

Metachronal Motion of Biological and Artificial Cilia



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
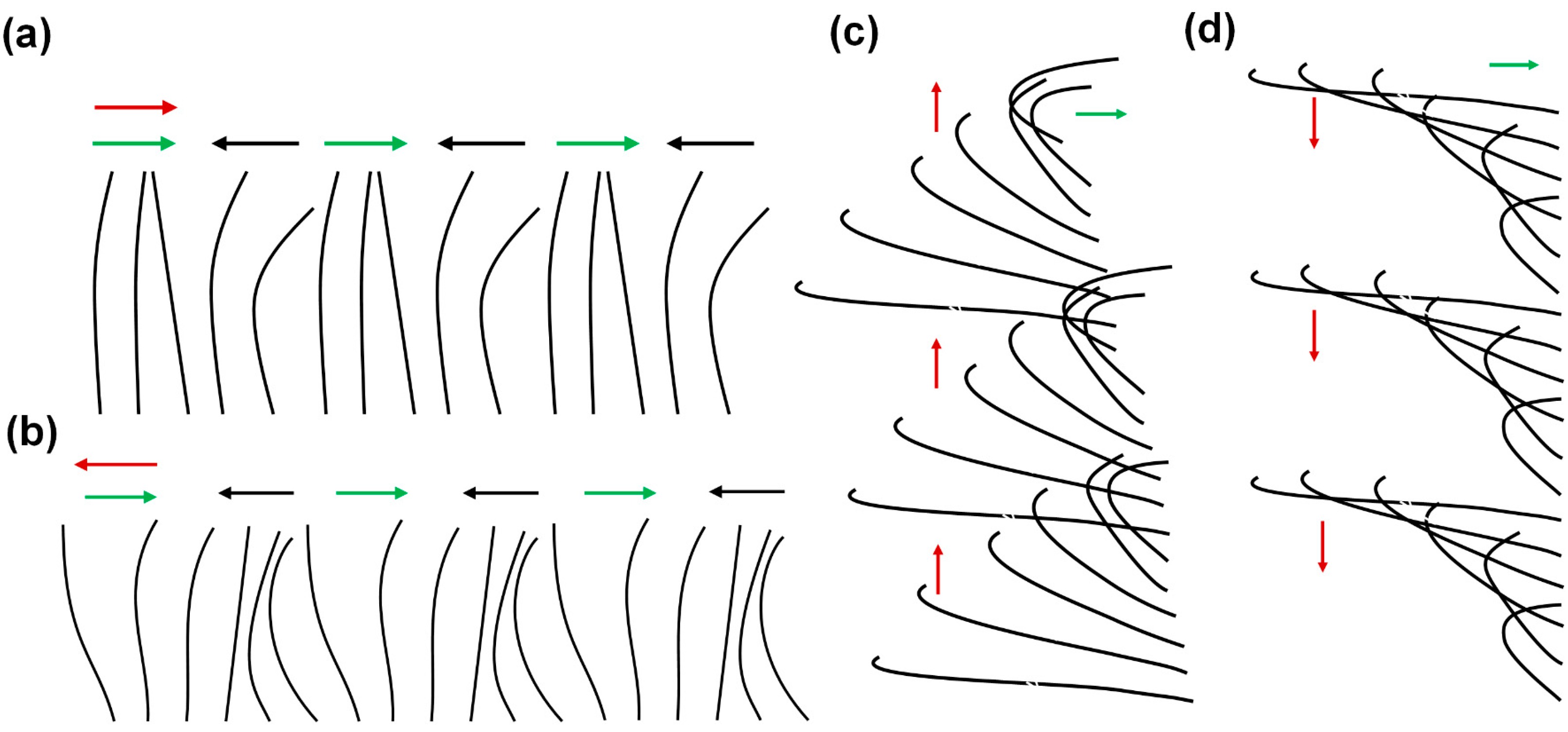
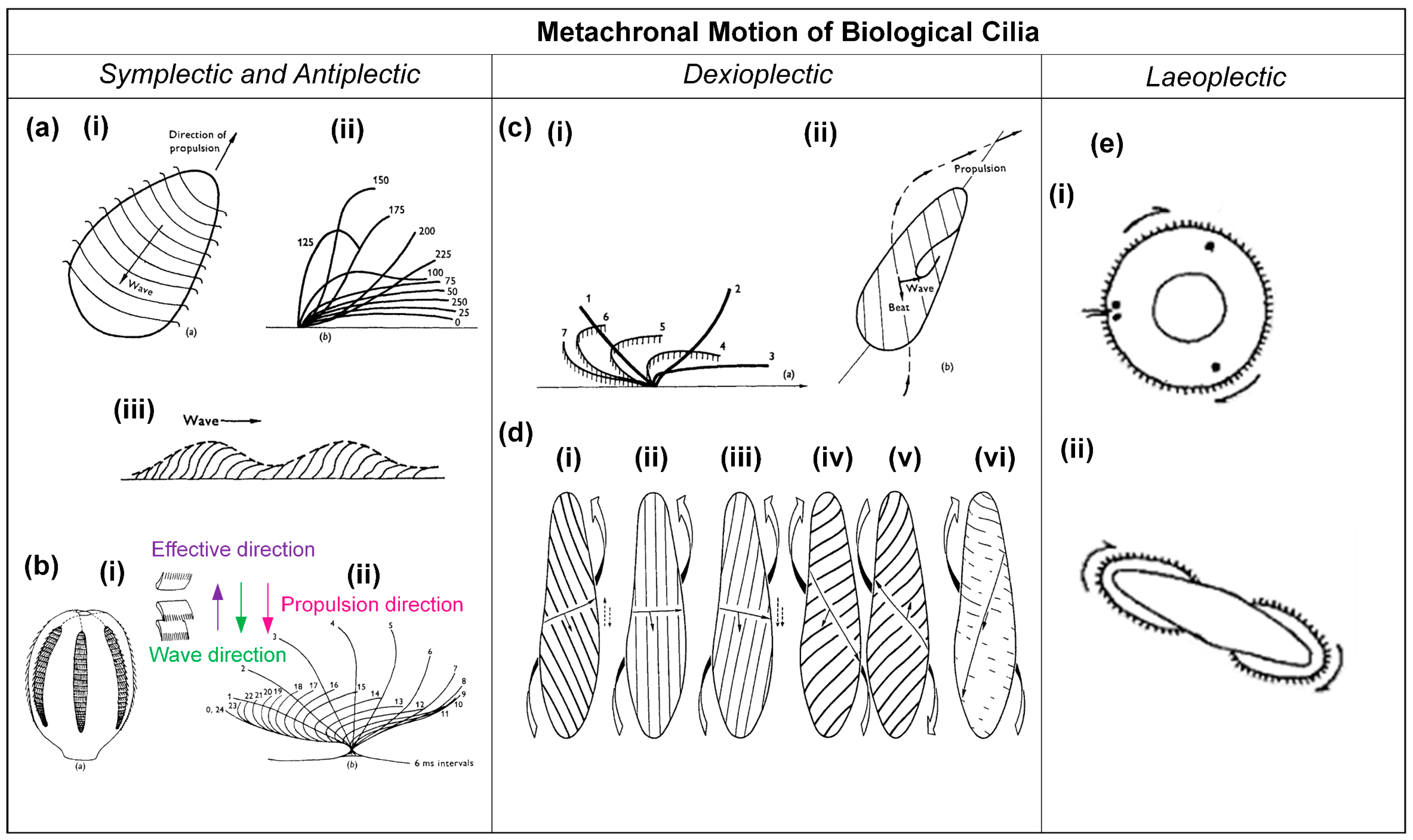
2. Metachronal Motion of Biological Cilia
3. Metachronal Motion of Artificial Cilia
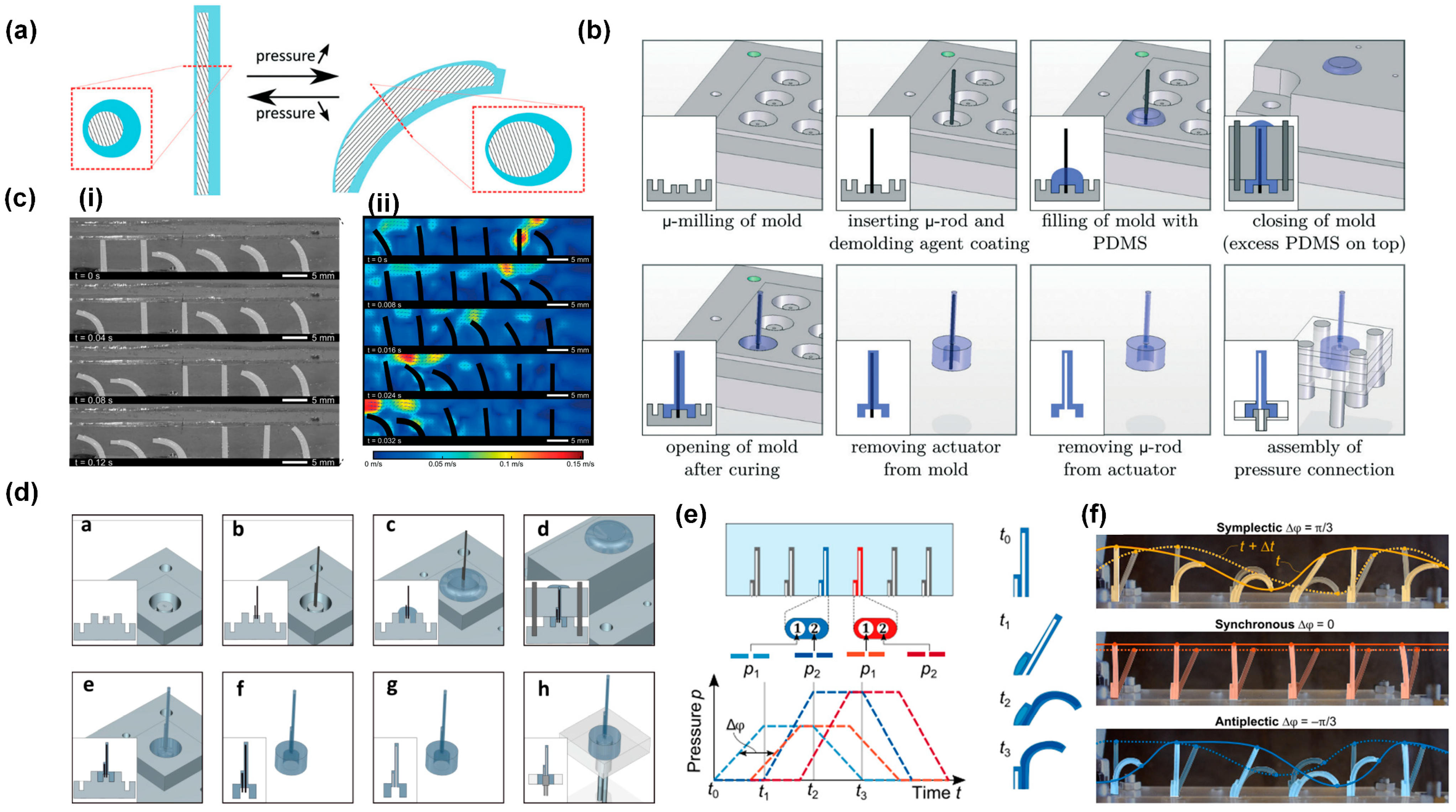
3.1. Metachronal Motion of Pneumatically Driven Artificial Cilia
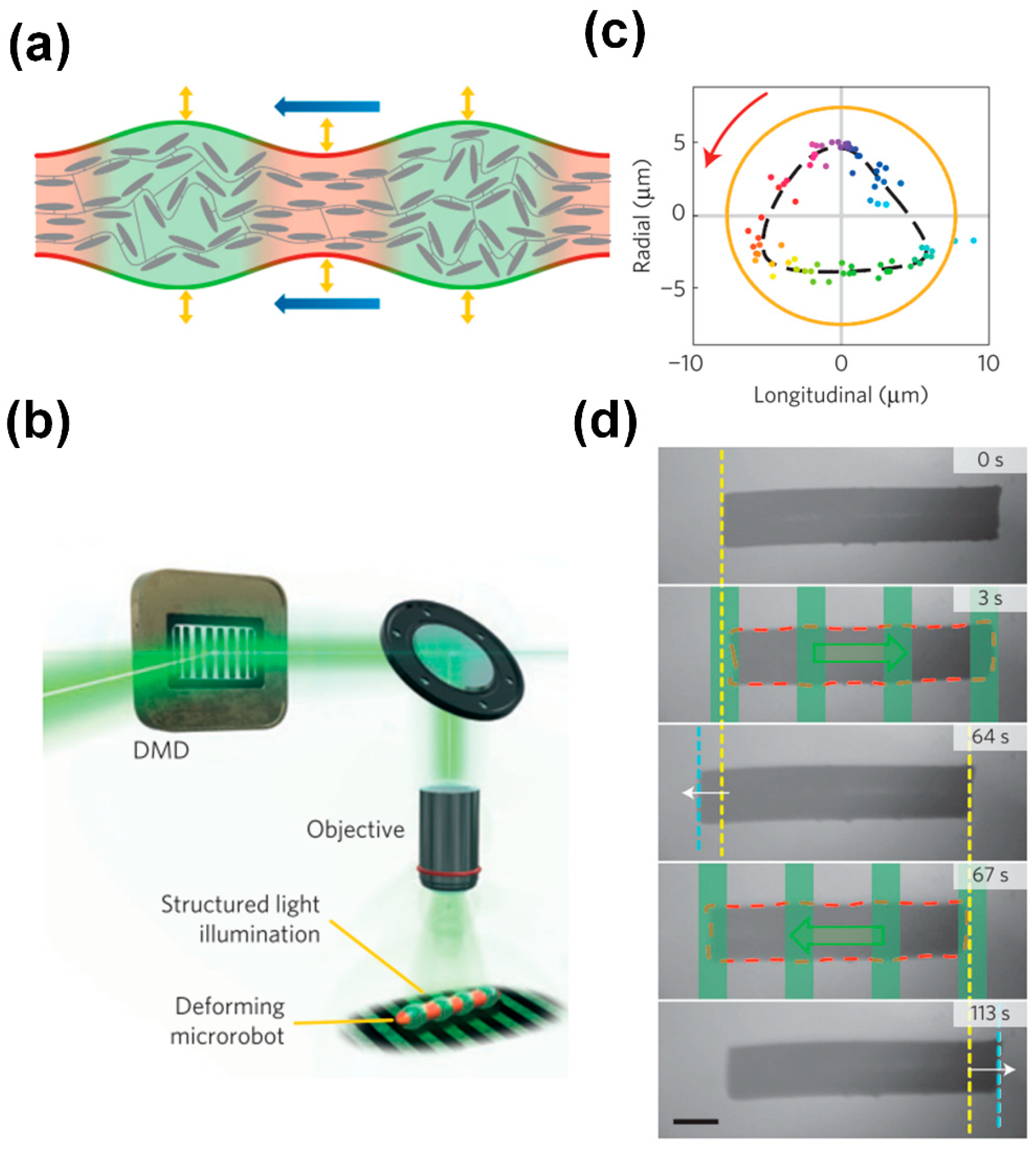
3.2. Metachronal Motion of Light-Driven Artificial Cilia
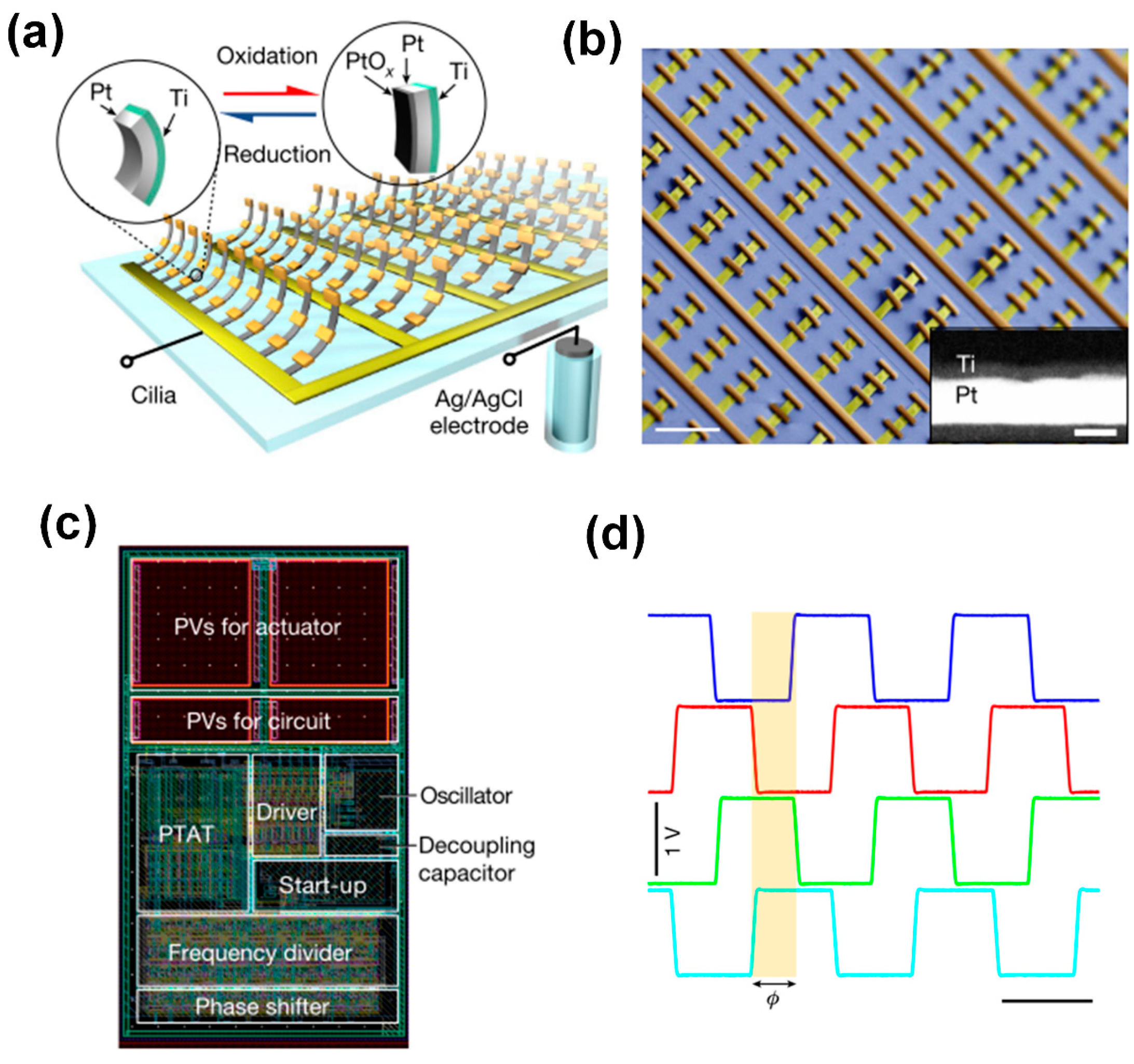
3.3. Metachronal Motion of Electrically Driven Artificial Cilia
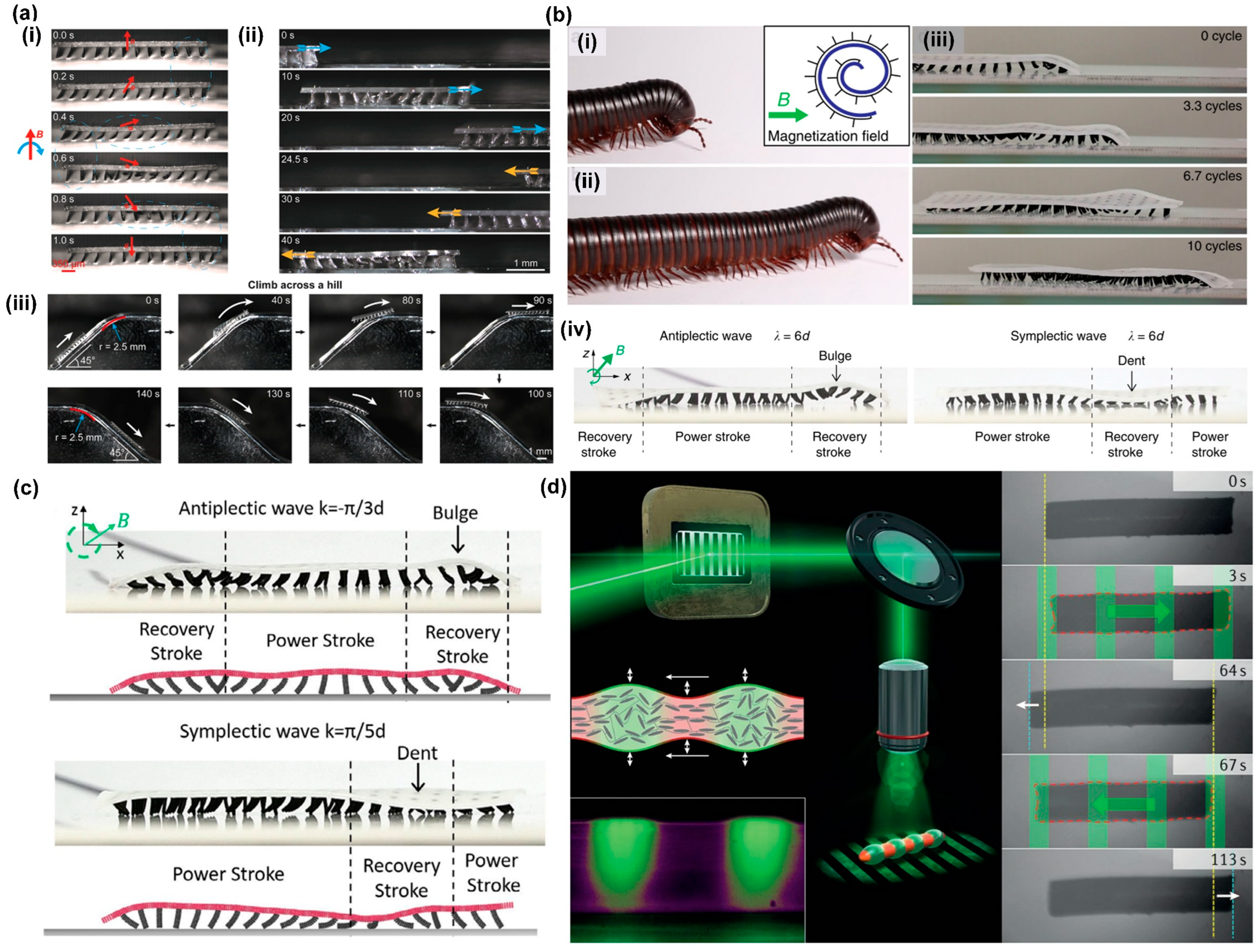
3.4. Metachronal Motion of Magnetically Driven Artificial Cilia
- Controlling the magnetic particle distribution within the artificial cilia.
- Controlling the geometry of the artificial cilia.
- Controlling the magnetic field.
4. Applications of Metachronal Motion in Artificial Cilia
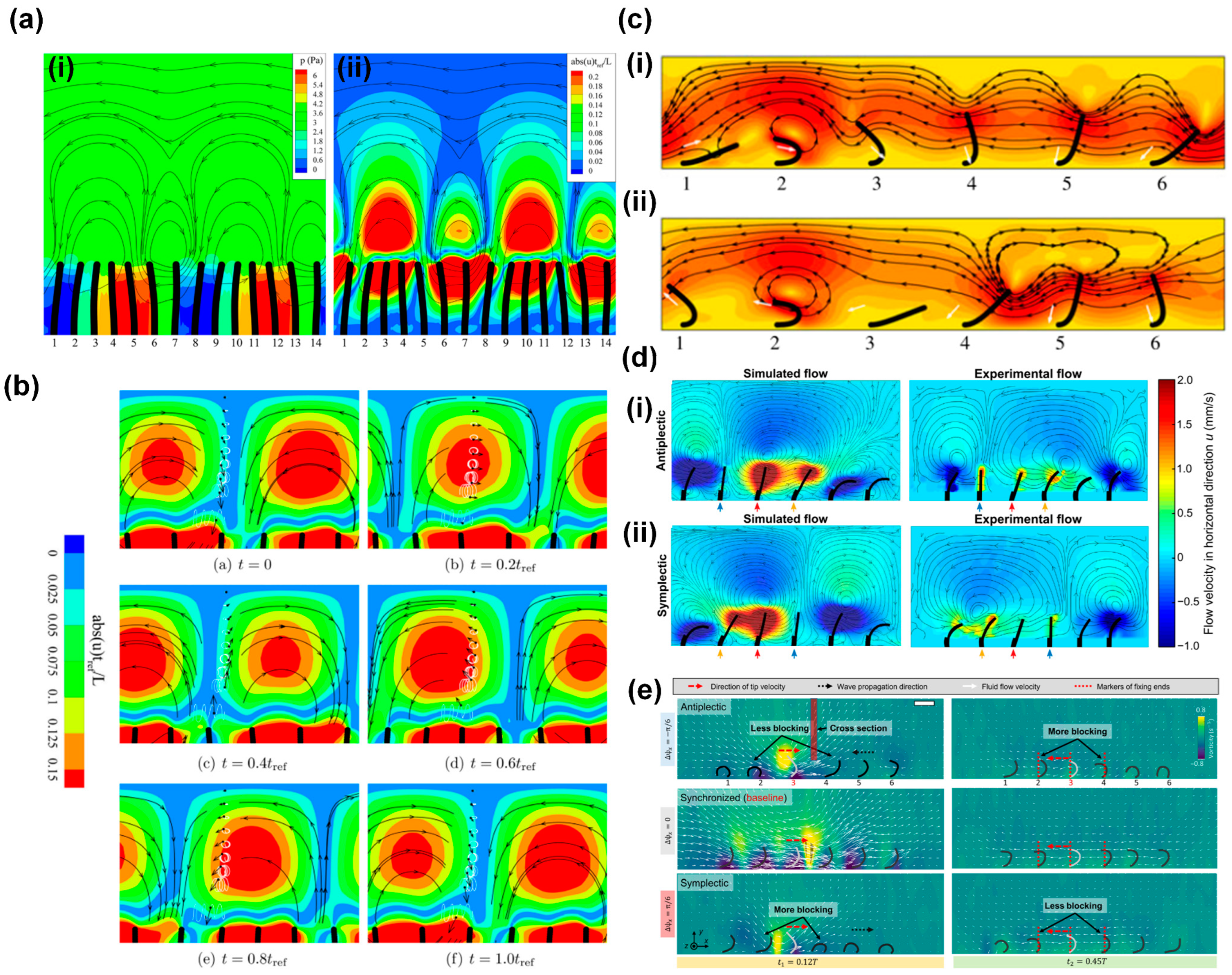
4.1. Flow Generation
4.2. Transportation
- Mucus transportation.
- Particle transportation.
- Droplet transportation.
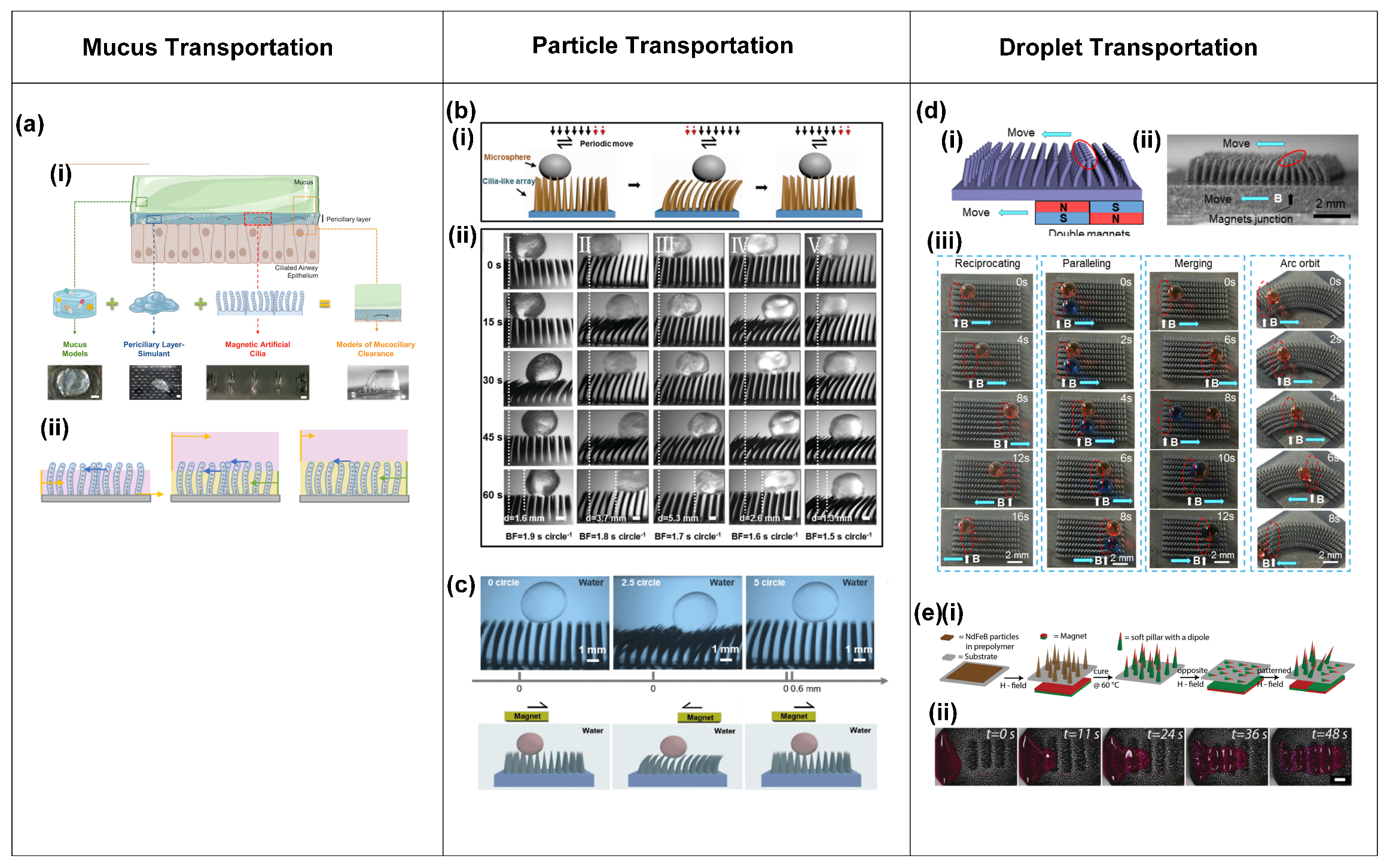
- Microrobot locomotion.
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bustamante-Marin, X.M.; Ostrowski, L.E. Cilia and Mucociliary Clearance. Cold Spring Harb. Perspect. Biol. 2017, 9, a02824. [Google Scholar] [CrossRef]
- Sanderson, M.J.; Sleigh, M.A. Ciliary Activity of Cultured Rabbit Tracheal Epithelium: Beat Pattern and Metachrony. J. Cell Sci. 1981, 47, 331–347. [Google Scholar] [CrossRef]
- ul Islam, T.; Wang, Y.; Aggarwal, I.; Cui, Z.; Eslami Amirabadi, H.; Garg, H.; Kooi, R.; Venkataramanachar, B.B.; Wang, T.; Zhang, S.; et al. Microscopic Artificial Cilia—A Review. Lab Chip 2022, 22, 1650–1679. [Google Scholar] [CrossRef]
- Machemer, H. Ciliary Activity and the Origin of Metachrony in Paramecium: Effects of Increased Viscosity. J. Exp. Biol. 1972, 57, 239–259. [Google Scholar] [CrossRef]
- Blake, J.R.; Sleigh, M.A. Mechanics of Ciliary Locomotion. Biol. Rev. Camb. Philos. Soc. 1974, 49, 85–125. [Google Scholar] [CrossRef]
- Mellor, J.S.; Hyams, J.S. Metachronism of Cilia of the Digestive Epithelium of Cirriformia Tentaculata. Micron 1978, 9, 91–94. [Google Scholar] [CrossRef]
- Satir, P. The Fixation of the Metaehronal Wave. J. Cell Biol. 1963, 18, 345–365. [Google Scholar] [CrossRef]
- Knight-Jones, E.W. Relations between Metachronism and the Direction of Ciliary Beat in Metazoa. J. Cell Sci. 1954, s3–s95, 503–521. [Google Scholar] [CrossRef]
- Fulford, G.R.; Blake, J.R. Muco-Ciliary Transport in the Lung. J. Theor. Biol. 1986, 121, 381–402. [Google Scholar] [CrossRef]
- Gueron, S.; Levit-Gurevich, K. Energetic Considerations of Ciliary Beating and the Advantage of Metachronal Coordination. Proc. Natl. Acad. Sci. USA 1999, 96, 12240–12245. [Google Scholar] [CrossRef]
- Guirao, B.; Joanny, J.F. Spontaneous Creation of Macroscopic Flow and Metachronal Waves in an Array of Cilia. Biophys. J. 2007, 92, 1900–1917. [Google Scholar] [CrossRef]
- Blake, J.R. Mechanics of Muco-Ciliary Transport. IMA J. Appl. Math. 1984, 32, 69–87. [Google Scholar] [CrossRef]
- Mitran, S.M. Metachronal Wave Formation in a Model of Pulmonary Cilia. Comput. Struct. 2007, 85, 763–774. [Google Scholar] [CrossRef]
- Norton, M.M.; Robinson, R.J.; Weinstein, S.J. Model of Ciliary Clearance and the Role of Mucus Rheology. Phys. Rev. E-Stat. Nonlinear Soft Matter. Phys. 2011, 83, 011921. [Google Scholar] [CrossRef]
- Chateau, S.; Favier, J.; Poncet, S.; D’Ortona, U. Why Antiplectic Metachronal Cilia Waves Are Optimal to Transport Bronchial Mucus. Phys. Rev. E 2019, 100, 042405. [Google Scholar] [CrossRef]
- Khaderi, S.N.; Craus, C.B.; Hussong, J.; Schorr, N.; Belardi, J.; Westerweel, J.; Prucker, O.; Rühe, J.; Den Toonder, J.M.J.; Onck, P.R. Magnetically-Actuated Artificial Cilia for Microfluidic Propulsion. Lab Chip 2011, 11, 2002–2010. [Google Scholar] [CrossRef]
- Elgeti, J.; Gompper, G. Emergence of Metachronal Waves in Cilia Arrays. Proc. Natl. Acad. Sci. USA 2013, 110, 4470–4475. [Google Scholar] [CrossRef]
- Cui, Z.; Wang, Y.; Zhang, S.; Wang, T.; den Toonder, J.M. Miniaturized Metachronal Magnetic Artificial Cilia. Proc. Natl. Acad. Sci. USA 2023, 120, e2304519120. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, J.; Fu, X.; Zhang, D.; Zhao, Y. Tailoring Flexible Arrays for Artificial Cilia Actuators. Adv. Intell. Syst. 2021, 3, 2000225. [Google Scholar] [CrossRef]
- Burn, A.; Schneiter, M.; Ryser, M.; Gehr, P.; Rička, J.; Frenz, M. A Quantitative Interspecies Comparison of the Respiratory Mucociliary Clearance Mechanism. Eur. Biophys. J. 2022, 51, 51–65. [Google Scholar] [CrossRef]
- Milana, E.; Zhang, R.; Vetrano, M.R.; Peerlinck, S.; de Volder, M.; Onck, P.R.; Reynaerts, D.; Gorissen, B. Metachronal Patterns in Artificial Cilia for Low Reynolds Number Fluid Propulsion. Sci. Adv. 2020, 6, eabd2508. [Google Scholar] [CrossRef]
- Toonder, J.D.; Bos, F.; Broer, D.; Filippini, L.; Gillies, M.; De Goede, J.; Mol, T.; Reijme, M.; Talen, W.; Wilderbeek, H.; et al. Artificial Cilia for Active Micro-Fluidic Mixing. Lab Chip 2008, 8, 533–541. [Google Scholar] [CrossRef]
- Wang, W.; Liu, Q.; Tanasijevic, I.; Reynolds, M.F.; Cortese, A.J.; Miskin, M.Z.; Cao, M.C.; Muller, D.A.; Molnar, A.C.; Lauga, E. Cilia Metasurfaces for Electronically Programmable Microfluidic Manipulation. Nature 2022, 605, 681–686. [Google Scholar] [CrossRef]
- Dai, B.; Li, S.; Xu, T.; Wang, Y.; Zhang, F.; Gu, Z.; Wang, S. Arti Fi Cial Asymmetric Cilia Array of Dielectric Elastomer for Cargo Transportation. ACS Appl. Mater. Interfaces 2018, 10, 6–11. [Google Scholar] [CrossRef]
- Demirörs, A.F.; Aykut, S.; Ganzeboom, S.; Meier, Y.A.; Hardeman, R.; de Graaf, J.; Mathijssen, A.J.T.M.T.M.; Poloni, E.; Carpenter, J.A.; Ünlü, C.; et al. Amphibious Transport of Fluids and Solids by Soft Magnetic Carpets. Adv. Sci. 2021, 8, 2102510. [Google Scholar] [CrossRef]
- Gorissen, B.; De Volder, M.; Reynaerts, D. Pneumatically-Actuated Artificial Cilia Array for Biomimetic Fluid Propulsion. Lab Chip 2015, 15, 4348–4355. [Google Scholar] [CrossRef]
- Zhang, R.; den Toonder, J.; Onck, P.R. Transport and Mixing by Metachronal Waves in Nonreciprocal Soft Robotic Pneumatic Artificial Cilia at Low Reynolds Numbers. Phys. Fluids 2021, 33, 092009. [Google Scholar] [CrossRef]
- Van Oosten, C.L.; Bastiaansen, C.W.M.; Broer, D.J. Printed Artificial Cilia from Liquid-Crystal Network Actuators Modularly Driven by Light. Nat. Mater. 2009, 8, 677–682. [Google Scholar] [CrossRef]
- Palagi, S.; Mark, A.G.; Reigh, S.Y.; Melde, K.; Qiu, T.; Zeng, H.; Parmeggiani, C.; Martella, D.; Sanchez-Castillo, A.; Kapernaum, N.; et al. Structured Light Enables Biomimetic Swimming and Versatile Locomotion of Photoresponsive Soft Microrobots. Nat. Mater. 2016, 15, 647–653. [Google Scholar] [CrossRef]
- Li, S.; Lerch, M.M.; Waters, J.T.; Deng, B.; Martens, R.S.; Yao, Y.; Kim, D.Y.; Bertoldi, K.; Grinthal, A.; Balazs, A.C.; et al. Self-Regulated Non-Reciprocal Motions in Single-Material Microstructures. Nature 2022, 605, 76–83. [Google Scholar] [CrossRef]
- Miao, J.; Zhang, T.; Li, G.; Shang, W.; Shen, Y. Magnetic Artificial Cilia Carpets for Transport, Mixing, and Directional Diffusion. Adv. Eng. Mater. 2021, 24, 2101399. [Google Scholar] [CrossRef]
- Chen, G.; Dai, Z.; Li, S.; Huang, Y.; Xu, Y.; She, J.; Zhou, B. Magnetically Responsive Film Decorated with Microcilia for Robust and Controllable Manipulation of Droplets. ACS Appl. Mater. Interfaces 2021, 13, 1754–1765. [Google Scholar] [CrossRef]
- Lin, Y.; Hu, Z.; Zhang, M.; Xu, T.; Feng, S.; Jiang, L.; Zheng, Y. Magnetically Induced Low Adhesive Direction of Nano/Micropillar Arrays for Microdroplet Transport. Adv. Funct. Mater. 2018, 28, 1800163. [Google Scholar] [CrossRef]
- Zhou, Y.; Huang, S.; Tian, X. Magnetoresponsive Surfaces for Manipulation of Nonmagnetic Liquids: Design and Applications. Adv. Funct. Mater. 2020, 30, 1906507. [Google Scholar] [CrossRef]
- Dong, X.; Lum, G.Z.; Hu, W.; Zhang, R.; Ren, Z.; Onck, P.R.; Sitti, M. Bioinspired Cilia Arrays with Programmable Nonreciprocal Motion and Metachronal Coordination. Sci. Adv. 2020, 6, eabc9323. [Google Scholar] [CrossRef]
- Hanasoge, S.; Hesketh, P.J.; Alexeev, A. Metachronal Motion of Artificial Magnetic Cilia. Soft Matter. 2018, 14, 3689–3693. [Google Scholar] [CrossRef]
- Zhang, R.; den Toonder, J.; Onck, P.R. Metachronal Patterns by Magnetically-Programmable Artificial Cilia Surfaces for Low Reynolds Number Fluid Transport and Mixing. Soft Matter. 2022, 18, 3902–3909. [Google Scholar] [CrossRef]
- Zhang, S.; Cui, Z.; Wang, Y.; Den Toonder, J.M.J.; Toonder, J. den Metachronal Actuation of Microscopic Magnetic Artificial Cilia Generates Strong Microfluidic Pumping. Lab Chip 2020, 20, 3569–3581. [Google Scholar] [CrossRef]
- Choksi, S.P.; Lauter, G.; Swoboda, P.; Roy, S. Switching on Cilia: Transcriptional Networks Regulating Ciliogenesis. Development 2014, 141, 1427–1441. [Google Scholar] [CrossRef]
- Gilpin, W.; Bull, M.S.; Prakash, M. The Multiscale Physics of Cilia and Flagella. Nat. Rev. Phys. 2020, 2, 74–88. [Google Scholar] [CrossRef]
- Elgeti, J.; Winkler, R.G.; Gompper, G. Physics of Microswimmers—Single Particle Motion and Collective Behavior: A Review. Rep. Prog. Phys. 2015, 78, 056601. [Google Scholar] [CrossRef]
- Wang, C.; Tang, H.; Zhang, X. Fluid-Structure Interaction of Bio-Inspired Flexible Slender Structures: A Review of Selected Topics. Bioinspir. Biomim. 2022, 17, 041002. [Google Scholar] [CrossRef]
- Pazour, G.J.; Quarmby, L.; Smith, A.O.; Desai, P.B.; Schmidts, M. Cilia in Cystic Kidney and Other Diseases. Cell Signal. 2019, 69, 109519. [Google Scholar] [CrossRef]
- Hirokawa, N.; Okada, Y.; Tanaka, Y. Fluid Dynamic Mechanism Responsible for Breaking the Left-Right Symmetry of the Human Body: The Nodal Flow. Annu. Rev. Fluid Mech. 2009, 41, 53–72. [Google Scholar] [CrossRef]
- Ul Islam, T.; Bellouard, Y.; Den Toonder, J.M.J. Highly Motile Nanoscale Magnetic Artificial Cilia. Proc. Natl. Acad. Sci. USA 2021, 118, e2104930118. [Google Scholar] [CrossRef]
- Vanaki, S.M.; Holmes, D.; Saha, S.C.; Chen, J.; Brown, R.J.; Jayathilake, P.G. Muco-Ciliary Clearance: A Review of Modelling Techniques. J. Biomech. 2020, 99, 109578. [Google Scholar] [CrossRef]
- Odor, D.L.; Blandau, R.J. Observations on the Solitary Cilium of Rabbit Oviductal Epithelium: Its Motility and Ultrastructure. Am. J. Anat. 1985, 174, 437–453. [Google Scholar] [CrossRef]
- Osterman, N.; Vilfan, A. Finding the Ciliary Beating Pattern with Optimal Efficiency. Proc. Natl. Acad. Sci. USA 2011, 108, 15727–15732. [Google Scholar] [CrossRef]
- Chateau, S.; Favier, J.; D’Ortona, U.; Poncet, S. Transport Efficiency of Metachronal Waves in 3D Cilium Arrays Immersed in a Two-Phase Flow. J. Fluid Mech. 2017, 824, 931–961. [Google Scholar] [CrossRef]
- Hanasoge, S.; Hesketh, P.J.; Alexeev, A. Metachronal Actuation of Microscale Magnetic Artificial Cilia. ACS Appl. Mater. Interfaces 2020, 12, 46963–46971. [Google Scholar] [CrossRef]
- Lyons, R.; Saridogan, E.; Djahanbakhch, O. The reproductive significance of human Fallopian tube cilia. Hum. Reprod. Update 2006, 12, 363–372. [Google Scholar] [CrossRef]
- Sitti, M.; Wiersma, D.S. Pros and Cons: Magnetic versus Optical Microrobots. Adv. Mater. 2020, 32, 1906766. [Google Scholar] [CrossRef]
- Liu, Z.; Li, M.; Dong, X.; Ren, Z.; Hu, W.; Sitti, M. Creating Three-Dimensional Magnetic Functional Microdevices via Molding-Integrated Direct Laser Writing. Nat. Commun. 2022, 13, 2016. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, K.; Liang, D.; Yan, L.; Ni, K.; Huang, H.; Li, B.; Guo, Z.; Wang, J.; Ma, X.; et al. Hybrid Magnetic Micropillar Arrays for Programmable Actuation. Adv. Mater. 2020, 32, 2001879. [Google Scholar] [CrossRef]
- Kim, J.; Chung, S.E.; Choi, S.E.; Lee, H.; Kim, J.; Kwon, S. Programming Magnetic Anisotropy in Polymeric Microactuators. Nat. Mater. 2011, 10, 747–752. [Google Scholar] [CrossRef]
- Zarzar, L.; Kim, P.; Aizenberg, J. Bio-inspired design of submerged hydrogel-actuated polymer microstructures operating in response to pH. Adv. Mater. 2011, 23, 1442–1446. [Google Scholar] [CrossRef]
- Zhang, S.; Cui, Z.; Wang, Y.; Den Toonder, J. Metachronal μ-Cilia for On-Chip Integrated Pumps and Climbing Robots. ACS Appl. Mater. Interfaces 2021, 13, 20845–20857. [Google Scholar] [CrossRef]
- Gu, H.; Boehler, Q.; Cui, H.; Secchi, E.; Savorana, G.; De Marco, C.; Gervasoni, S.; Peyron, Q.; Huang, T.Y.; Pane, S.; et al. Magnetic Cilia Carpets with Programmable Metachronal Waves. Nat. Commun. 2020, 11, 2637. [Google Scholar] [CrossRef]
- Tsumori, F.; Marume, R.; Saijou, A.; Kudo, K.; Osada, T.; Miura, H. Metachronal Wave of Artificial Cilia Array Actuated by Applied Magnetic Field. JPN J. Appl. Phys. 2016, 55, 06GP19. [Google Scholar] [CrossRef]
- Milana, E.; Gorissen, B.; Peerlinck, S.; De Volder, M.; Reynaerts, D. Artificial Soft Cilia with Asymmetric Beating Patterns for Biomimetic Low-Reynolds-Number Fluid Propulsion. Adv. Funct. Mater. 2019, 29, 1900462. [Google Scholar] [CrossRef]
- Hussong, J.; Breugem, W.P.; Westerweel, J. A Continuum Model for Flow Induced by Metachronal Coordination between Beating Cilia. J. Fluid Mech. 2011, 684, 137–162. [Google Scholar] [CrossRef]
- Hall, J.; Clarke, N. The Mechanics of Cilium Beating: Quantifying the Relationship between Metachronal Wavelength and Fluid Flow Rate. J. Fluid Mech. 2020, 891, A20. [Google Scholar] [CrossRef]
- Khaderi, S.N.; Den Toonder, J.M.J.; Onck, P.R. Microfluidic Propulsion by the Metachronal Beating of Magnetic Artificial Cilia: A Numerical Analysis. J. Fluid Mech. 2011, 688, 44–65. [Google Scholar] [CrossRef]
- Khaderi, S.N.; den Toonder, J.M.J.; Onck, P.R. Fluid Flow Due to Collective Non-Reciprocal Motion of Symmetrically-Beating Artificial Cilia. Biomicrofluidics 2012, 6, 014106. [Google Scholar] [CrossRef]
- Mesdjian, O.; Wang, C.; Gsell, S.; D’Ortona, U.; Favier, J.; Viallat, A.; Loiseau, E. Longitudinal to Transverse Metachronal Wave Transitions in an in Vitro Model of Ciliated Bronchial Epithelium. Phys. Rev. Lett. 2022, 129, 38101. [Google Scholar] [CrossRef]
- Pedersoli, L.; Zhang, S.; Briatico-Vangosa, F.; Petrini, P.; Cardinaels, R.; den Toonder, J.; Peneda Pacheco, D. Engineered Modular Microphysiological Models of the Human Airway Clearance Phenomena. Biotechnol. Bioeng. 2021, 118, 3898–3913. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, Y.; Onck, P.; den Toonder, J. A Concise Review of Microfluidic Particle Manipulation Methods. Microfluid. Nanofluidics 2020, 24, 24. [Google Scholar] [CrossRef]
- Ben, S.; Tai, J.; Ma, H.; Peng, Y.; Zhang, Y.; Tian, D.; Liu, K.; Jiang, L. Cilia-Inspired Flexible Arrays for Intelligent Transport of Viscoelastic Microspheres. Adv. Funct. Mater. 2018, 28, 1706666. [Google Scholar] [CrossRef]
- Ben, S.; Yao, J.; Ning, Y.; Zhao, Z.; Zha, J.; Tian, D.; Liu, K.; Jiang, L. A Bioinspired Magnetic Responsive Cilia Array Surface for Microspheres Underwater Directional Transport. Sci. China Chem. 2020, 63, 347–353. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, S.M.; Lee, B.J.; Ko, H.; Bae, W. Remote Manipulation of Droplets on a Flexible Magnetically Responsive Film. Nat. Publ. Gr. 2015, 5, 17843. [Google Scholar] [CrossRef]
- Ben, S.; Zhou, T.; Ma, H.; Yao, J.; Ning, Y.; Tian, D.; Liu, K.; Jiang, L. Multifunctional Magnetocontrollable Superwettable-Microcilia Surface for Directional Droplet Manipulation. Adv. Sci. 2019, 6, 1900834. [Google Scholar] [CrossRef]
- Song, Y.; Jiang, S.; Li, G.; Zhang, Y.; Wu, H.; Xue, C.; You, H.; Zhang, D.; Cai, Y.; Zhu, J.; et al. Cross-Species Bioinspired Anisotropic Surfaces for Active Droplet Transportation Driven by Unidirectional Microcolumn Waves. ACS Appl. Mater. Interfaces 2020, 12, 42264–42273. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Z.; Wang, Z.; Liang, Y.; Cui, Z.; Zhao, J.; Li, X.; Ren, L. Multistimuli-Responsive Microstructured Superamphiphobic Surfaces with Large-Range, Reversible Switchable Wettability for Oil. ACS Appl. Mater. Interfaces 2019, 11, 28478–28486. [Google Scholar] [CrossRef]
- Jiang, S.; Hu, Y.; Wu, H.; Li, R.; Zhang, Y.; Chen, C.; Xue, C.; Xu, B.; Zhu, W.; Li, J.; et al. Three-Dimensional Multifunctional Magnetically Responsive Liquid Manipulator Fabricated by Femtosecond Laser Writing and Soft Transfer. Nano Lett. 2020, 20, 7519–7529. [Google Scholar] [CrossRef]
- Zeng, H.; Wasylczyk, P.; Parmeggiani, C.; Martella, D.; Burresi, M.; Wiersma, D.S. Light-Fueled Microscopic Walkers. Adv. Mater. 2015, 27, 3883–3887. [Google Scholar] [CrossRef]
- Zeng, H.; Wani, O.M.; Wasylczyk, P.; Priimagi, A. Light-Driven, Caterpillar-Inspired Miniature Inching Robot. Macromol. Rapid Commun. 2018, 39, 1700224. [Google Scholar] [CrossRef]
- Zheng, Q.; Wu, Z.L. Light-Steered Locomotion of Muscle-like Hydrogel by Self-Coordinated Shape Change and Friction Modulation. Nat. Commun. 2020, 11, 5166. [Google Scholar] [CrossRef]
- Joyee, E.B.; Pan, Y. A Fully Three-Dimensional Printed Inchworm-Inspired Soft Robot with Magnetic Actuation. Soft Robot. 2019, 6, 333–345. [Google Scholar] [CrossRef]
- Alapan, Y.; Karacakol, A.C.; Guzelhan, S.N.; Isik, I.; Sitti, M. Reprogrammable Shape Morphing of Magnetic Soft Machines. Sci. Adv. 2020, 6, eabc6414. [Google Scholar] [CrossRef]
- Venkiteswaran, V.K.; Samaniego, L.F.P.; Sikorski, J.; Misra, S. Bio-Inspired Terrestrial Motion of Magnetic Soft Millirobots. IEEE Robot. Autom. Lett. 2019, 4, 1753–1759. [Google Scholar] [CrossRef]
- Xu, T.; Zhang, J.; Salehizadeh, M.; Onaizah, O.; Diller, E. Millimeter-Scale Flexible Robots with Programmable Three-Dimensional Magnetization and Motions. Sci. Robot. 2019, 4, eaav4494. [Google Scholar] [CrossRef]
- Hu, W.; Lum, G.Z.; Mastrangeli, M.; Sitti, M. Small-Scale Soft-Bodied Robot with Multimodal Locomotion. Nature 2018, 554, 81–85. [Google Scholar] [CrossRef]
- Palagi, S.; Fischer, P. Bioinspired Microrobots. Nat. Rev. Mater. 2018, 3, 113–124. [Google Scholar] [CrossRef]
- Jiang, H.; Gu, H.; Nelson, B.J.; Zhang, T. Numerical Study of Metachronal Wave-Modulated Locomotion in Magnetic Cilia Carpets. Adv. Intell. Syst. 2023, 5, 2300212. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, Z.; Wang, Y.; den Toonder, J.M.J. Metachronal Motion of Biological and Artificial Cilia. Biomimetics 2024, 9, 198. https://doi.org/10.3390/biomimetics9040198
Cui Z, Wang Y, den Toonder JMJ. Metachronal Motion of Biological and Artificial Cilia. Biomimetics. 2024; 9(4):198. https://doi.org/10.3390/biomimetics9040198
Chicago/Turabian StyleCui, Zhiwei, Ye Wang, and Jaap M. J. den Toonder. 2024. "Metachronal Motion of Biological and Artificial Cilia" Biomimetics 9, no. 4: 198. https://doi.org/10.3390/biomimetics9040198
APA StyleCui, Z., Wang, Y., & den Toonder, J. M. J. (2024). Metachronal Motion of Biological and Artificial Cilia. Biomimetics, 9(4), 198. https://doi.org/10.3390/biomimetics9040198