
Multifunctionality in Nature: Structure–Function Relationships in Biological Materials


Abstract
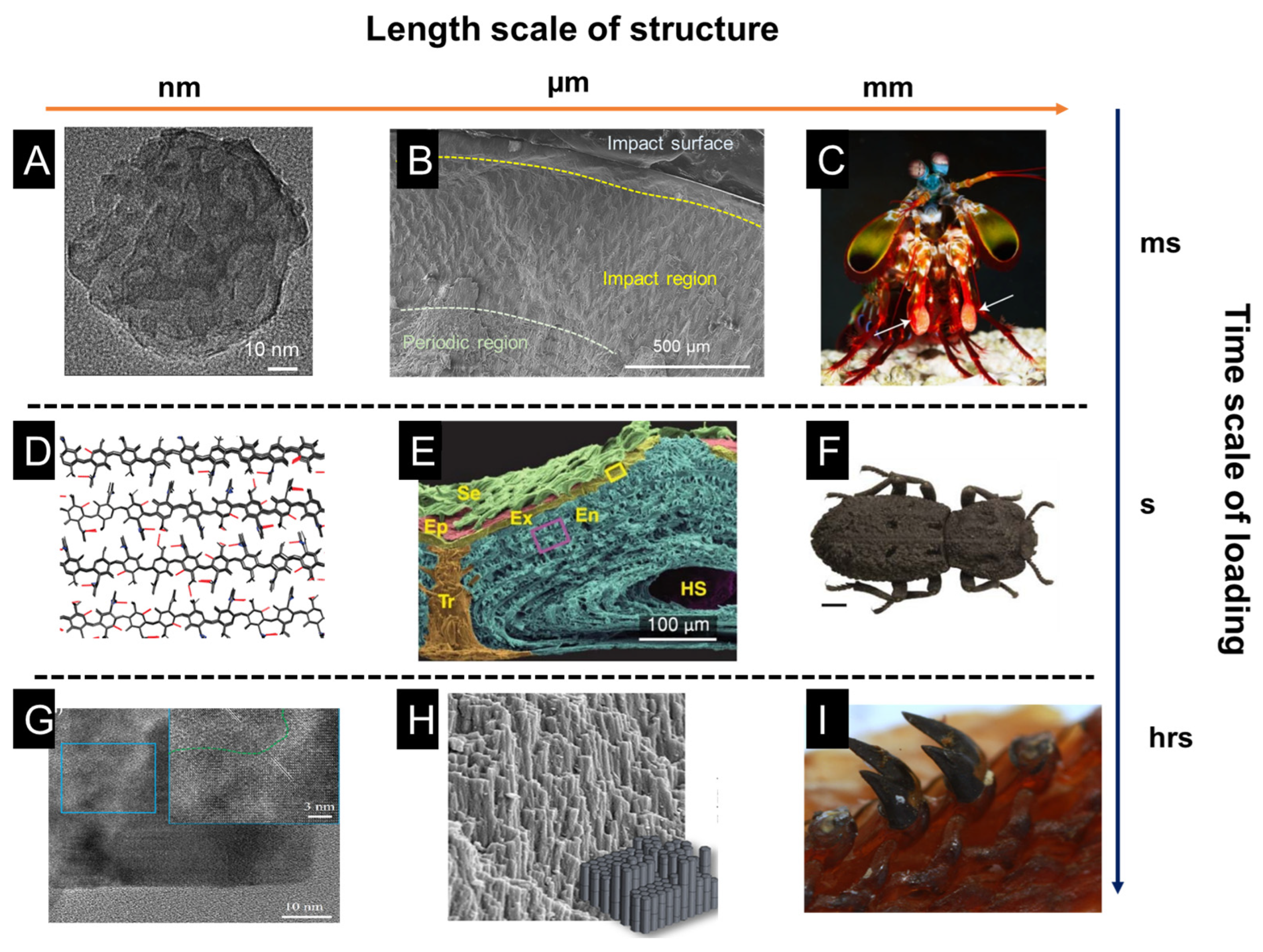
1. Introduction
2. Impact, Wear, and Crush Resistance
3. Self-Healing and Shape Memory Effect
3.1. Spider Silk
3.2. Mussel Byssal Threads
4. Adhesion and Anti-Fouling
4.1. Mussel Byssus Plaque
4.2. Gecko Toe Pads
5. Sensing, Actuating, and Camouflage
5.1. Moth Wings
5.2. Chameleons
5.3. Banksia Seed Pods
6. Life in Extreme Environments
7. Bioinspired Applications and Challenges
7.1. Bioinspired Applications
7.2. Challenges
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wegst, U.G.K.; Bai, H.; Saiz, E.; Tomsia, A.P.; Ritchie, R.O. Bioinspired Structural Materials. Nat. Mater. 2015, 14, 23–36. [Google Scholar] [CrossRef]
- Wang, Y.; Naleway, S.E.; Wang, B. Biological and Bioinspired Materials: Structure Leading to Functional and Mechanical Performance. Bioact. Mater. 2020, 5, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Eder, M.; Amini, S.; Fratzl, P. Biological Composites-Complex Structures for Functional Diversity. Science 2018, 362, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Meyers, M.A.; Chen, P.-Y.; Lin, A.Y.-M.; Seki, Y. Biological Materials: Structure and Mechanical Properties. Prog. Mater. Sci. 2008, 53, 1–206. [Google Scholar] [CrossRef]
- Meyers, M.A.; McKittrick, J.; Chen, P.-Y. Structural Biological Materials: Critical Mechanics-Materials Connections. Science 2013, 339, 773–779. [Google Scholar] [CrossRef]
- Nepal, D.; Kang, S.; Adstedt, K.M.; Kanhaiya, K.; Bockstaller, M.R.; Brinson, L.C.; Buehler, M.J.; Coveney, P.V.; Dayal, K.; El-Awady, J.A.; et al. Hierarchically Structured Bioinspired Nanocomposites. Nat. Mater. 2023, 22, 18–35. [Google Scholar] [CrossRef]
- Barthelat, F.; Yin, Z.; Buehler, M.J. Structure and Mechanics of Interfaces in Biological Materials. Nat. Rev. Mater. 2016, 1, 16007. [Google Scholar] [CrossRef]
- Mayer, G. Rigid Biological Systems as Models for Synthetic Composites. Science 2005, 310, 1144–1147. [Google Scholar] [CrossRef]
- Huang, W.; Restrepo, D.; Jung, J.; Su, F.Y.; Liu, Z.; Ritchie, R.O.; McKittrick, J.; Zavattieri, P.; Kisailus, D. Multiscale Toughening Mechanisms in Biological Materials and Bioinspired Designs. Adv. Mater. 2019, 31, 1901561. [Google Scholar] [CrossRef]
- Espinosa, H.D.; Rim, J.E.; Barthelat, F.; Buehler, M.J. Merger of Structure and Material in Nacre and Bone-Perspectives on de Novo Biomimetic Materials. Prog. Mater. Sci. 2009, 54, 1059–1100. [Google Scholar] [CrossRef]
- Waite, J.H. Mussel Adhesion—Essential Footwork. J. Exp. Biol. 2017, 220, 517–530. [Google Scholar] [CrossRef]
- Schmitt, C.N.Z.; Politi, Y.; Reinecke, A.; Harrington, M.J. Role of Sacrificial Protein–Metal Bond Exchange in Mussel Byssal Thread Self-Healing. Biomacromolecules 2015, 16, 2852–2861. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Shishehbor, M.; Guarín-Zapata, N.; Kirchhofer, N.D.; Li, J.; Cruz, L.; Wang, T.; Bhowmick, S.; Stauffer, D.; Manimunda, P.; et al. A Natural Impact-Resistant Bicontinuous Composite Nanoparticle Coating. Nat. Mater. 2020, 19, 1236–1243. [Google Scholar] [CrossRef]
- Rivera, J.; Hosseini, M.S.; Restrepo, D.; Murata, S.; Vasile, D.; Parkinson, D.Y.; Barnard, H.S.; Arakaki, A.; Zavattieri, P.; Kisailus, D. Toughening Mechanisms of the Elytra of the Diabolical Ironclad Beetle. Nature 2020, 586, 543–548. [Google Scholar] [CrossRef]
- Anton, A.M.; Heidebrecht, A.; Mahmood, N.; Beiner, M.; Scheibel, T.; Kremer, F. Foundation of the Outstanding Toughness in Biomimetic and Natural Spider Silk. Biomacromolecules 2017, 18, 3954–3962. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Mcadams, D.A.; Grunlan, J.C. Nano/Micro-Manufacturing of Bioinspired Materials: A Review of Methods to Mimic Natural Structures. Adv. Mater. 2016, 28, 6292–6321. [Google Scholar] [CrossRef]
- Stavenga, D.G.; Wallace, J.R.A.; Warrant, E.J. Bogong Moths Are Well Camouflaged by Effectively Decolourized Wing Scales. Front. Physiol. 2020, 11, 95. [Google Scholar] [CrossRef]
- Teyssier, J.; Saenko, S.V.; van der Marel, D.; Milinkovitch, M.C. Photonic Crystals Cause Active Colour Change in Chameleons. Nat. Commun. 2015, 6, 6368. [Google Scholar] [CrossRef]
- Huang, W.; Ertekin, E.; Wang, T.; Cruz, L.; Dailey, M.; DiRuggiero, J.; Kisailus, D. Mechanism of Water Extraction from Gypsum Rock by Desert Colonizing Microorganisms. Proc. Natl. Acad. Sci. USA 2020, 117, 10681–10687. [Google Scholar] [CrossRef] [PubMed]
- Barthelat, F. Architectured Materials in Engineering and Biology: Fabrication, Structure, Mechanics and Performance. Int. Mater. Rev. 2015, 60, 413–430. [Google Scholar] [CrossRef]
- Naleway, S.E.; Porter, M.M.; McKittrick, J.; Meyers, M.A. Structural Design Elements in Biological Materials: Application to Bioinspiration. Adv. Mater. 2015, 27, 5455–5476. [Google Scholar] [CrossRef]
- Yao, H.-B.; Ge, J.; Mao, L.-B.; Yan, Y.-X.; Yu, S.-H. 25th Anniversary Article: Artificial Carbonate Nanocrystals and Layered Structural Nanocomposites Inspired by Nacre: Synthesis, Fabrication and Applications. Adv. Mater. 2014, 26, 163–188. [Google Scholar] [CrossRef]
- Addadi, L.; Joester, D.; Nudelman, F.; Weiner, S. Mollusk Shell Formation: A Source of New Concepts for Understanding Biomineralization Processes. Chem. Eur. J. 2006, 12, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.-B.; Gao, H.-L.; Yao, H.-B.; Liu, L.; Cölfen, H.; Liu, G.; Chen, S.-M.; Li, S.-K.; Yan, Y.-X.; Liu, Y.-Y.; et al. Synthetic Nacre by Predesigned Matrix-Directed Mineralization. Science 2016, 354, 107–110. [Google Scholar] [CrossRef]
- Checa, A.G.; Cartwright, J.H.E.; Willinger, M.-G. The Key Role of the Surface Membrane in Why Gastropod Nacre Grows in Towers. Proc. Natl. Acad. Sci. USA 2009, 106, 38–43. [Google Scholar] [CrossRef]
- Kuprian, E.; Munkler, C.; Resnyak, A.; Zimmermann, S.; Tuong, T.D.; Gierlinger, N.; Müller, T.; Livingston, D.P.; Neuner, G. Complex Bud Architecture and Cell-Specific Chemical Patterns Enable Supercooling of Picea abies Bud Primordia. Plant Cell Environ. 2017, 40, 3101–3112. [Google Scholar] [CrossRef]
- Moyroud, E.; Wenzel, T.; Middleton, R.; Rudall, P.J.; Banks, H.; Reed, A.; Mellers, G.; Killoran, P.; Westwood, M.M.; Steiner, U.; et al. Disorder in Convergent Floral Nanostructures Enhances Signalling to Bees. Nature 2017, 550, 469–474. [Google Scholar] [CrossRef]
- Sullivan, T.N.; Wang, B.; Espinosa, H.D.; Meyers, M.A. Extreme Lightweight Structures: Avian Feathers and Bones. Mater. Today 2017, 20, 377–391. [Google Scholar] [CrossRef]
- Torquato, S.; Hyun, S.; Donev, A. Optimal Design of Manufacturable Three-Dimensional Composites with Multifunctional Characteristics. J. Appl. Phys. 2003, 94, 5748–5755. [Google Scholar] [CrossRef]
- Ashby, M.F.; Bréchet, Y.J.M.; Cebon, D.; Salvo, L. Selection Strategies for Materials and Processes. Mater. Des. 2004, 25, 51–67. [Google Scholar] [CrossRef]
- Chandler, C.J.; Wilts, B.D.; Brodie, J.; Vignolini, S. Structural Color in Marine Algae. Adv. Opt. Mater. 2017, 5, 1600646. [Google Scholar] [CrossRef]
- Arzt, E. Biological and Artificial Attachment Devices: Lessons for Materials Scientists from Flies and Geckos. Mater. Sci. Eng. C 2006, 26, 1245–1250. [Google Scholar] [CrossRef]
- Studart, A.R. Biological and Bioinspired Composites with Spatially Tunable Heterogeneous Architectures. Adv. Funct. Mater. 2013, 23, 4423–4436. [Google Scholar] [CrossRef]
- Studart, A.R. Biologically Inspired Dynamic Material Systems. Angew. Chem. Int. Ed. 2015, 54, 3400–3416. [Google Scholar] [CrossRef]
- Liu, Z.; Meyers, M.A.; Zhang, Z.; Ritchie, R.O. Functional Gradients and Heterogeneities in Biological Materials: Design Principles, Functions, and Bioinspired Applications. Prog. Mater. Sci. 2017, 88, 467–498. [Google Scholar] [CrossRef]
- Svensson, L.; Oldberg, Å.; Heinegård, D. Collagen Binding Proteins. Osteoarthr. Cartil. 2001, 9, S23–S28. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.; Wang, N.; You, X.-G.; Zhang, A.-D.; Chen, X.-G.; Liu, Y. Collagen-Based Biocomposites Inspired by Bone Hierarchical Structures for Advanced Bone Regeneration: Ongoing Research and Perspectives. Biomater. Sci. 2022, 10, 318–353. [Google Scholar] [CrossRef] [PubMed]
- Cagini, C.; Riccitelli, F.; Messina, M.; Piccinelli, F.; Torroni, G.; Said, D.; Al Maazmi, A.; Dua, H.S. Epi-off-Lenticule-on Corneal Collagen Cross-Linking in Thin Keratoconic Corneas. Int. Ophthalmol. 2020, 40, 3403–3412. [Google Scholar] [CrossRef] [PubMed]
- Kong, B.; Sun, L.; Liu, R.; Chen, Y.; Shang, Y.; Tan, H.; Zhao, Y.; Sun, L. Recombinant Human Collagen Hydrogels with Hierarchically Ordered Microstructures for Corneal Stroma Regeneration. Chem. Eng. J. 2022, 428, 131012. [Google Scholar] [CrossRef]
- Wei, L.; Wang, D.; Liao, Z.; Gong, Z.; Zhao, W.; Gu, J.; Li, Y.; Li, J. Extraction of Keratin from Pig Nails and Electrospinning of Keratin/Nylon6 Nanofibers for Copper (II) Adsorption. Polymers 2023, 15, 467. [Google Scholar] [CrossRef]
- Huang, W.; Zaheri, A.; Yang, W.; Kisailus, D.; Ritchie, R.O.; Espinosa, H.; McKittrick, J. How Water Can Affect Keratin: Hydration-Driven Recovery of Bighorn Sheep (Ovis canadensis) Horns. Adv. Funct. Mater. 2019, 29, 1901077. [Google Scholar] [CrossRef]
- Marshall, R.C.; Gillespie, J. The Keratin Proteins of Wool, Horn and Hoof from Sheep. Aust. Jnl. Bio. Sci. 1977, 30, 389. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Forrest, R.; Li, S.; Wang, J.; Dyer, J.; Luo, Y.; Hickford, J. Wool Keratin-Associated Protein Genes in Sheep—A Review. Genes 2016, 7, 24. [Google Scholar] [CrossRef]
- Lee, J.N.; Lee, S.Y.; Park, W.H. Bioinspired Self-Healable Polyallylamine-Based Hydrogels for Wet Adhesion: Synergistic Contributions of Catechol-Amino Functionalities and Nanosilicate. ACS Appl. Mater. Interfaces 2021, 13, 18324–18337. [Google Scholar] [CrossRef] [PubMed]
- Harrington, M.J.; Jehle, F.; Priemel, T. Mussel Byssus Structure-Function and Fabrication as Inspiration for Biotechnological Production of Advanced Materials. Biotechnol. J. 2018, 13, 1800133. [Google Scholar] [CrossRef]
- Liu, Y.; Zheng, J.; Zhang, X.; Du, Y.; Li, K.; Yu, G.; Jia, Y.; Zhang, Y. Mussel-Inspired and Aromatic Disulfide-Mediated Polyurea-Urethane with Rapid Self-Healing Performance and Water-Resistance. J. Colloid Interface Sci. 2021, 593, 105–115. [Google Scholar] [CrossRef]
- Jing, X.; Mi, H.-Y.; Napiwocki, B.N.; Peng, X.-F.; Turng, L.-S. Mussel-Inspired Electroactive Chitosan/Graphene Oxide Composite Hydrogel with Rapid Self-Healing and Recovery Behavior for Tissue Engineering. Carbon 2017, 125, 557–570. [Google Scholar] [CrossRef]
- Krauss, S.; Metzger, T.H.; Fratzl, P.; Harrington, M.J. Self-Repair of a Biological Fiber Guided by an Ordered Elastic Framework. Biomacromolecules 2013, 14, 1520–1528. [Google Scholar] [CrossRef]
- Ahn, B.K.; Lee, D.W.; Israelachvili, J.N.; Waite, J.H. Surface-Initiated Self-Healing of Polymers in Aqueous Media. Nat. Mater. 2014, 13, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Shawkey, M.D.; Dhinojwala, A. Bioinspired Melanin-Based Optically Active Materials. Adv. Optical Mater. 2020, 8, 2000932. [Google Scholar] [CrossRef]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin Pigmentation in Mammalian Skin and Its Hormonal Regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef] [PubMed]
- Moses, D.N.; Mattoni, M.A.; Slack, N.L.; Waite, J.H.; Zok, F.W. Role of Melanin in Mechanical Properties of Glycera Jaws. Acta Biomater. 2006, 2, 521–530. [Google Scholar] [CrossRef]
- Hu, D.-N.; Simon, J.D.; Sarna, T. Role of Ocular Melanin in Ophthalmic Physiology and Pathology. Photochem. Photobiol. 2008, 84, 639–644. [Google Scholar] [CrossRef]
- Arao, T.; Perkins, E. The Skin of Primates. XLIII. Further Observations on the Philippine Tarsier (Tarsius syrichta). Am. J. Phys. Anthropol. 1969, 31, 93–96. [Google Scholar] [CrossRef]
- Wang, T.; Huang, W.; Pham, C.H.; Murata, S.; Herrera, S.; Kirchhofer, N.D.; Arkook, B.; Stekovic, D.; Itkis, M.E.; Goldman, N.; et al. Mesocrystalline Ordering and Phase Transformation of Iron Oxide Biominerals in the Ultrahard Teeth of Cryptochiton Stelleri. Small Struct. 2022, 3, 2100202. [Google Scholar] [CrossRef]
- Yaraghi, N.A.; Trikanad, A.A.; Restrepo, D.; Huang, W.; Rivera, J.; Herrera, S.; Zhernenkov, M.; Parkinson, D.Y.; Caldwell, R.L.; Zavattieri, P.D.; et al. The Stomatopod Telson: Convergent Evolution in the Development of a Biological Shield. Adv. Funct. Mater. 2019, 29, 1902238. [Google Scholar] [CrossRef]
- Velasco-Hogan, A.; Huang, W.; Serrano, C.; Kisailus, D.; Meyers, M.A. Tooth Structure, Mechanical Properties, and Diet Specialization of Piranha and Pacu (Serrasalmidae): A Comparative Study. Acta Biomater. 2021, 134, 531–545. [Google Scholar] [CrossRef]
- Kiakojouri, F.; De Biagi, V.; Abbracciavento, L. Design for Robustness: Bio-Inspired Perspectives in Structural Engi-neering. Biomimetics 2023, 8, 95. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.C.; Milliron, G.W.; Miserez, A.; Evans-Lutterodt, K.; Herrera, S.; Gallana, I.; Mershon, W.J.; Swanson, B.; Zavattieri, P.; DiMasi, E.; et al. The Stomatopod Dactyl Club: A Formidable Damage-Tolerant Biological Hammer. Science 2012, 336, 1275–1280. [Google Scholar] [CrossRef]
- Huang, W.; Zaheri, A.; Jung, J.-Y.; Espinosa, H.D.; Mckittrick, J. Hierarchical Structure and Compressive Deformation Mechanisms of Bighorn Sheep (Ovis canadensis) Horn. Acta Biomater. 2017, 64, 1–14. [Google Scholar] [CrossRef]
- Kitchener, A. An Analysis of the Forces of Fighting of the Blackbuck (Antilope cervicapra) and the Bighorn Sheep (Ovis canadensis) and the Mechanical Design of the Horn of Bovids. J. Zool. 1988, 214, 1–20. [Google Scholar] [CrossRef]
- Huang, W.; Montroni, D.; Wang, T.; Murata, S.; Arakaki, A.; Nemoto, M.; Kisailus, D. Nanoarchitected Tough Biological Composites from Assembled Chitinous Scaffolds. Acc. Chem. Res. 2022, 55, 1360–1371. [Google Scholar] [CrossRef]
- Beckham, G.T.; Crowley, M.F. Examination of the α-Chitin Structure and Decrystallization Thermodynamics at the Nanoscale. J. Phys. Chem. B 2011, 115, 4516–4522. [Google Scholar] [CrossRef]
- De Obaldia, E.E.; Jeong, C.; Grunenfelder, L.K.; Kisailus, D.; Zavattieri, P. Analysis of the Mechanical Response of Biomimetic Materials with Highly Oriented Microstructures through 3D Printing, Mechanical Testing and Modeling. J. Mech. Behav. Biomed. Mater. 2015, 48, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Qi, H.J.; Xie, T. Recent Progress in Shape Memory Polymer: New Behavior, Enabling Materials, and Mechanistic Understanding. Prog. Polym. Sci. 2015, 49–50, 79–120. [Google Scholar] [CrossRef]
- Meng, H.; Li, G. A Review of Stimuli-Responsive Shape Memory Polymer Composites. Polymer 2013, 54, 2199–2221. [Google Scholar] [CrossRef]
- Taylor, D.L.; In Het Panhuis, M. Self-Healing Hydrogels. Adv. Mater. 2016, 28, 9060–9093. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zuo, J. Self-Healing Polymers Based on Coordination Bonds. Adv. Mater. 2020, 32, 1903762. [Google Scholar] [CrossRef]
- Speck, O.; Speck, T. An Overview of Bioinspired and Biomimetic Self-Repairing Materials. Biomimetics 2019, 4, 26. [Google Scholar] [CrossRef]
- Lazaris, A.; Arcidiacono, S.; Huang, Y.; Zhou, J.-F.; Duguay, F.; Chretien, N.; Welsh, E.A.; Soares, J.W.; Karatzas, C.N. Spider Silk Fibers Spun from Soluble Recombinant Silk Produced in Mammalian Cells. Science 2002, 295, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Vollrath, F.; Knight, D.P. Liquid Crystalline Spinning of Spider Silk. Nature 2001, 410, 541–548. [Google Scholar] [CrossRef]
- Cranford, S.W.; Tarakanova, A.; Pugno, N.M.; Buehler, M.J. Nonlinear Material Behaviour of Spider Silk Yields Robust Webs. Nature 2012, 482, 72–76. [Google Scholar] [CrossRef]
- Agnarsson, I.; Kuntner, M.; Blackledge, T.A. Bioprospecting Finds the Toughest Biological Material: Extraordinary Silk from a Giant Riverine Orb Spider. PLoS ONE 2010, 5, e11234. [Google Scholar] [CrossRef]
- Yang, Q.; Li, G. Spider-Silk-like Shape Memory Polymer Fiber for Vibration Damping. Smart Mater. Struct. 2014, 23, 105032. [Google Scholar] [CrossRef]
- Huang, H.; Hu, J.; Zhu, Y. Shape-Memory Biopolymers Based on β-Sheet Structures of Polyalanine Segments Inspired by Spider Silks: Shape-Memory Biopolymers Based on β-Sheet Structures of Polyalanine Segments Inspired by Spider Silks. Macromol. Biosci. 2013, 13, 161–166. [Google Scholar] [CrossRef]
- Venkatesan, H.; Chen, J.; Liu, H.; Kim, Y.; Na, S.; Liu, W.; Hu, J. Artificial Spider Silk Is Smart like Natural One: Having Humidity-Sensitive Shape Memory with Superior Recovery Stress. Mater. Chem. Front. 2019, 3, 2472–2482. [Google Scholar] [CrossRef]
- Ryou, M.-H.; Kim, J.; Lee, I.; Kim, S.; Jeong, Y.K.; Hong, S.; Ryu, J.H.; Kim, T.-S.; Park, J.-K.; Lee, H.; et al. Mussel-Inspired Adhesive Binders for High-Performance Silicon Nanoparticle Anodes in Lithium-Ion Batteries. Adv. Mater. 2013, 25, 1571–1576. [Google Scholar] [CrossRef]
- Ahn, B.K. Perspectives on Mussel-Inspired Wet Adhesion. J. Am. Chem. Soc. 2017, 139, 10166–10171. [Google Scholar] [CrossRef] [PubMed]
- Work, A.; Lian, Y. A Critical Review of the Measurement of Ice Adhesion to Solid Substrates. Prog. Aerosp. Sci. 2018, 98, 1–26. [Google Scholar] [CrossRef]
- Maboudian, R. Critical Review: Adhesion in Surface Micromechanical Structures. J. Vac. Sci. Technol. B 1997, 15, 1. [Google Scholar] [CrossRef]
- Zheng, S.; Bawazir, M.; Dhall, A.; Kim, H.-E.; He, L.; Heo, J.; Hwang, G. Implication of Surface Properties, Bacterial Motility, and Hydrodynamic Conditions on Bacterial Surface Sensing and Their Initial Adhesion. Front. Bioeng. Biotechnol. 2021, 9, 643722. [Google Scholar] [CrossRef] [PubMed]
- Davis, G.D. Contamination of Surfaces: Origin, Detection and Effect on Adhesion. Surf. Interface Anal. 1993, 20, 368–372. [Google Scholar] [CrossRef]
- Barrett, D.G.; Fullenkamp, D.E.; He, L.; Holten-Andersen, N.; Lee, K.Y.C.; Messersmith, P.B. PH-Based Regulation of Hydrogel Mechanical Properties Through Mussel-Inspired Chemistry and Processing. Adv. Funct. Mater. 2013, 23, 1111–1119. [Google Scholar] [CrossRef] [PubMed]
- Burkett, J.R.; Wojtas, J.L.; Cloud, J.L.; Wilker, J.J. A Method for Measuring the Adhesion Strength of Marine Mussels. J. Adhes. 2009, 85, 601–615. [Google Scholar] [CrossRef]
- Das, S.; Miller, D.R.; Kaufman, Y.; Martinez Rodriguez, N.R.; Pallaoro, A.; Harrington, M.J.; Gylys, M.; Israelachvili, J.N.; Waite, J.H. Tough Coating Proteins: Subtle Sequence Variation Modulates Cohesion. Biomacromolecules 2015, 16, 1002–1008. [Google Scholar] [CrossRef]
- Desmond, K.W.; Zacchia, N.A.; Waite, J.H.; Valentine, M.T. Dynamics of Mussel Plaque Detachment. Soft Matter 2015, 11, 6832–6839. [Google Scholar] [CrossRef]
- Mian, S.A.; Yang, L.-M.; Saha, L.C.; Ahmed, E.; Ajmal, M.; Ganz, E. A Fundamental Understanding of Catechol and Water Adsorption on a Hydrophilic Silica Surface: Exploring the Underwater Adhesion Mechanism of Mussels on an Atomic Scale. Langmuir 2014, 30, 6906–6914. [Google Scholar] [CrossRef]
- Zhao, Q.; Lee, D.W.; Ahn, B.K.; Seo, S.; Kaufman, Y.; Israelachvili, J.N.; Waite, J.H. Underwater Contact Adhesion and Microarchitecture in Polyelectrolyte Complexes Actuated by Solvent Exchange. Nat. Mater. 2016, 15, 407–412. [Google Scholar] [CrossRef]
- Autumn, K.; Liang, Y.A.; Hsieh, S.T.; Zesch, W.; Chan, W.P.; Kenny, T.W.; Fearing, R.; Full, R.J. Adhesive Force of a Single Gecko Foot-Hair. Nature 2000, 405, 681–685. [Google Scholar] [CrossRef]
- Lee, H.; Lee, B.P.; Messersmith, P.B. A Reversible Wet/Dry Adhesive Inspired by Mussels and Geckos. Nature 2007, 448, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Du, J.; Wu, J.; Jiang, L. Superhydrophobic Gecko Feet with High Adhesive Forces towards Water and Their Bio-Inspired Materials. Nanoscale 2012, 4, 768–772. [Google Scholar] [CrossRef]
- Liu, K.; Jiang, L. Bio-Inspired Design of Multiscale Structures for Function Integration. Nano Today 2011, 6, 155–175. [Google Scholar] [CrossRef]
- Autumn, K. Gecko adhesion: Structure, function, and applications. MRS Bull. 2007, 32, 473–478. [Google Scholar] [CrossRef]
- Sethi, S.; Ge, L.; Ci, L.; Ajayan, P.M.; Dhinojwala, A. Gecko-Inspired Carbon Nanotube-Based Self-Cleaning Adhesives. Nano Lett. 2008, 8, 822–825. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Huang, J.; Chen, Z.; Chen, G.; Lai, Y. A Review on Special Wettability Textiles: Theoretical Models, Fabrication Technologies and Multifunctional Applications. J. Mater. Chem. A 2017, 5, 31–55. [Google Scholar] [CrossRef]
- Shen, Z.; Chen, F.; Zhu, X.; Yong, K.-T.; Gu, G. Stimuli-Responsive Functional Materials for Soft Robotics. J. Mater. Chem. B 2020, 8, 8972–8991. [Google Scholar] [CrossRef]
- Rus, D.; Tolley, M.T. Design, Fabrication and Control of Soft Robots. Nature 2015, 521, 467–475. [Google Scholar] [CrossRef]
- White, T.J.; Broer, D.J. Programmable and Adaptive Mechanics with Liquid Crystal Polymer Networks and Elastomers. Nat. Mater. 2015, 14, 1087–1098. [Google Scholar] [CrossRef]
- Rich, S.I.; Wood, R.J.; Majidi, C. Untethered Soft Robotics. Nat. Electron. 2018, 1, 102–112. [Google Scholar] [CrossRef]
- Liao, H.; Liao, C.; Blamires, S.J.; Tso, I. Multifunctionality of an Arthropod Predator’s Body Coloration. Funct. Ecol. 2019, 33, 1067–1075. [Google Scholar] [CrossRef]
- Yoshioka, S.; Nakano, T.; Nozue, Y.; Kinoshita, S. Coloration Using Higher Order Optical Interference in the Wing Pattern of the Madagascan Sunset Moth. J. R. Soc. Interface 2008, 5, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Stavenga, D.G.; Leertouwer, H.L.; Wilts, B.D. Colouration Principles of Nymphaline Butterflies—Thin Films, Melanin, Ommochromes and Wing Scale Stacking. J. Exp. Biol. 2014, 217, 2171–2180. [Google Scholar] [CrossRef] [PubMed]
- Cuthill, I.C.; Stevens, M.; Sheppard, J.; Maddocks, T.; Párraga, C.A.; Troscianko, T.S. Disruptive Coloration and Background Pattern Matching. Nature 2005, 434, 72–74. [Google Scholar] [CrossRef]
- Ziai, Y.; Petronella, F.; Rinoldi, C.; Nakielski, P.; Zakrzewska, A.; Kowalewski, T.A.; Augustyniak, W.; Li, X.; Calogero, A.; Sabała, I.; et al. Chameleon-Inspired Multifunctional Plasmonic Nanoplatforms for Biosensing Applications. NPG Asia Mater 2022, 14, 18. [Google Scholar] [CrossRef]
- Vatankhah-Varnosfaderani, M.; Keith, A.N.; Cong, Y.; Liang, H.; Rosenthal, M.; Sztucki, M.; Clair, C.; Magonov, S.; Ivanov, D.A.; Dobrynin, A.V.; et al. Chameleon-like Elastomers with Molecularly Encoded Strain-Adaptive Stiffening and Coloration. Science 2018, 359, 1509–1513. [Google Scholar] [CrossRef]
- Bu, X.; Bai, H. Recent Progress of Bio-Inspired Camouflage Materials: From Visible to Infrared Range. Chem. Res. Chin. Univ. 2023, 39, 19–29. [Google Scholar] [CrossRef]
- Berg, P.; Berg, J.; Berg, R. Predator–Prey Interaction between a Boomslang, Dispholidus Typus, and a Flap-necked Chameleon, Chamaeleo Dilepis. Afr. J. Ecol. 2020, 58, 855–859. [Google Scholar] [CrossRef]
- Huss, J.C.; Schoeppler, V.; Merritt, D.J.; Best, C.; Maire, E.; Adrien, J.; Spaeker, O.; Janssen, N.; Gladisch, J.; Gierlinger, N.; et al. Climate-Dependent Heat-Triggered Opening Mechanism of Banksia Seed Pods. Adv. Sci. 2018, 5, 1700572. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Timonen, J.V.I.; Carlson, A.; Drotlef, D.-M.; Zhang, C.T.; Kolle, S.; Grinthal, A.; Wong, T.-S.; Hatton, B.; Kang, S.H.; et al. Multifunctional Ferrofluid-Infused Surfaces with Reconfigurable Multiscale Topography. Nature 2018, 559, 77–82. [Google Scholar] [CrossRef]
- Hu, W.; Lum, G.Z.; Mastrangeli, M.; Sitti, M. Small-Scale Soft-Bodied Robot with Multimodal Locomotion. Nature 2018, 554, 81–85. [Google Scholar] [CrossRef]
- Huss, J.C.; Spaeker, O.; Gierlinger, N.; Merritt, D.J.; Miller, B.P.; Neinhuis, C.; Fratzl, P.; Eder, M. Temperature-Induced Self-Sealing Capability of Banksia Follicles. J. R. Soc. Interface 2018, 15, 20180190. [Google Scholar] [CrossRef]
- Huang, W.; Wang, T.; Perez-Fernandez, C.; DiRuggiero, J.; Kisailus, D. Iron Acquisition and Mineral Transformation by Cyanobacteria Living in Extreme Environments. Mater. Today Bio 2022, 17, 100493. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Hogan, A.; Deheyn, D.D.; Koch, M.; Nothdurft, B.; Arzt, E.; Meyers, M.A. On the Nature of the Transparent Teeth of the Deep-Sea Dragonfish, Aristostomias Scintillans. Matter 2019, 1, 235–249. [Google Scholar] [CrossRef]
- Munch, E.; Launey, M.E.; Alsem, D.H.; Saiz, E.; Tomsia, A.P.; Ritchie, R.O. Tough, Bio-Inspired Hybrid Materials. Science 2008, 322, 1516–1520. [Google Scholar] [CrossRef]
- Deville, S.; Saiz, E.; Nalla, R.K.; Tomsia, A.P. Freezing as a Path to Build Complex Composites. Science 2006, 311, 515–518. [Google Scholar] [CrossRef]
- Liu, Q.; Ye, F.; Gao, Y.; Liu, S.; Yang, H.; Zhou, Z. Fabrication of a New SiC/2024Al Co-Continuous Composite with Lamellar Microstructure and High Mechanical Properties. J. Alloy. Compd. 2014, 585, 146–153. [Google Scholar] [CrossRef]
- Zhang, X.; Meng, Q.; Zhang, K.; Zhu, R.; Qu, Z.; Li, Y.; He, R. 3D-Printed Bioinspired Al2O3/Polyurea Dual-Phase Architecture with High Robustness, Energy Absorption, and Cyclic Life. Chem. Eng. J. 2023, 463, 142378. [Google Scholar] [CrossRef]
- Wang, J.; Hu, D.; Zhang, Z.; Pei, B.; Xu, R.; Wu, X. Anti-Impact Performance of Bionic Tortoiseshell-like Composites. Compos. Struct. 2023, 303, 116315. [Google Scholar] [CrossRef]
- Katz, Z.; Yazdani Sarvestani, H.; Gholipour, J.; Ashrafi, B. Bioinspired Hierarchical Ceramic Sutures for Multi-Modal Performance. Adv. Mater. Inter. 2023, 10, 2300098. [Google Scholar] [CrossRef]
- Wickramasinghe, S.; Al-Ketan, O.; Peng, C.; Tee, Y.L.; Kajtaz, M.; Tran, P. Influence of Design Parameters on the Flexural Properties of a Bio-Inspired Suture Structure. Virtual Phys. Prototyp. 2023, 18, e2204845. [Google Scholar] [CrossRef]
- Cui, Y.; Li, D.; Gong, C.; Chang, C. Bioinspired Shape Memory Hydrogel Artificial Muscles Driven by Solvents. ACS Nano 2021, 15, 13712–13720. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Bae, J.; Jeon, H.; Kim, T. Spider-Inspired Regenerated Silk Fibroin Fiber Actuator via Microfluidic Spinning. Chem. Eng. J. 2022, 444, 136556. [Google Scholar] [CrossRef]
- Greco, G.; Arndt, T.; Schmuck, B.; Francis, J.; Bäcklund, F.G.; Shilkova, O.; Barth, A.; Gonska, N.; Seisenbaeva, G.; Kessler, V.; et al. Tyrosine Residues Mediate Supercontraction in Biomimetic Spider Silk. Commun. Mater. 2021, 2, 43. [Google Scholar] [CrossRef]
- Kim, H.; Jang, Y.; Lee, D.Y.; Moon, J.H.; Choi, J.G.; Spinks, G.M.; Gambhir, S.; Officer, D.L.; Wallace, G.G.; Kim, S.J. Bio-Inspired Stretchable and Contractible Tough Fiber by the Hybridization of GO/MWNT/Polyurethane. ACS Appl. Mater. Interfaces 2019, 11, 31162–31168. [Google Scholar] [CrossRef] [PubMed]
- Shuai, L.; Guo, Z.H.; Zhang, P.; Wan, J.; Pu, X.; Wang, Z.L. Stretchable, Self-Healing, Conductive Hydrogel Fibers for Strain Sensing and Triboelectric Energy-Harvesting Smart Textiles. Nano Energy 2020, 78, 105389. [Google Scholar] [CrossRef]
- Wu, Y.; Shah, D.U.; Wang, B.; Liu, J.; Ren, X.; Ramage, M.H.; Scherman, O.A. Biomimetic Supramolecular Fibers Exhibit Water-Induced Supercontraction. Adv. Mater. 2018, 30, 1707169. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.Q.; Shafiq, M.; Li, J.; Yu, K.; Liu, Z.; Zhou, X.; Zhu, M. Recent Developments in Artificial Spider Silk and Functional Gel Fibers. SmartMat 2023, e1189. [Google Scholar] [CrossRef]
- Zhang, X.; Cui, M.; Wang, S.; Han, F.; Xu, P.; Teng, L.; Zhao, H.; Wang, P.; Yue, G.; Zhao, Y.; et al. Extensible and Self-Recoverable Proteinaceous Materials Derived from Scallop Byssal Thread. Nat. Commun. 2022, 13, 2731. [Google Scholar] [CrossRef]
- Mehdizadeh, M.; Weng, H.; Gyawali, D.; Tang, L.; Yang, J. Injectable Citrate-Based Mussel-Inspired Tissue Bioadhesives with High Wet Strength for Sutureless Wound Closure. Biomaterials 2012, 33, 7972–7983. [Google Scholar] [CrossRef]
- Wang, R.; Li, J.; Chen, W.; Xu, T.; Yun, S.; Xu, Z.; Xu, Z.; Sato, T.; Chi, B.; Xu, H. A Biomimetic Mussel-Inspired ε-Poly- L -Lysine Hydrogel with Robust Tissue-Anchor and Anti-Infection Capacity. Adv. Funct. Mater. 2017, 27, 1604894. [Google Scholar] [CrossRef]
- Ma, Y.; Ma, S.; Wu, Y.; Pei, X.; Gorb, S.N.; Wang, Z.; Liu, W.; Zhou, F. Remote Control over Underwater Dynamic Attachment/Detachment and Locomotion. Adv. Mater. 2018, 30, 1801595. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Pei, X.; Duan, L.; Zhu, Z.; Liu, Y.; Chen, J.; Chen, T.; Ji, P.; Wan, Q.; Wang, J. A Mussel-Inspired Film for Adhesion to Wet Buccal Tissue and Efficient Buccal Drug Delivery. Nat. Commun. 2021, 12, 1689. [Google Scholar] [CrossRef]
- Nie, X.; Wu, S.; Lv, P.; Ke, H.; Huang, F.; Wei, Q. Chameleon-Inspired Iridescent Structural Color Textiles with Reversible Multiple Stimulus-Responsive Functions. Chem. Eng. J. 2022, 433, 134410. [Google Scholar] [CrossRef]
- Zhang, Y.-S.; Jiang, S.-A.; Lin, J.-D.; Lee, C.-R. Bio-Inspired Design of Active Photo-Mechano-Chemically Dual-Responsive Photonic Film Based on Cholesteric Liquid Crystal Elastomers. J. Mater. Chem. C 2020, 8, 5517–5524. [Google Scholar] [CrossRef]
- Dong, Y.; Bazrafshan, A.; Pokutta, A.; Sulejmani, F.; Sun, W.; Combs, J.D.; Clarke, K.C.; Salaita, K. Chameleon-Inspired Strain-Accommodating Smart Skin. ACS Nano 2019, 13, 9918–9926. [Google Scholar] [CrossRef]
- Chou, H.-H.; Nguyen, A.; Chortos, A.; To, J.W.F.; Lu, C.; Mei, J.; Kurosawa, T.; Bae, W.-G.; Tok, J.B.-H.; Bao, Z. A Chameleon-Inspired Stretchable Electronic Skin with Interactive Colour Changing Controlled by Tactile Sensing. Nat. Commun. 2015, 6, 8011. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Choi, J.; Kim, K.K.; Won, P.; Hong, S.; Ko, S.H. Biomimetic Chameleon Soft Robot with Artificial Crypsis and Disruptive Coloration Skin. Nat. Commun. 2021, 12, 4658. [Google Scholar] [CrossRef] [PubMed]
- Boutin, R.; Munnier, E.; Renaudeau, N.; Girardot, M.; Pinault, M.; Chevalier, S.; Chourpa, I.; Clément-Larosière, B.; Imbert, C.; Boudesocque-Delaye, L. Spirulina Platensis Sustainable Lipid Extracts in Alginate-Based Nanocarriers: An Algal Approach against Biofilms. Algal Res. 2019, 37, 160–168. [Google Scholar] [CrossRef]
- Bernal, O.I.; Bharti, B.; Flickinger, M.C.; Velev, O.D. Fabrication of Photoreactive Biocomposite Coatings via Electric Field-Assisted Assembly of Cyanobacteria. Langmuir 2017, 33, 5304–5313. [Google Scholar] [CrossRef]
- Ritchie, R.O. The Conflicts between Strength and Toughness. Nat. Mater. 2011, 10, 817–822. [Google Scholar] [CrossRef]
- Tadepalli, S.; Slocik, J.M.; Gupta, M.K.; Naik, R.R.; Singamaneni, S. Bio-Optics and Bio-Inspired Optical Materials. Chem. Rev. 2017, 117, 12705–12763. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.X.; Chen, C.-T.; Richmond, D.J.; Buehler, M.J. Bioinspired Hierarchical Composite Design Using Machine Learning: Simulation, Additive Manufacturing, and Experiment. Mater. Horiz. 2018, 5, 939–945. [Google Scholar] [CrossRef]
- Liu, Z.; Shao, C.; Jin, B.; Zhang, Z.; Zhao, Y.; Xu, X.; Tang, R. Crosslinking Ionic Oligomers as Conformable Precursors to Calcium Carbonate. Nature 2019, 574, 394–398. [Google Scholar] [CrossRef]
- Fang, W.; Mu, Z.; He, Y.; Kong, K.; Jiang, K.; Tang, R.; Liu, Z. Organic–Inorganic Covalent–Ionic Molecules for Elastic Ceramic Plastic. Nature 2023, 1–7. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Main Functions | Representative Organisms | Main Structure and Mechanisms | Bioinspired Applications | References |
|---|---|---|---|---|
| Impact, crush and wear resistance | Ironclad beetle, Chiton and mantis shrimp | Helicoidal fibers, interdigitated sutures, lamellae | Structural components, cutting tools, armors | [114,115,116,117,118,119,120,121] |
| Self-healing and shape memory | Spider, mussel | Fibers, reversible bonds, elastic network | Artificial skin, artificial muscle | [122,123,124,125,126,127,128] |
| Adhesion and anti-fouling | Gecko feet, mussel, lotus leaf | Pillar arrays, dopamine | Medical tapes, robotic arms, coatings | [90,129,130,131,132] |
| Sensing, actuating, and camouflage | Moths, chameleons, and banksia plants | Hierarchical structure, photonic crystal | Artificial skin, military applications, soft robots | [133,134,135,136,137] |
| Strategies of living in extreme environment | Cyanobacteria | Biofilms, metabolism, photosynthesis | Living materials, space exploration, and colonization | [138,139] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, J.; Huang, W.; Zhou, H. Multifunctionality in Nature: Structure–Function Relationships in Biological Materials. Biomimetics 2023, 8, 284. https://doi.org/10.3390/biomimetics8030284
Zhong J, Huang W, Zhou H. Multifunctionality in Nature: Structure–Function Relationships in Biological Materials. Biomimetics. 2023; 8(3):284. https://doi.org/10.3390/biomimetics8030284
Chicago/Turabian StyleZhong, Jiaming, Wei Huang, and Huamin Zhou. 2023. "Multifunctionality in Nature: Structure–Function Relationships in Biological Materials" Biomimetics 8, no. 3: 284. https://doi.org/10.3390/biomimetics8030284
APA StyleZhong, J., Huang, W., & Zhou, H. (2023). Multifunctionality in Nature: Structure–Function Relationships in Biological Materials. Biomimetics, 8(3), 284. https://doi.org/10.3390/biomimetics8030284