
Sporopollenin Capsules as Biomimetic Templates for the Synthesis of Hydroxyapatite and β-TCP

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Purification of Exine Capsules from Raw Pollen Grains
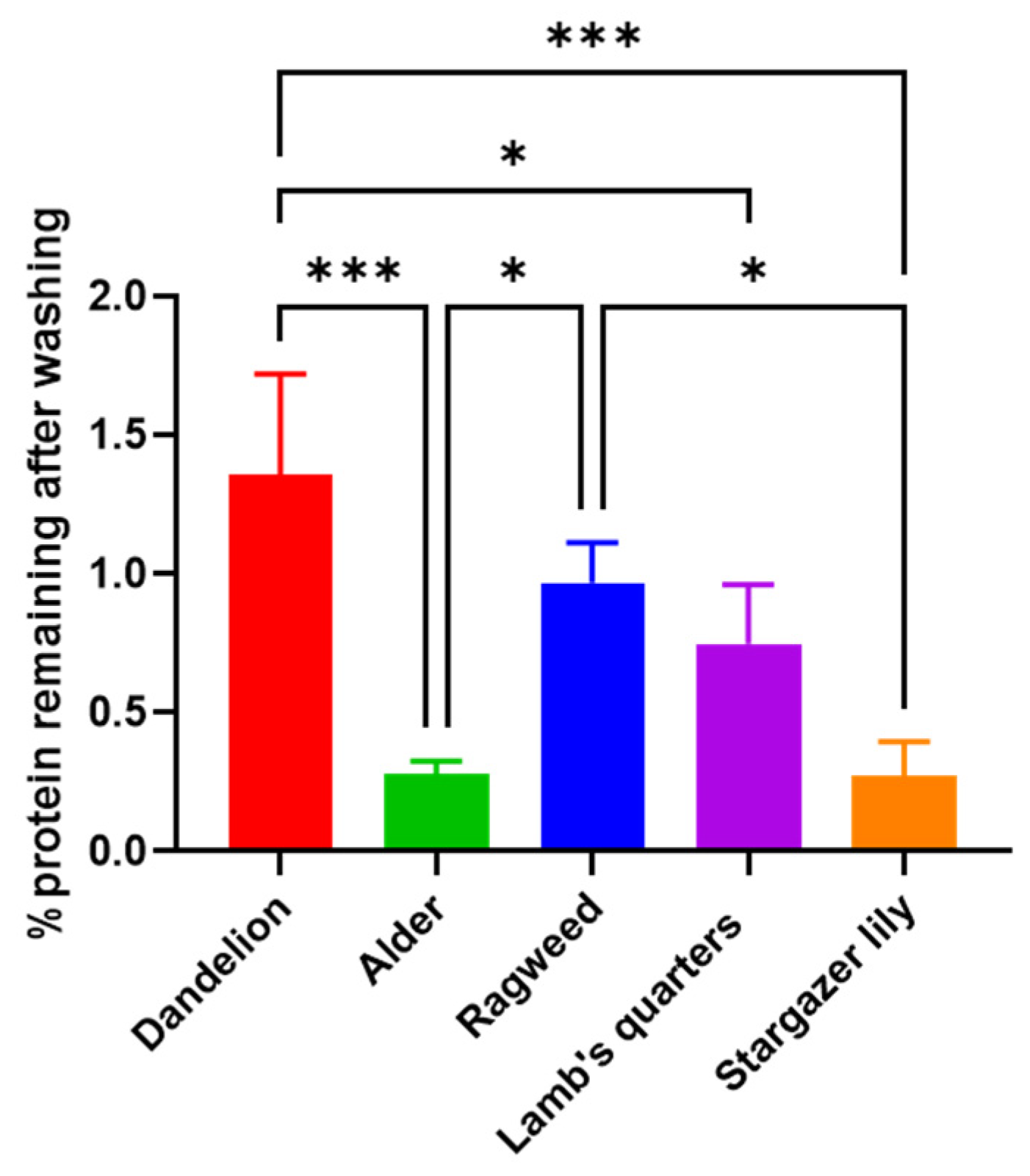
2.3. Bicinchoninic Acid (BCA) Protein Assay
2.4. Synthesis of Hydroxyapatite on Bio-Templates
2.5. Physicochemical Characterisation
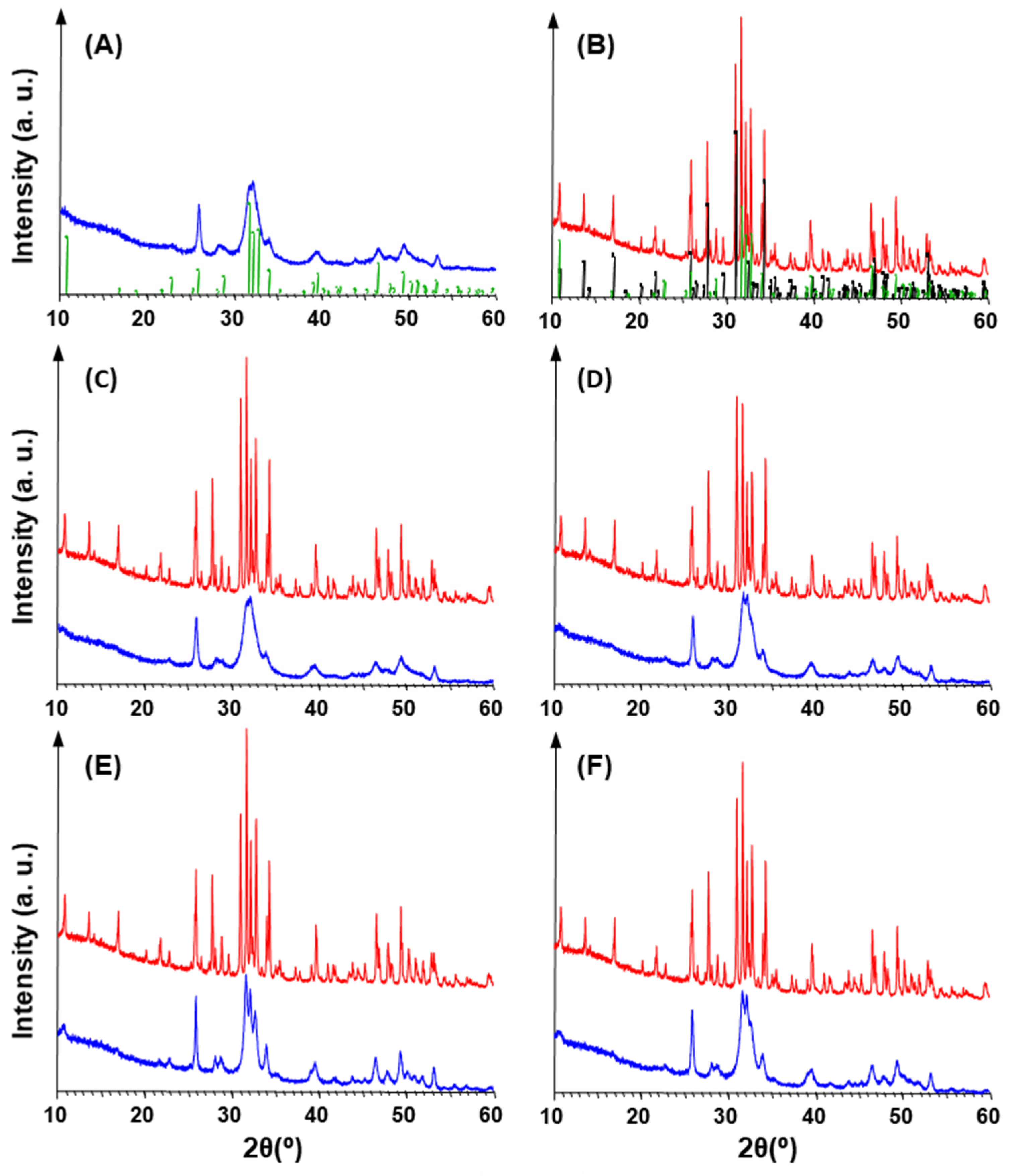
2.5.1. X-ray Diffraction Analysis (XRD)
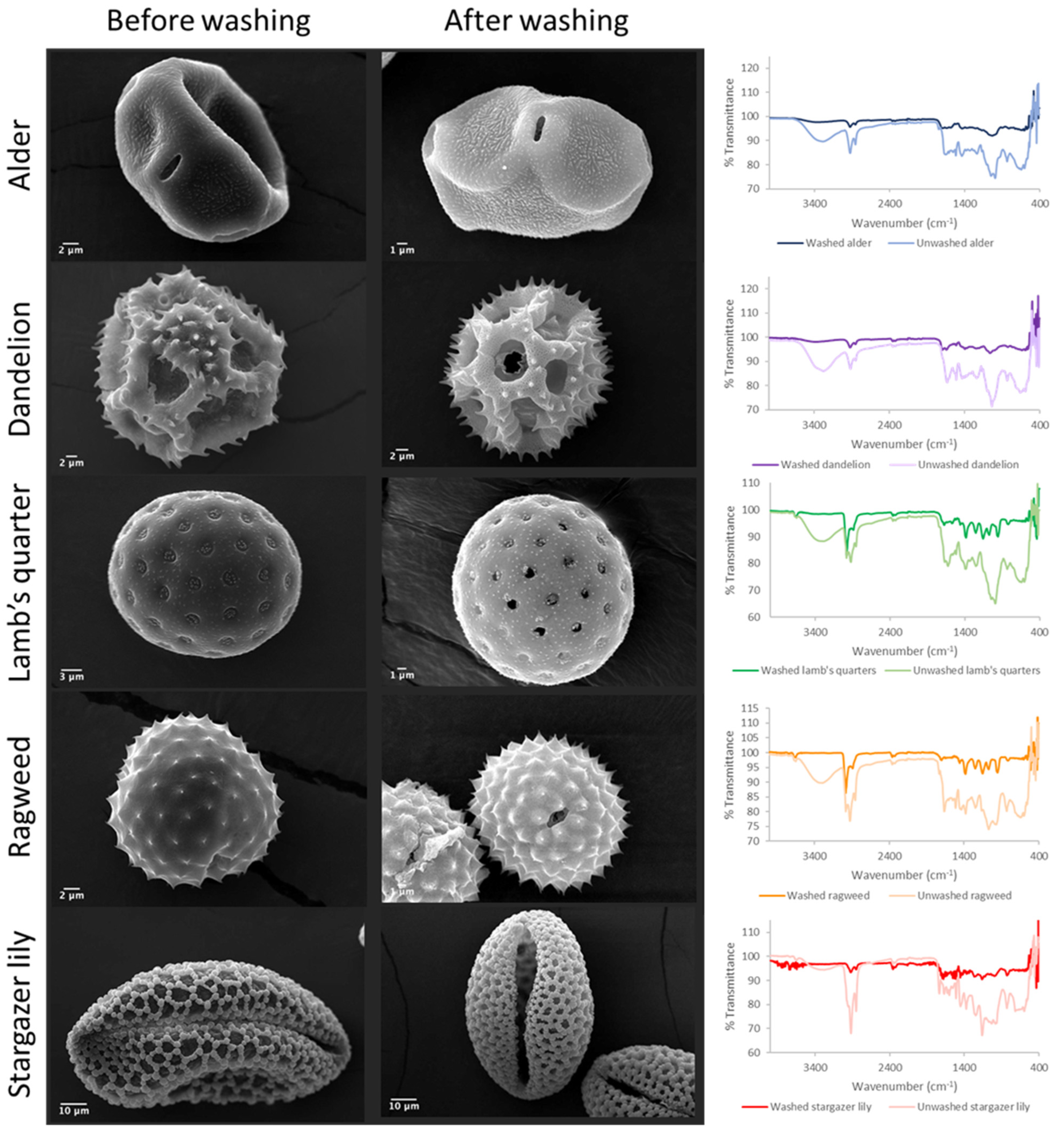
2.5.2. Scanning Electron Microscopy (SEM)
2.5.3. Fourier-Transform Infrared Spectroscopy (FTIR) Analysis
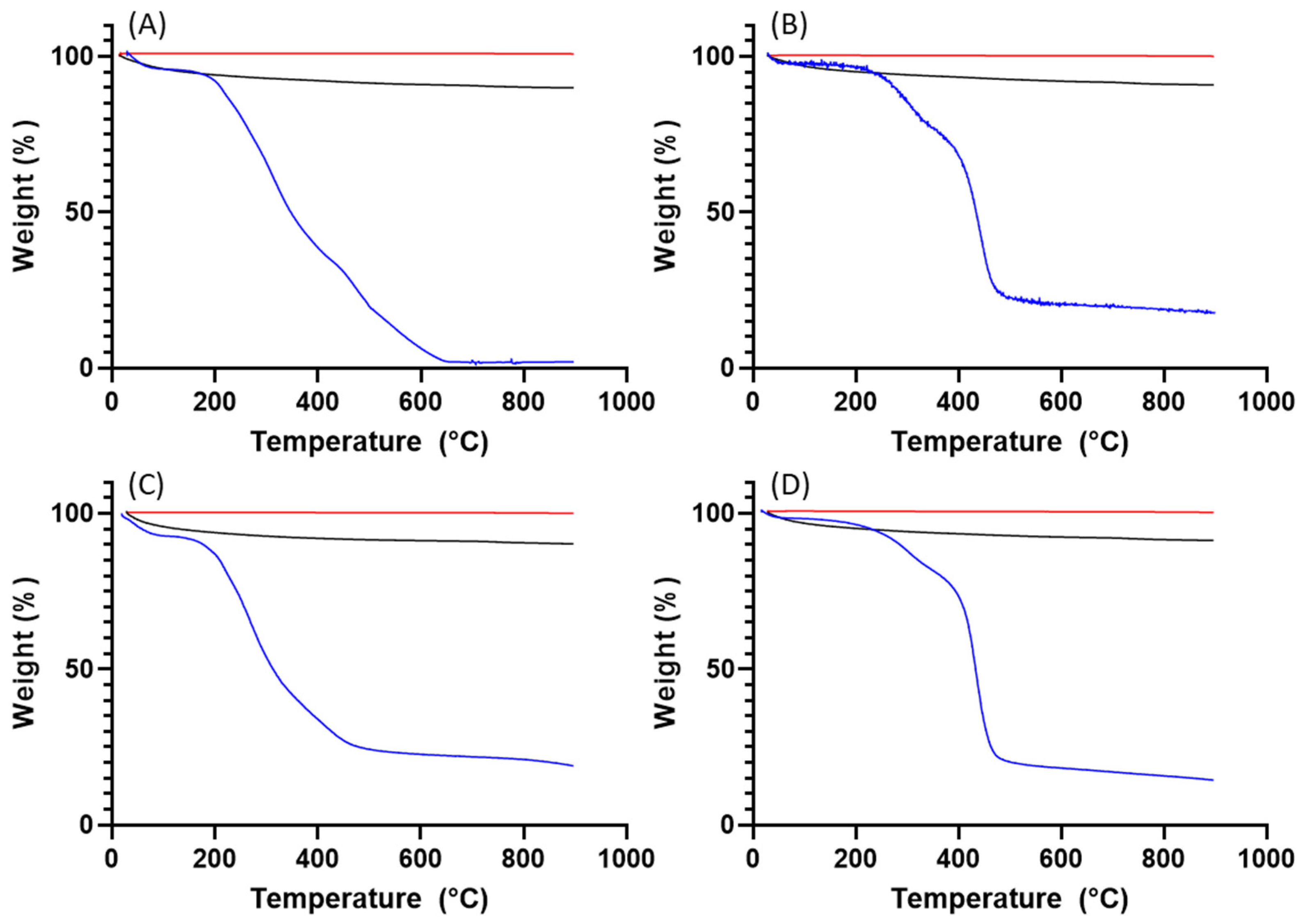
2.5.4. Thermal Gravimetric Analysis (TGA)
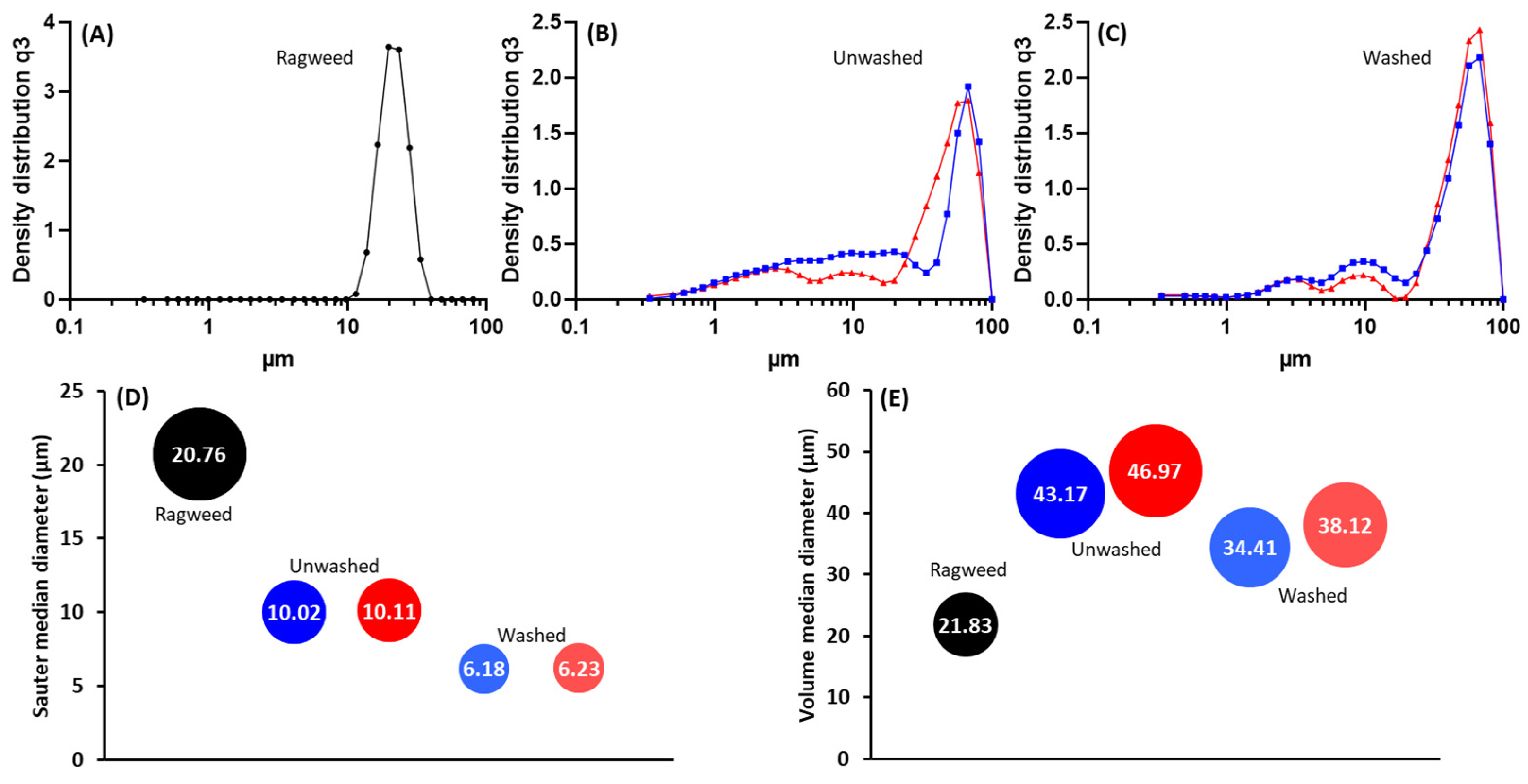
2.5.5. Laser Diffraction
2.6. Statistical Analysis
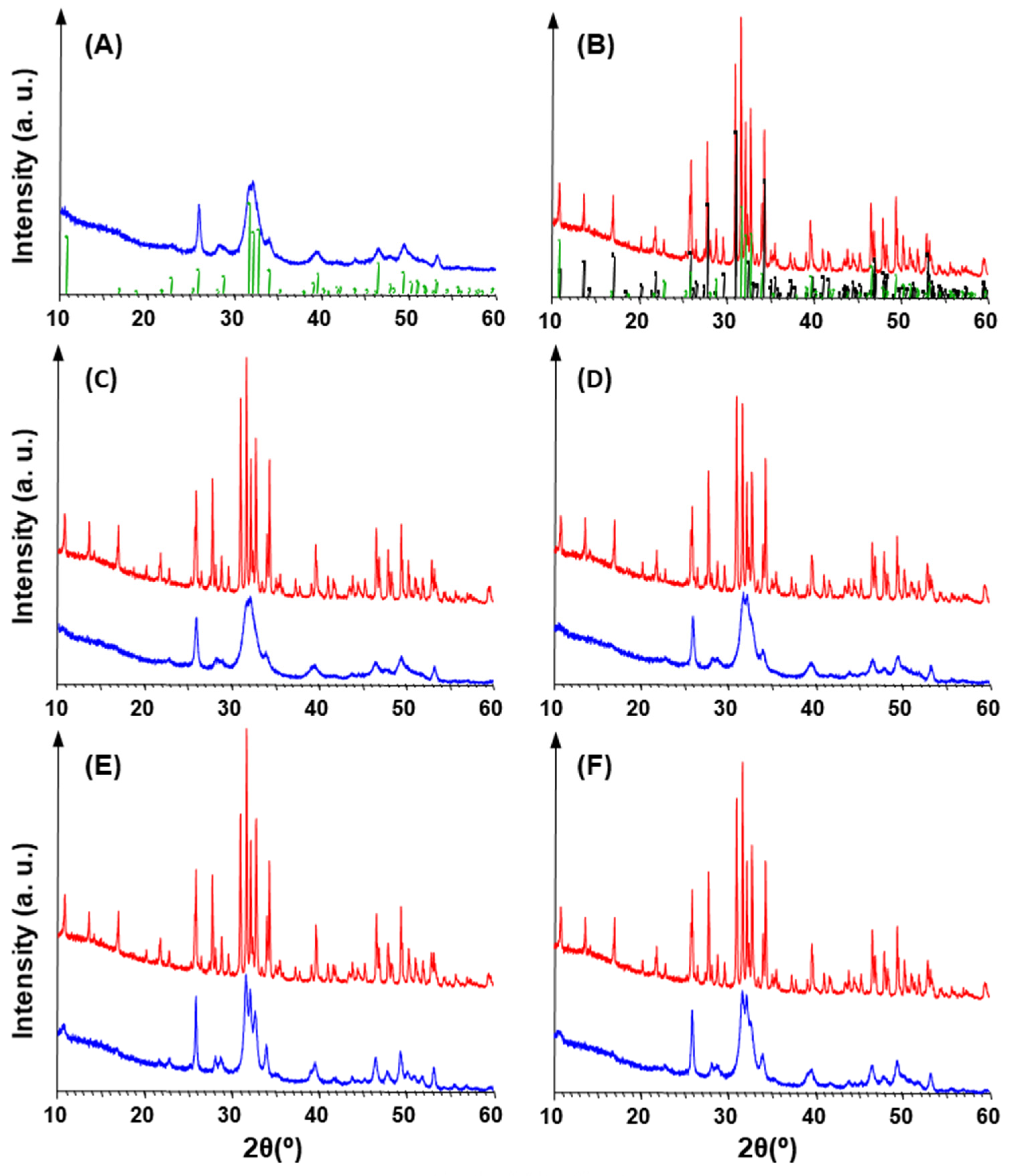
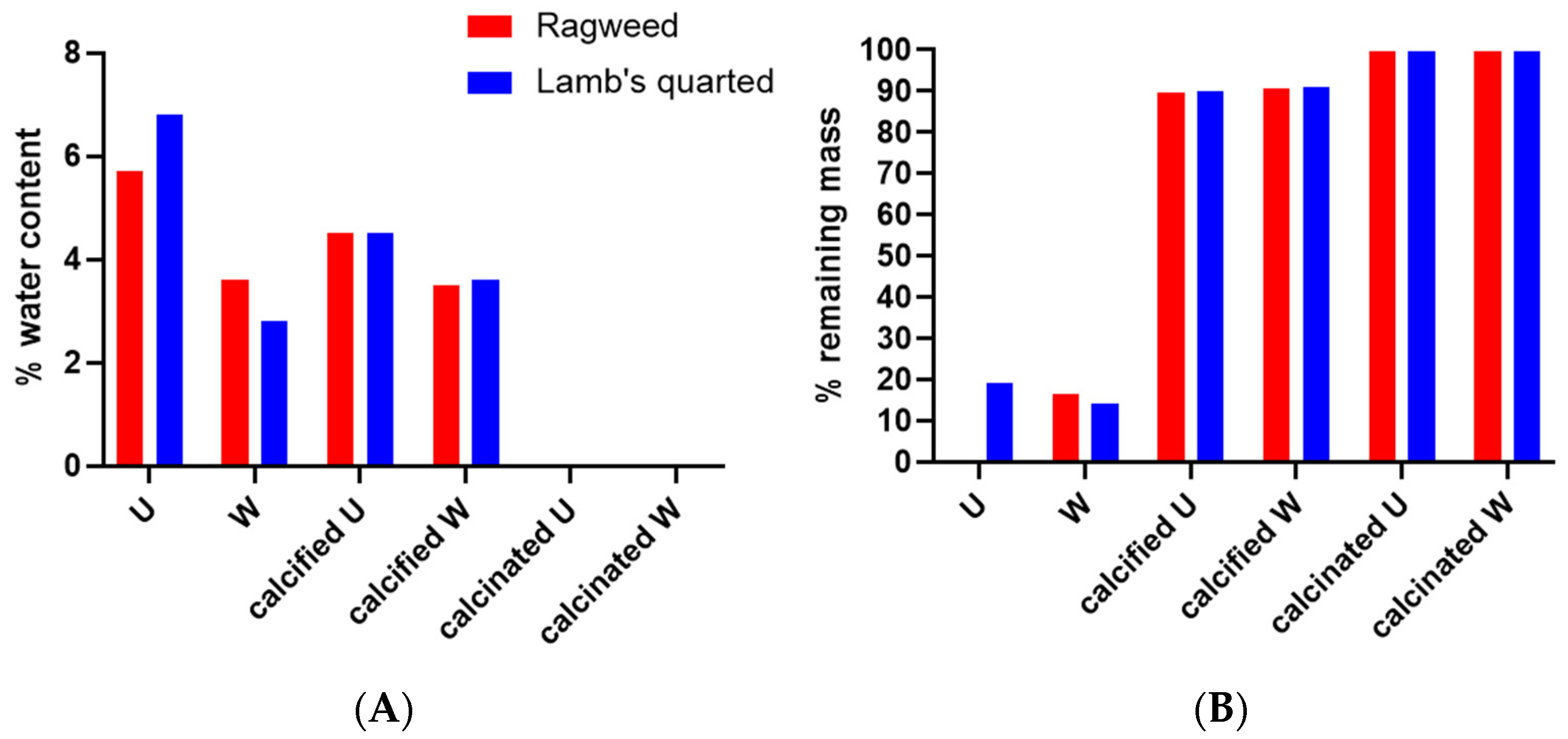
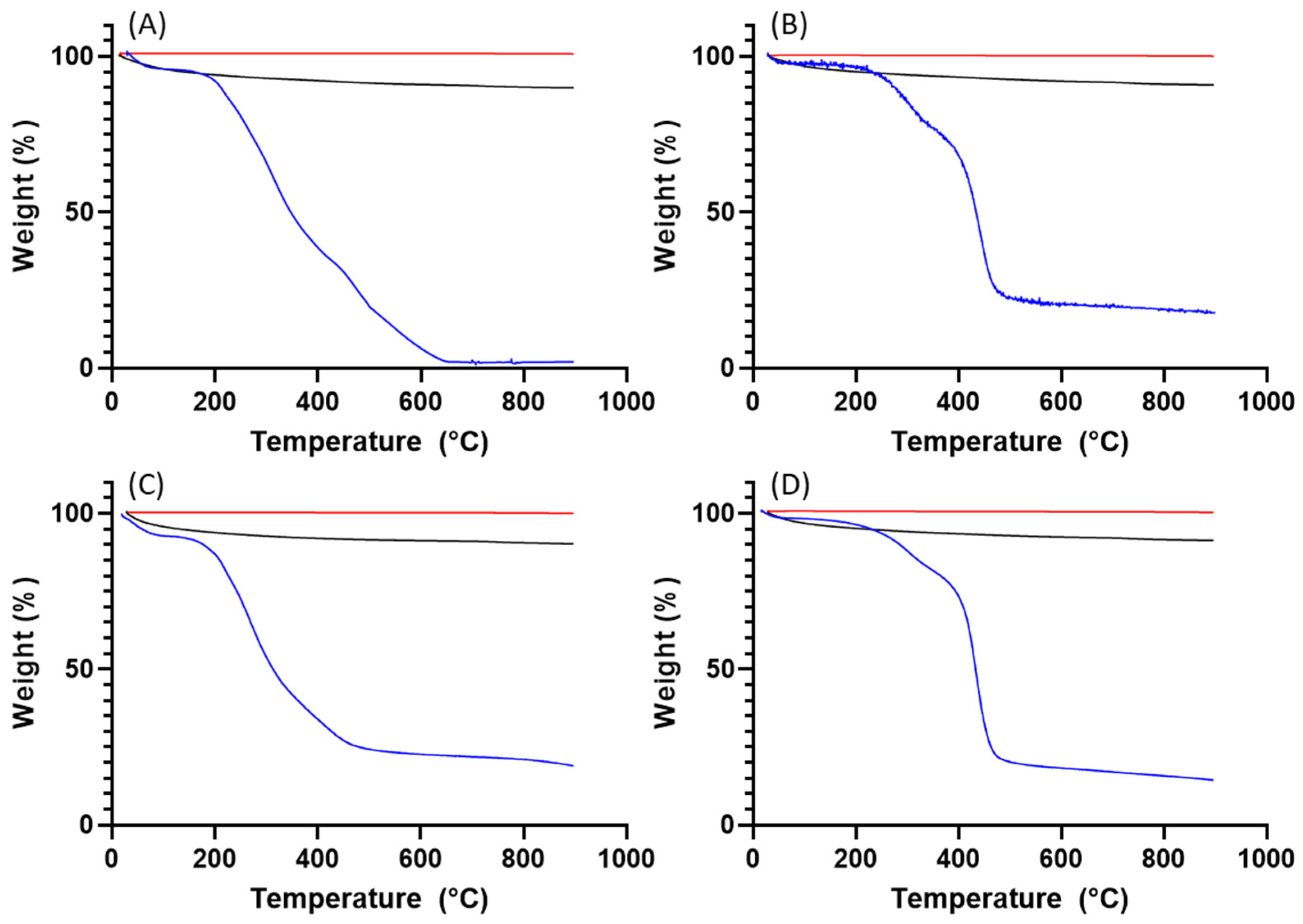
3. Results
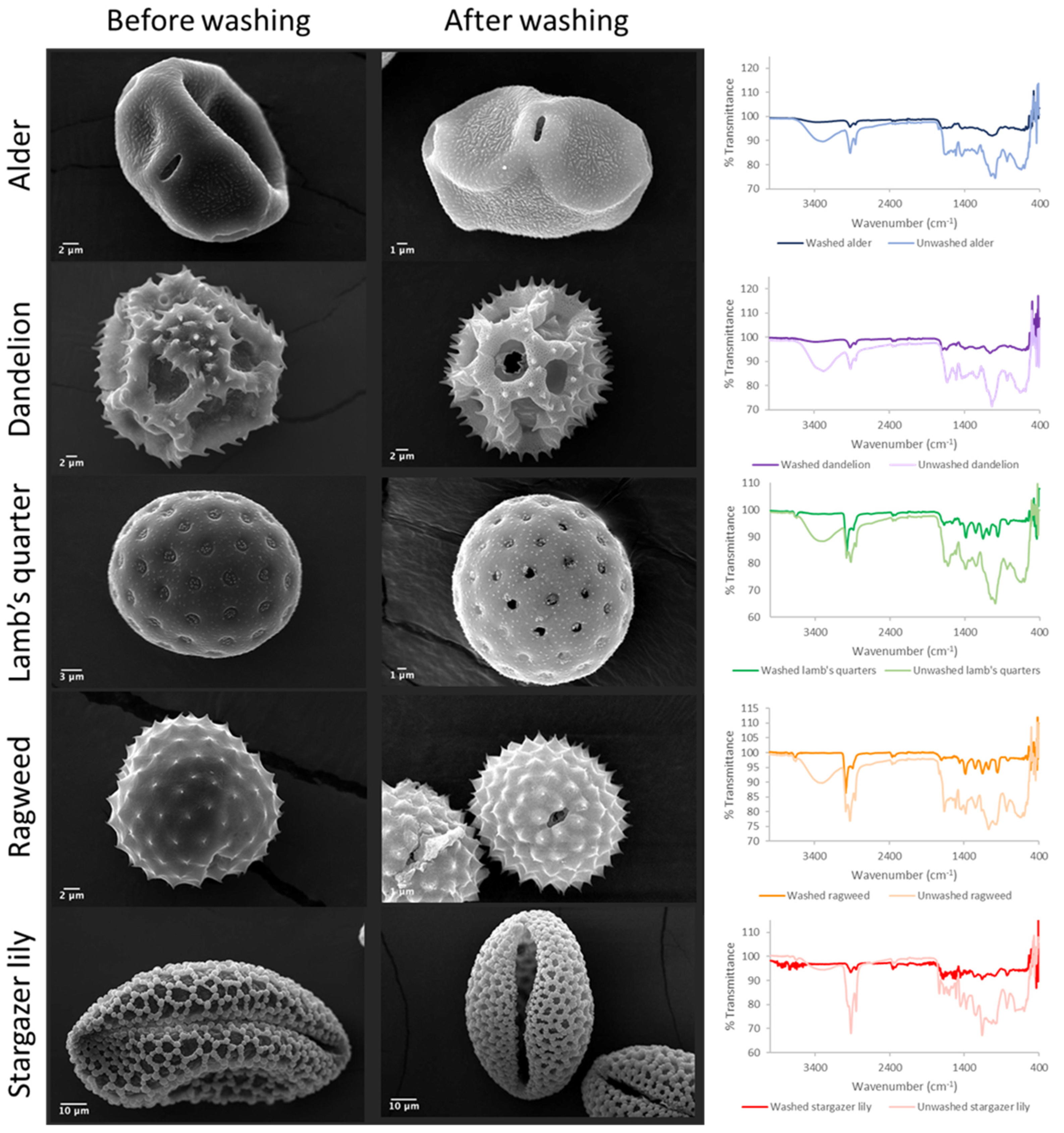
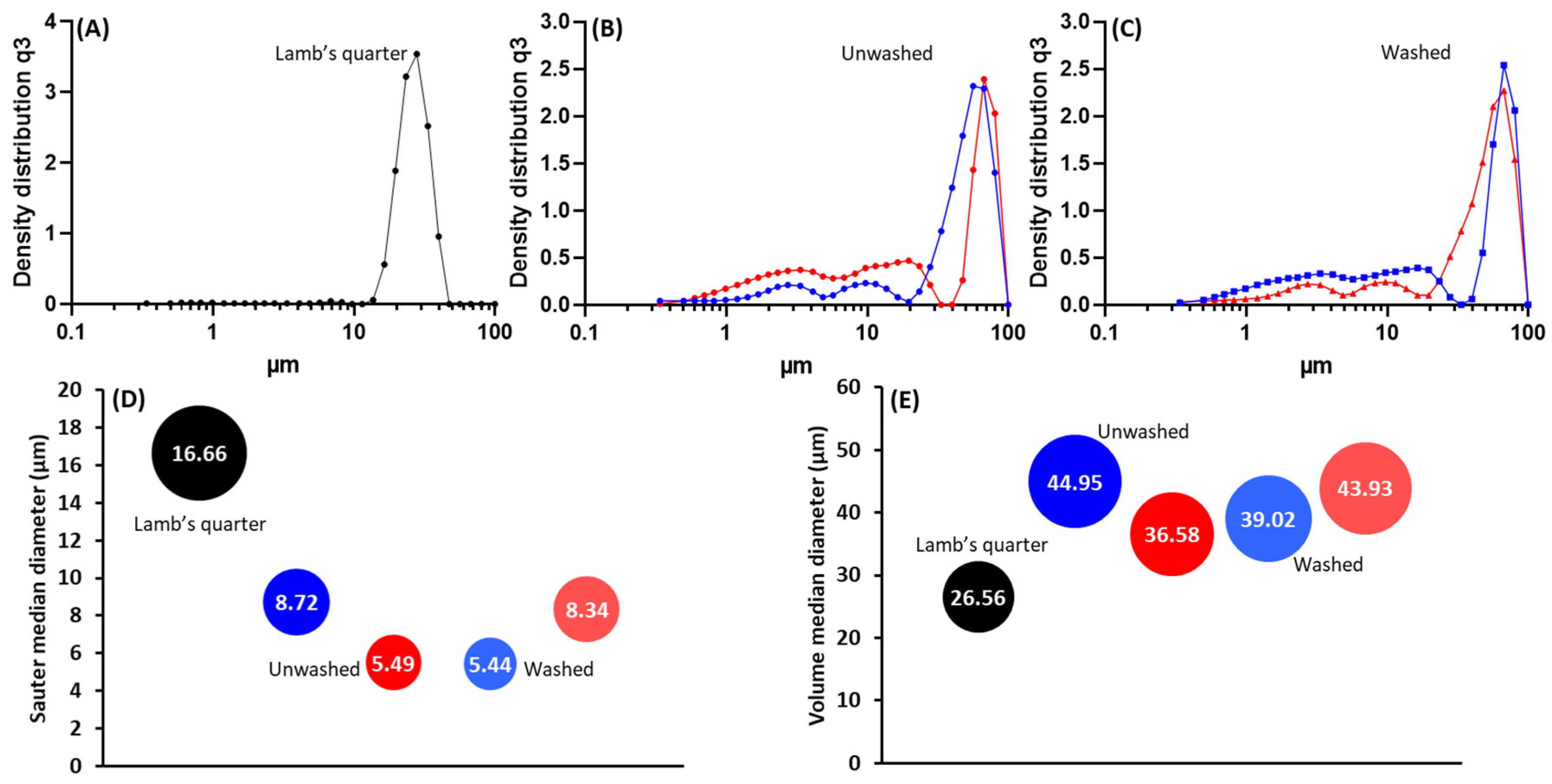
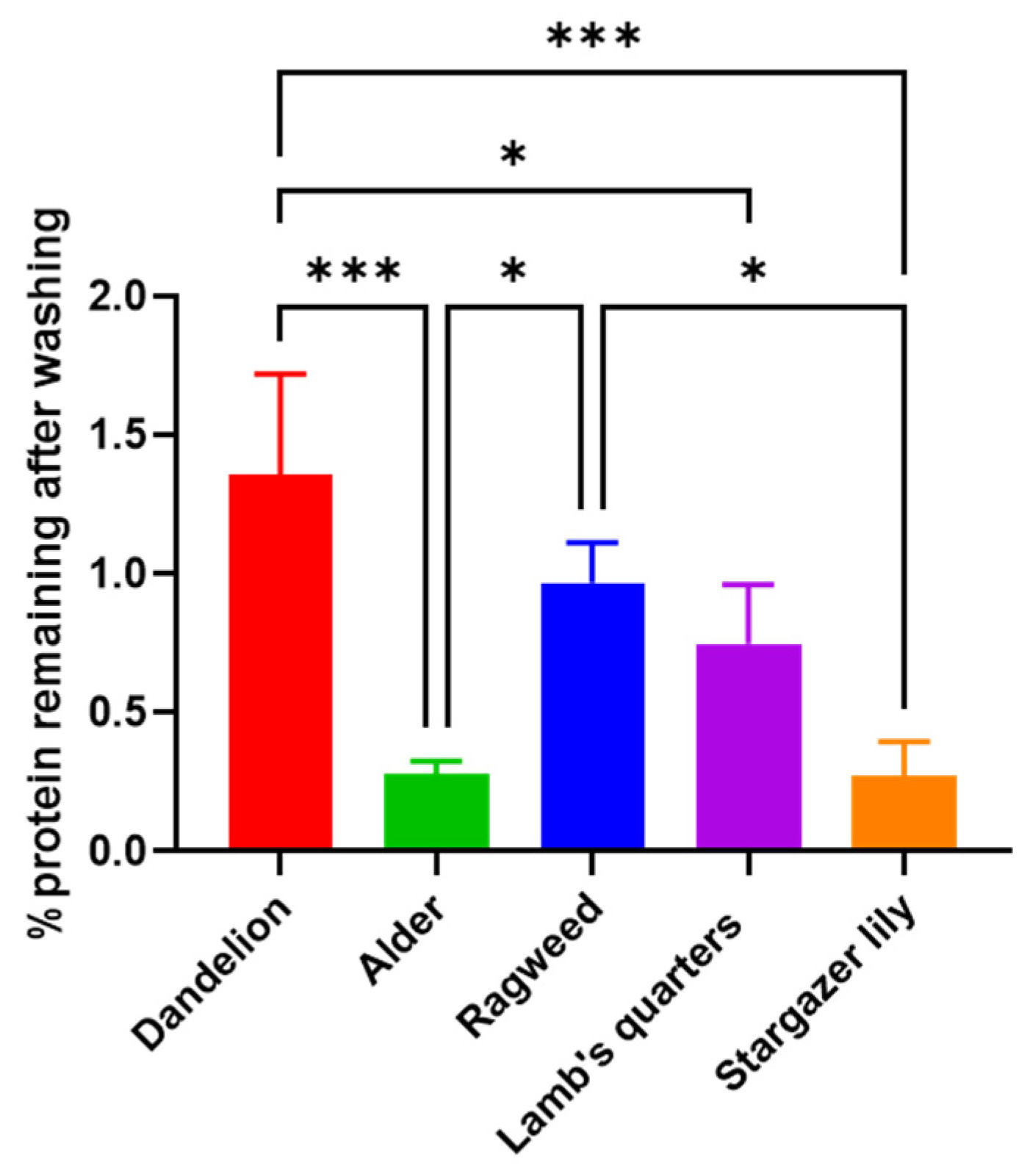
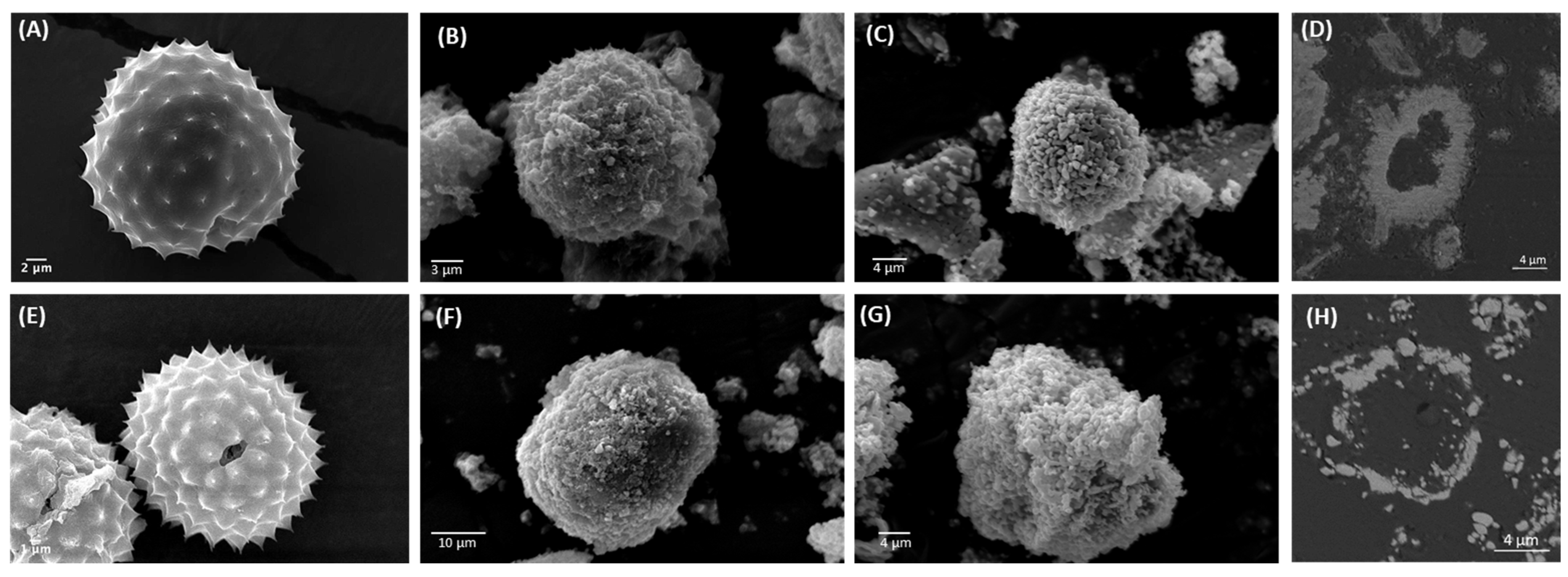
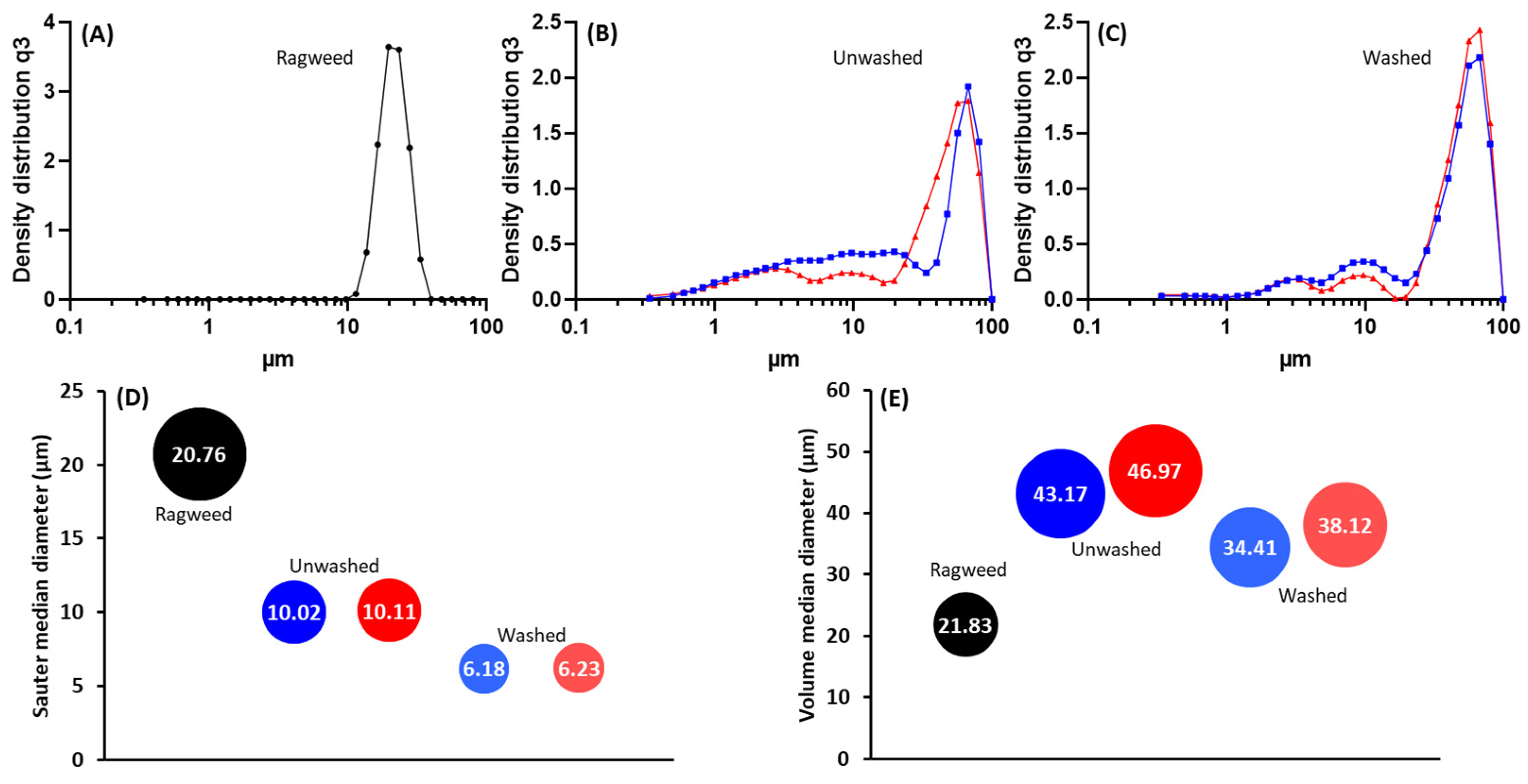
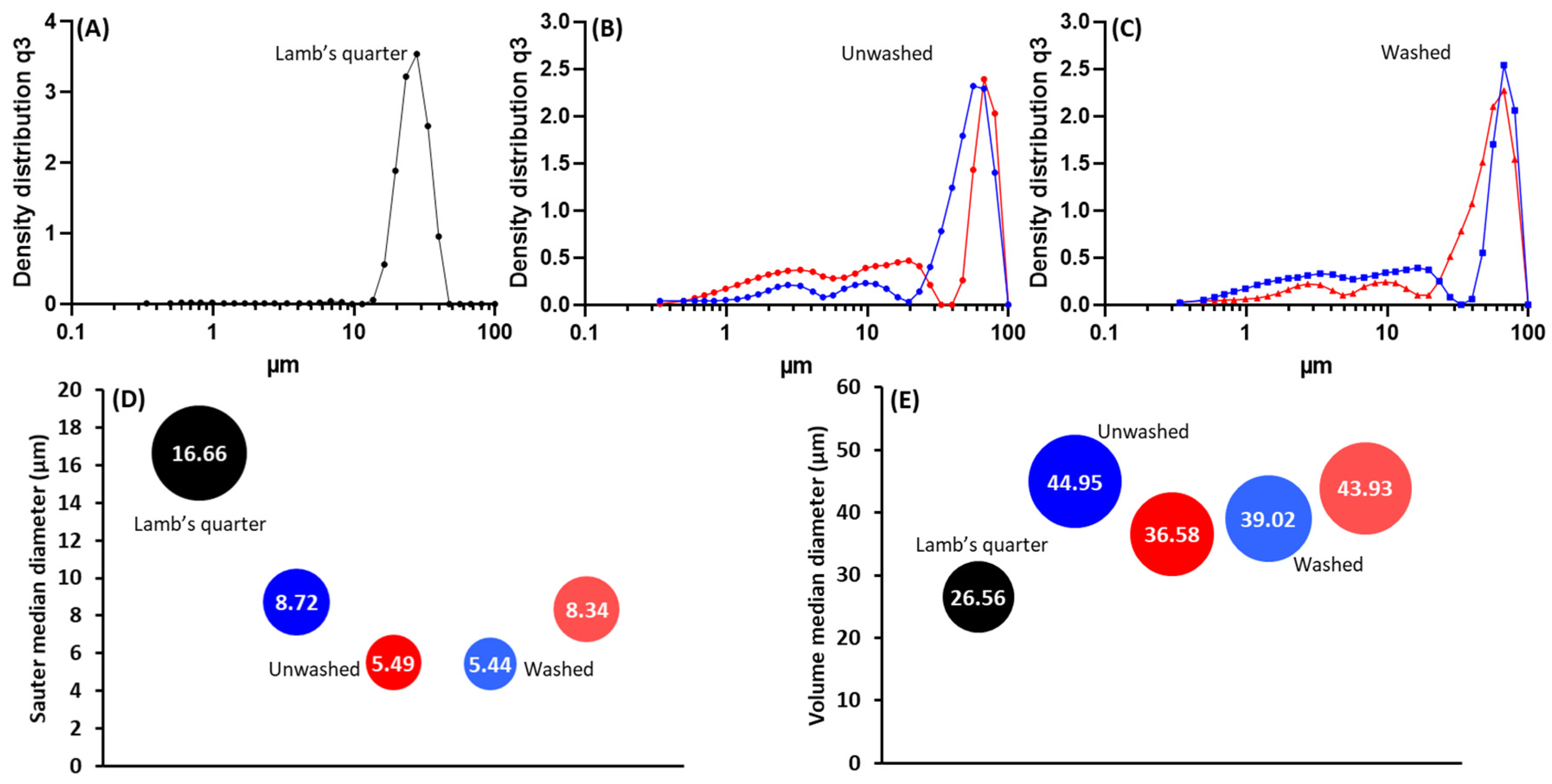
3.1. Purification and Analysis of Sporopollenin Capsules
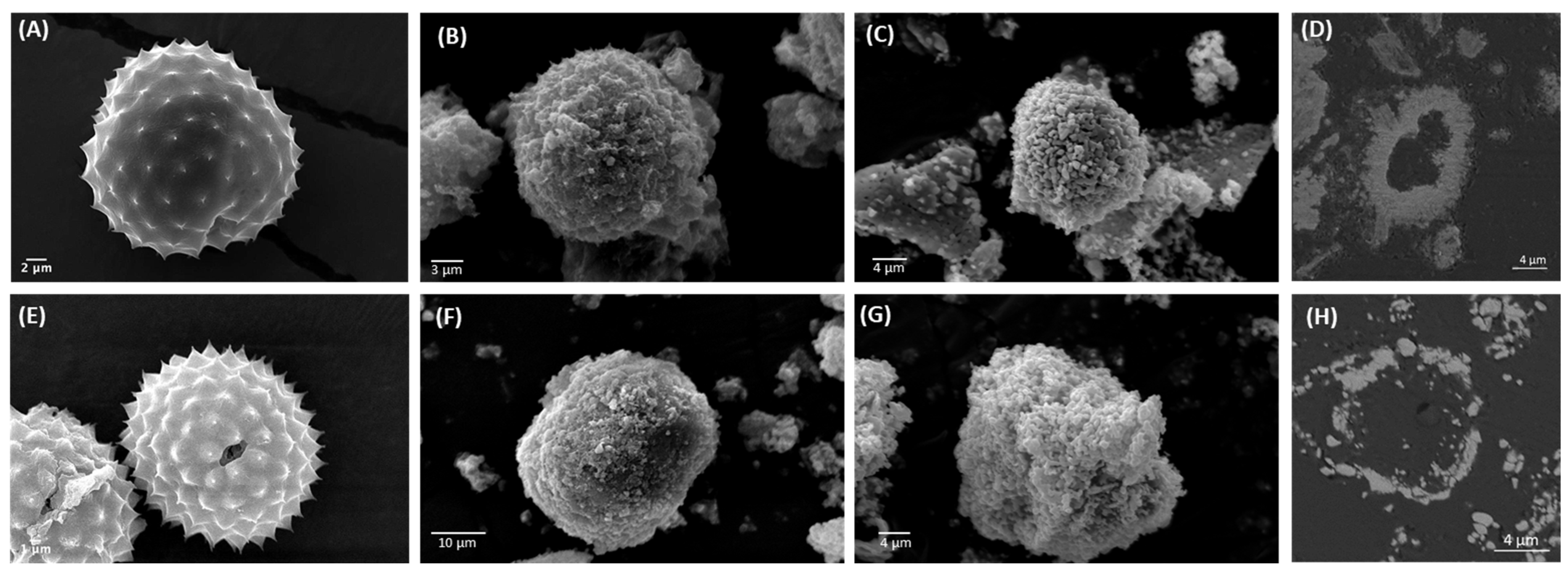
3.2. Pollen Grains as Bio-Templates
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Aylanc, V.; Peixoto, A.F.; Vale, N.; Freire, C.; Vilas-Boas, M. Sporopollenin-based bio-microcapsules as green carriers for controlled delivery of pharmaceutical drugs. Appl. Mater. Today 2023, 33, 101860. [Google Scholar] [CrossRef]
- Atalay, F.E.; Culum, A.A.; Kaya, H.; Gokturk, G.; Yigit, E. Different Plant Sporopollenin Exine Capsules and Their Multifunctional Usage. ACS Appl. Bio Mater. 2022, 5, 1348–1360. [Google Scholar] [CrossRef]
- Mundargi, R.C.; Potroz, M.G.; Park, S.; Shirahama, H.; Lee, J.H.; Seo, J.; Cho, N. Natural Sunflower Pollen as a Drug Delivery Vehicle. Small 2016, 12, 1167–1173. [Google Scholar] [CrossRef]
- Uddin, M.J.; Liyanage, S.; Abidi, N.; Gill, H.S. Physical and Biochemical Characterization of Chemically Treated Pollen Shells for Potential Use in Oral Delivery of Therapeutics. J. Pharm. Sci. 2018, 107, 3047–3059. [Google Scholar] [CrossRef]
- Gonzalez-Cruz, P.; Uddin, M.J.; Atwe, S.U.; Abidi, N.; Gill, H.S. Chemical Treatment Method for Obtaining Clean and Intact Pollen Shells of Different Species. ACS Biomater. Sci. Eng. 2018, 4, 2319–2329. [Google Scholar] [CrossRef]
- Lale, S.V.; Gill, H.S. Pollen grains as a novel microcarrier for oral delivery of proteins. Int. J. Pharm. 2018, 552, 352–359. [Google Scholar] [CrossRef]
- Ávila-Avilés, R.D.; Torres-Gómez, N.; Camacho-López, M.A.; Vilchis-Nestor, A.R. SERS activity of hybrid nano/microstructures Ag-Fe3O4 based on Dimorphotheca ecklonis pollen grains as bio-template. Sci. Rep. 2020, 10, 16633. [Google Scholar] [CrossRef] [PubMed]
- Jardine, P.E.; Palazzesi, L.; Tellería, M.C.; Barreda, V.D. Why does pollen morphology vary? Evolutionary dynamics and morphospace occupation in the largest angiosperm order (Asterales). N. Phytol. 2022, 234, 1075–1087. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Ye, X.; Liu, J. Morphological differences between anemophilous and entomophilous pollen. Microsc. Res. Technol. 2022, 85, 1056–1064. [Google Scholar] [CrossRef] [PubMed]
- Edlund, A.F.; Swanson, R.; Preuss, D. Pollen and Stigma Structure and Function: The Role of Diversity in Pollination. Plant Cell 2004, 16, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Jiao, K.; Yan, J.F.; Wan, M.C.; Wan, Q.Q.; Breschi, L.; Chen, J.H.; Tay, F.R.; Niu, L. na Biological and synthetic template-directed syntheses of mineralized hybrid and inorganic materials. Prog. Mater. Sci. 2021, 116, 100712. [Google Scholar] [CrossRef]
- Zheng, K.; Bortuzzo, J.A.; Liu, Y.; Li, W.; Pischetsrieder, M.; Roether, J.; Lu, M.; Boccaccini, A.R. Bio-templated bioactive glass particles with hierarchical macro–nano porous structure and drug delivery capability. Colloids Surf. B Biointerfaces 2015, 135, 825–832. [Google Scholar] [CrossRef]
- Fan, T.; Park, J.H.; Pham, Q.A.; Tan, E.L.; Mundargi, R.C.; Potroz, M.G.; Jung, H.; Cho, N.J. Extraction of cage-like sporopollenin exine capsules from dandelion pollen grains. Sci. Rep. 2018, 8, 6565. [Google Scholar] [CrossRef]
- Chiappe, C.; Demontis, G.C.; Di Bussolo, V.; Rodriguez Douton, M.J.; Rossella, F.; Pomelli, C.S.; Sartini, S.; Caporali, S. From pollen grains to functionalized microcapsules: A facile chemical route using ionic liquids. Green Chem. 2017, 19, 1028–1033. [Google Scholar] [CrossRef]
- Cao, F.; Li, D. Biotemplate synthesis of monodispersed iron phosphate hollow microspheres. Bioinspiration Biomim. 2010, 5, 16005. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, H.; Costa, C.; Abreu, I.; Esteves da Silva, J.C.G. Effect of O3 and NO2 atmospheric pollutants on Platanus x acerifolia pollen: Immunochemical and spectroscopic analysis. Sci. Total Environ. 2017, 599–600, 291–297. [Google Scholar] [CrossRef]
- Costantini, L.; Bouropoulos, N.; Fatouros, D.G.; Kontopoulou, I.; Roldo, M. Synthesis of carbon nanotubes loaded hydroxyapatite: Potential for controlled drug release from bone implants. J. Adv. Ceram. 2016, 5, 232–243. [Google Scholar] [CrossRef]
- Hammood, A.S.; Hassan, S.S.; Alkhafagy, M.T. Comparison of Natural and Nano-synthetically-Produced Hydroxyapatite Powder. JOM 2019, 71, 272–278. [Google Scholar] [CrossRef]
- Wu, S.C.; Hsu, H.C.; Hsu, S.K.; Chang, Y.C.; Ho, W.F. Synthesis of hydroxyapatite from eggshell powders through ball milling and heat treatment. J. Asian Ceram. Soc. 2016, 4, 85–90. [Google Scholar] [CrossRef]
- Xiong, X.; Zhou, D.; Xu, L.; Liu, T.; Yue, X.; Liu, W.; Cao, J. BcPME37c is involved in pollen intine formation in Brassica campestris. Biochem. Biophys. Res. Commun. 2019, 517, 63–68. [Google Scholar] [CrossRef]
- Roulston, A.H.; Cane, J.H.; Buchmann, S.L. What Governs Protein Content of Pollen: Pollinator Preferences, Pollen-Pistil Interactions or Phylogeny? Ecol. Monogr. 2000, 70, 617–643. [Google Scholar]
- Yasmeen, S.; Lo, M.K.; Bajracharya, S.; Roldo, M. Injectable scaffolds for bone regeneration. Langmuir 2014, 30, 12977–12985. [Google Scholar] [CrossRef]
- LeGeros, R.Z. Calcium Phosphate-Based Osteoinductive Materials. Chem. Rev. 2008, 108, 4742–4753. [Google Scholar] [CrossRef] [PubMed]
- Juang, H.Y.; Hon, M.H. Effect of calcination on sintering of hydroxyapatite. Biomaterials 1996, 17, 2059–2064. [Google Scholar] [CrossRef]
- Tõnsuaadu, K.; Gross, K.A.; Plūduma, L.; Veiderma, M. A review on the thermal stability of calcium apatites. J. Therm. Anal. Calorim. 2012, 110, 647–659. [Google Scholar] [CrossRef]
- Bulina, N.V.; Makarova, S.V.; Baev, S.G.; Matvienko, A.A.; Gerasimov, K.B.; Logutenko, O.A.; Bystrov, V.S. A Study of Thermal Stability of Hydroxyapatite. Minerals 2021, 11, 1310. [Google Scholar] [CrossRef]
- Cao, F.; Li, D.X. Morphology-controlled synthesis of SiO2 hollow microspheres using pollen grain as a biotemplate. Biomed. Mater. 2009, 4, 025009. [Google Scholar] [CrossRef]
- Poncet, P.; Sénéchal, H.; Charpin, D. Update on pollen-food allergy syndrome. Expert Rev. Clin. Immunol. 2020, 16, 561–578. [Google Scholar] [CrossRef]
- Visez, N.; de Nadaï, P.; Choël, M.; Farah, J.; Hamzé, M.; Sénéchal, H.; Pauwels, M.; Frérot, H.; Thibaudon, M.; Poncet, P. Biochemical composition of Phleum pratense pollen grains: A review. Mol. Immunol. 2021, 136, 98–109. [Google Scholar] [CrossRef]
- Maruthi, Y.A.; Ramakrishna, S. Sporopollenin—Invincible biopolymer for sustainable biomedical applications. Int. J. Biol. Macromol. 2022, 222, 2957–2965. [Google Scholar] [CrossRef]
- Bohner, M.; Santoni, B.L.G.; Döbelin, N. β-Tricalcium Phosphate for Bone Substitution: Synthesis and Properties. Acta Mater. 2020, 113, 23–41. [Google Scholar]
- Rakovan, J.F.; Pasteris, J.D. A Technological Gem: Materials, Medical, and Environmental Mineralogy of Apatite. Elements 2015, 11, 195–200. [Google Scholar] [CrossRef]
- Chang, B.-S.; Lee, C.-K.; Hong, K.-S.; Youn, H.-J.; Ryu, H.-S.; Chung, S.-S.; Park, K.-W. Osteoconduction at porous hydroxyapatite with various pore configurations. Biomaterials 2000, 21, 1291–1298. [Google Scholar] [CrossRef] [PubMed]
- Fiume, E.; Magnaterra, G.; Rahdar, A.; Verné, E.; Baino, F. Hydroxyapatite for biomedical applications: A short overview. Ceramics 2021, 4, 542–563. [Google Scholar] [CrossRef]
- Tozzi, G.; De Mori, A.; Oliveira, A.; Roldo, M. Composite Hydrogels for Bone Regeneration. Materials 2016, 9, 267. [Google Scholar] [CrossRef] [PubMed]
- De Mori, A.; Fernández, M.P.; Blunn, G.; Tozzi, G.; Roldo, M. 3D printing and electrospinning of composite hydrogels for cartilage and bone tissue engineering. Polymers 2018, 10, 285. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pollen Species | Defatted | Percentage Yield (%) | Length Unwashed (µm) | Length Washed (µm) |
|---|---|---|---|---|
| Alder | No | 6.5 | 24.9 ± 0.7 | 21.7 ± 0.9 a |
| Dandelion | Yes | 15.9 | 33.6 ± 2.7 | 26.0 ± 2.9 b |
| Lamb’s quarters | Yes | 5.0 | 27.4 ± 0.6 | 22.5 ± 1.4 c |
| Ragweed | No | 19.8 | 20.7 ± 1.2 | 17.2 ± 2.3 d |
| Stargazer lily | No | 3.5 | 107.0 ± 16.5 | 74.7 ± 4.5 e |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Mori, A.; Quizon, D.; Dalton, H.; Yavuzyegit, B.; Cerri, G.; Antonijevic, M.; Roldo, M. Sporopollenin Capsules as Biomimetic Templates for the Synthesis of Hydroxyapatite and β-TCP. Biomimetics 2024, 9, 159. https://doi.org/10.3390/biomimetics9030159
De Mori A, Quizon D, Dalton H, Yavuzyegit B, Cerri G, Antonijevic M, Roldo M. Sporopollenin Capsules as Biomimetic Templates for the Synthesis of Hydroxyapatite and β-TCP. Biomimetics. 2024; 9(3):159. https://doi.org/10.3390/biomimetics9030159
Chicago/Turabian StyleDe Mori, Arianna, Daniel Quizon, Hannah Dalton, Berzah Yavuzyegit, Guido Cerri, Milan Antonijevic, and Marta Roldo. 2024. "Sporopollenin Capsules as Biomimetic Templates for the Synthesis of Hydroxyapatite and β-TCP" Biomimetics 9, no. 3: 159. https://doi.org/10.3390/biomimetics9030159
APA StyleDe Mori, A., Quizon, D., Dalton, H., Yavuzyegit, B., Cerri, G., Antonijevic, M., & Roldo, M. (2024). Sporopollenin Capsules as Biomimetic Templates for the Synthesis of Hydroxyapatite and β-TCP. Biomimetics, 9(3), 159. https://doi.org/10.3390/biomimetics9030159