo Martinez, D.; Melis, A. Recombinant Protein Stability in Cyanobacteria. ACS Synth. Biol. 2021, 10, 810–825. [Google Scholar] [CrossRef] [PubMed]
- Kouba, P.; Kohout, P.; Haddadi, F.; Bushuiev, A.; Samusevich, R.; Sedlar, J.; Damborsky, J.; Pluskal, T.; Sivic, J.; Mazurenko, S. Machine Learning-Guided Protein Engineering. ACS Catal. 2023, 13, 13863–13895. [Google Scholar] [CrossRef]
- Singh, N.; Malik, S.; Gupta, A.; Srivastava, K.R. Revolutionizing enzyme engineering through artificial intelligence and machine learning. Emerg. Top. Life Sci. 2021, 5, 113–125. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).