2.1. Shunting Inhibition
Based on a detailed kinetic model of synaptic transmission [
31], a phenomenological model of the postsynaptic current in a neuronal cell can be described as follows:
where
,
, and
are the postsynaptic membrane potential, synaptic conductance, and synaptic reversal potential, respectively. This description is phenomenological, and in the relevant voltage ranges, the induced synaptic current exhibits an approximately linear dependence on the difference between
and
. In the simplest models, the time-dependent synaptic conductance
(t) has a bi-exponential profile similar to that of an alpha function. From Equation (1), it can be seen that the polarity of the synaptic current induced in the neuronal cells depends not only on the synaptic receptor involved (that fixes
), but also on the instantaneous
. Synapses with their
significantly higher (lower) than the resting
have excitatory (inhibitory) effects; upon activation, they depolarize (hyperpolarize) the postsynaptic neuronal cell membrane. Synapses with their
close to the resting
are called shunting inhibitory synapses. Most contemporary low-power analog synapse circuits [
26,
32] implement a current-based synapse model that mimics the bi-exponential profile of the synaptic current, but ignores its dependence on the difference between the instantaneous
and
. As is clear from Equation (1), the circuit implementation requires a resistor-like element between
and
.
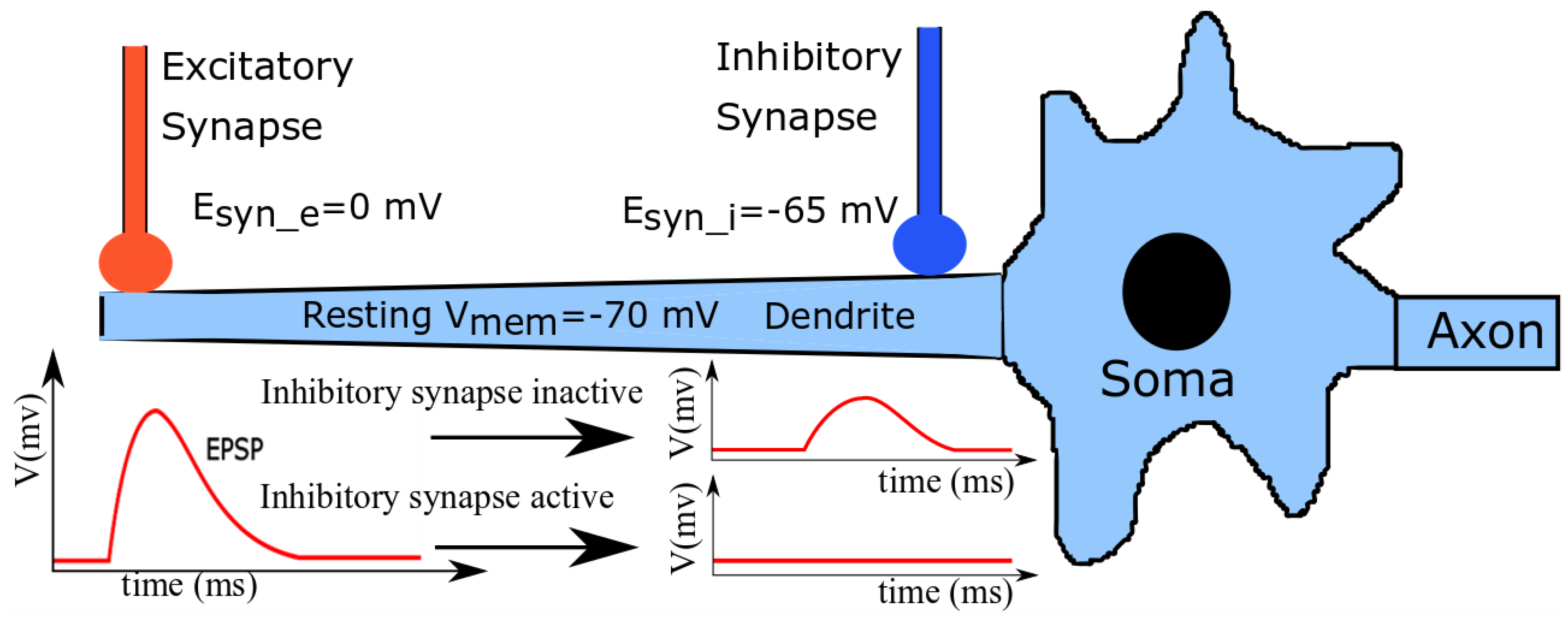
A simplified schematic illustration of a neuronal cell is shown in
Figure 1. A group of excitatory and shunting inhibitory synapses distal and proximal to the soma impinge on a dendritic branch. In line with generally measured neurophysiological values [
33], the resting
Vmem of the cell, synaptic reversal potential of the excitatory synapses (
), and that of shunting inhibitory synapses (
) are approximately −70 mV, 0 mV (typical for α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) synapses), and −65 mV (typical for GABAa (γ-aminobutyric acid type A) synapses), respectively. Upon the standalone activation of the excitatory synapses, the induced current depolarizes the local membrane and drives its potential towards
. This disturbance travels as a gradually attenuating excitatory postsynaptic potential (EPSP) towards the soma and causes it to spike (if the depolarizing current is strong enough). The standalone activation of the shunting inhibitory synapses has a negligible depolarization effect on the neuronal membrane, but increases the local membrane conductance around the soma (as
is close to
). However, when both excitatory and shunting inhibitory synapses are activated together, the EPSPs generated by the excitatory synapses are attenuated by the shunting inhibitory synapses when the traveling EPSPs reach near the soma. If the inhibition is strong enough, the depolarization induced by the excitatory synapses is completely silenced and the soma’s membrane potential remains undisturbed; this phenomenon is called shunting inhibition.
The relative spatial placements and temporal activations of the shunting inhibitory and excitatory synapses on dendritic arbors are believed to play a significant role in the computational capabilities of neuronal cells. In previous studies [
19,
33], it was shown that shunting inhibition implements an approximate multiplication between the excitatory and inhibitory synaptic conductance within the dendritic tree. In another study [
20], it was demonstrated that in a neuronal cell, the shunting inhibitory synapses can modify the number of electrically isolated dendritic compartments; these then act as independent compartments for detecting the coincidence of incoming spikes. A study discussing the escape behavior of crayfish [
18] hypothesized and then experimentally confirmed how the relative positioning and activation of excitatory and shunting inhibitory synapses on a neuronal cell can help fish decide whether to initiate an escape response or continue usual feeding activity in response to a potentially dangerous situation. Thus, implementing the shunting inhibitory synapse circuits is necessary to replicate the electrical behavior of the neuronal circuits in the brain. Moreover, given their potential to enhance neuronal computation via specific non-linear interactions with excitatory inputs, it is evident that low-power shunting inhibitory synapse circuits can add to the capabilities of neuromorphic chips implementing multi-compartment or reduced-compartment neuron models.
2.2. Conductance-Based Synapse Circuit
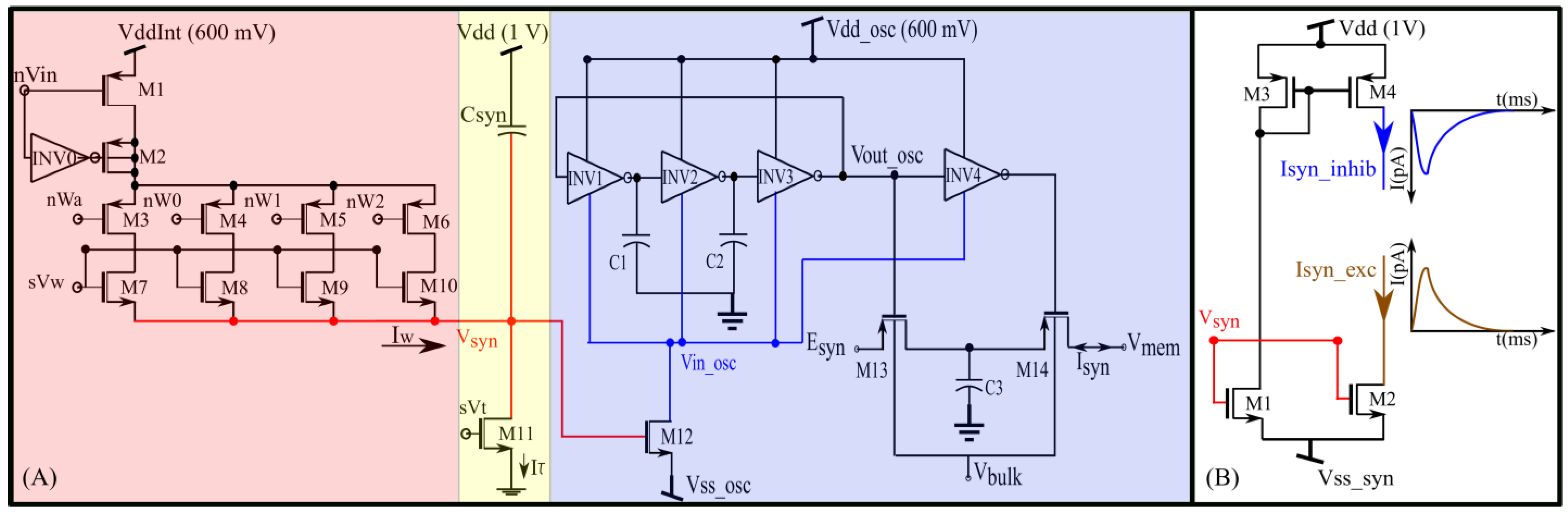
A schematic of the proposed synapse circuit is shown in
Figure 2A. It has three stages: a digital-to-analog converter (DAC, M1–M10), an integrator stage (
and M11) similar to the log domain integrator (LDI), and a transconductance stage (M12–M14, C1–C3, and INV1–INV4). The DAC and integrator stages are similar to the synapse circuit proposed in our previous study [
27,
34].
In the DAC, M7–M10 are binary-weighted transistors. The bias voltage
controls the strength of the synaptic current, and the switches M3–M6 configure the four-bit synaptic efficacy. The efficacy is stored in digital memories updated by a learning circuitry. Upon the application of an input pulse (~2 ms wide) at the gate of M1, depending on the value of
and the synaptic efficacy, the DAC sources a current into the node V
syn and charges it for the duration of the input pulse. The inverter INV0 along with M2 is for reducing the charge injection effect. Once the input pulse is turned off, V
syn is discharged linearly by a constant current sunk by transistor M11, which operates in the saturation region (for V
syn > 4 U
T). The bias voltage
and capacitance
control the discharge rate. In most contemporary current-based synapse circuits, the node V
syn activates a MOS transistor (M2 in
Figure 2B) for converting the linear voltage V
syn to an exponential current (I
syn_exc), as shown in
Figure 2B. In the proposed circuit, V
syn activates the transconductance stage.
The transconductance stage is designed using an unbalanced switched-capacitor-like circuit (INV4, M13, M14, and C3) activated by a ring oscillator-type circuit (INV1–3). In a typical ring oscillator, the source terminals of the PMOS and NMOS devices of the inverters (
and node
are connected to constant voltage sources, and the circuit generates a pulsed waveform with the maximum and minimum values of
and
, respectively. The propagation time of the inverters determines the pulse width and frequency of the waveform. It is calculated as the average time taken by the inverter’s PMOS transistor to charge its output capacitance and that taken by the NMOS transistor to discharge the same capacitance. To derive this, we consider the inverter INV1 in
Figure 2A. When its gate voltage
is near
its NMOS transistor discharges its output node, as follows:
where
and
are the current scaling factor and capacitive coupling ratio of the inverter’s NMOS transistor, respectively.
is the thermal voltage.
is the output node of INV1. The body effect is ignored in the calculations. Separating the variables in Equation (2) and integrating yields as follows:
By choosing the halfway point between
and
to calculate the discharging time (
, the integral limit on the left-hand side ranges from
to
and that on the right-hand side ranges from 0 to
Solving Equation (3) for
yields as follows:
By repeating the same derivation for the charging process via the inverter’s PMOS transistor, the charging time is given as follows:
where
and
are the current scaling factor and capacitive coupling ratio of the PMOS transistor in the inverter, respectively. In Equation (4),
is calculated when
and
. Similarly, in Equation (5),
is calculated when
and
. To simplify the equation, we assume that
=
. Based on these substitutions, and considering that
, the propagation time of the inverter (the average of
and
), is given as follows:
In the proposed circuit, only
is a constant-voltage source (600 mV). The node
is not a voltage source. The current sourced and sunk by the inverters (INV1–4) and M12 determine its voltage. The propagation time (
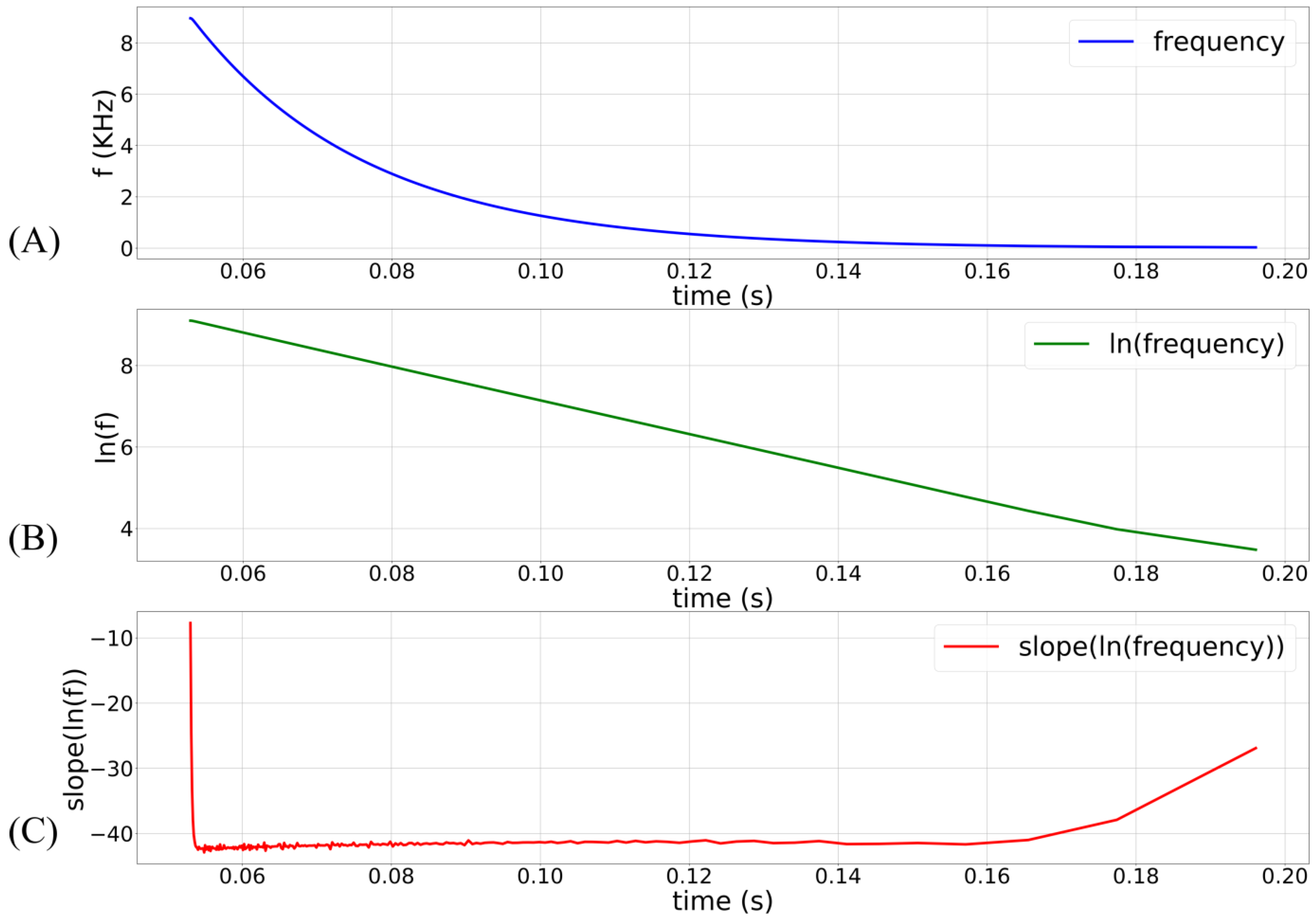
) of the inverters (which controls the oscillator’s frequency and pulse width) is thus not constant, and the frequency has an exponential dependence on
(see
Figure 3). The terminal
is kept above 0 V (~35 mV) to minimize the leakage current via M12. This renders the oscillator circuit inactive when there is no input pulse. In this inactive state, V
syn and
are close to 0 V and
, respectively. The oscillator remains off because there is insufficient headroom for oscillation. In response to a pulse input to the DAC stage, the linearly charging and discharging V
syn activates M12 that sinks current out of
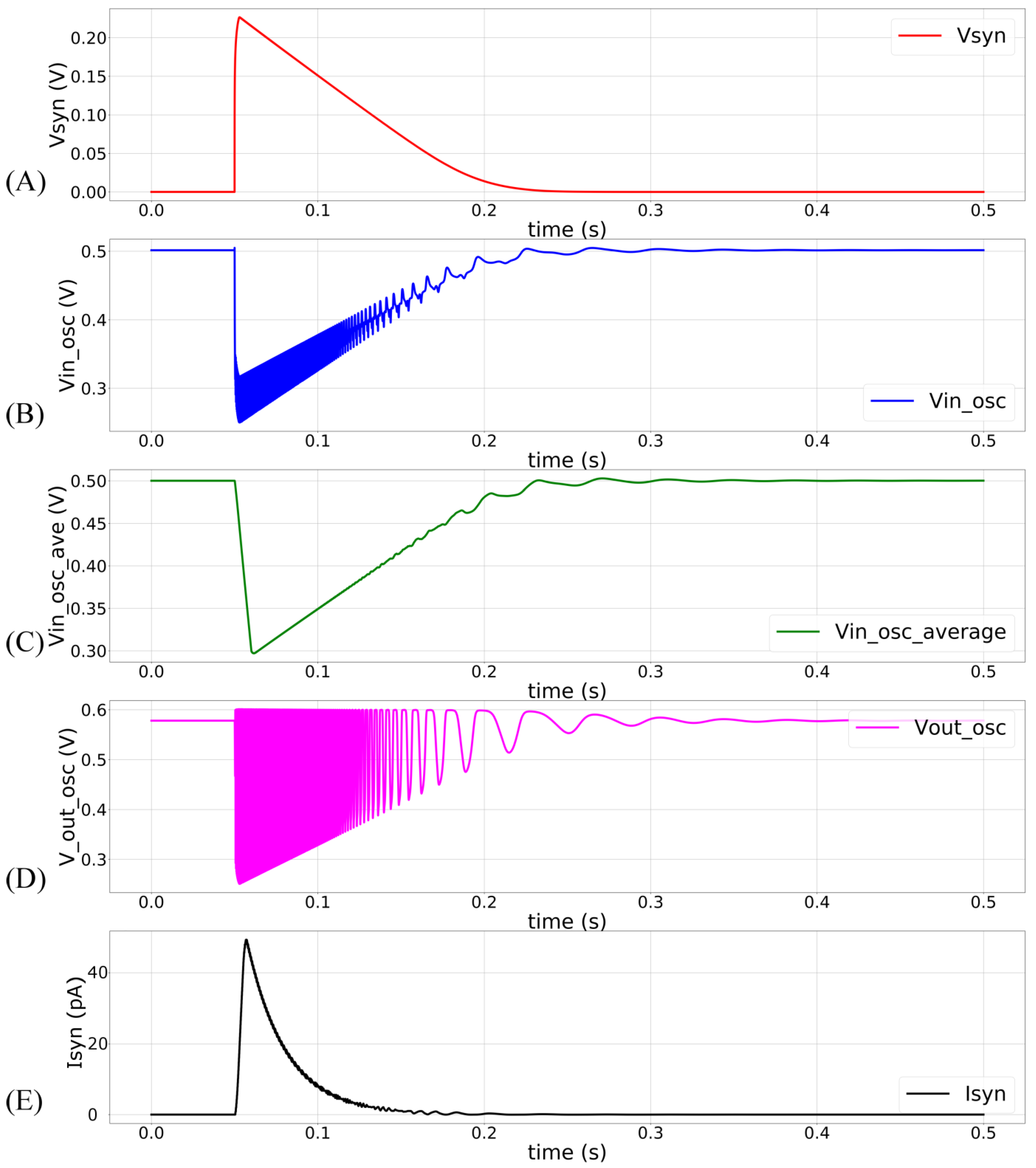
, pulling it down and activating the oscillator. The profiles of V
syn and
upon circuit activation at 50 ms (obtained via Spectre simulation) are shown in
Figure 4A,B. The voltage
is approximately linearly related to V
syn, as indicated by the moving average of
(
Figure 4C). The oscillator’s output (V
out_osc) is shown in
Figure 4D. The oscillator’s output V
out_osc activates the switched-capacitor-like circuit (M13, M14, INV4, and C3) that implements an asymmetric resistor-type element between
and
. Here,
is fixed at 600 mV. When inactive, the gates of M13 and M14 remain close to
and a very small current flows out of
(if
). Upon circuit activation, M13 and M14 receive out-of-phase pulses (via INV4) whose amplitudes decrease from
to
. These pulses activate M13 and M14 in the subthreshold domain (where the drain current of the MOS device is exponentially related to its gate voltage). As the amplitude of these pulses decreases linearly over time (because
increases linearly over time), an exponential current is induced out of
(for
).
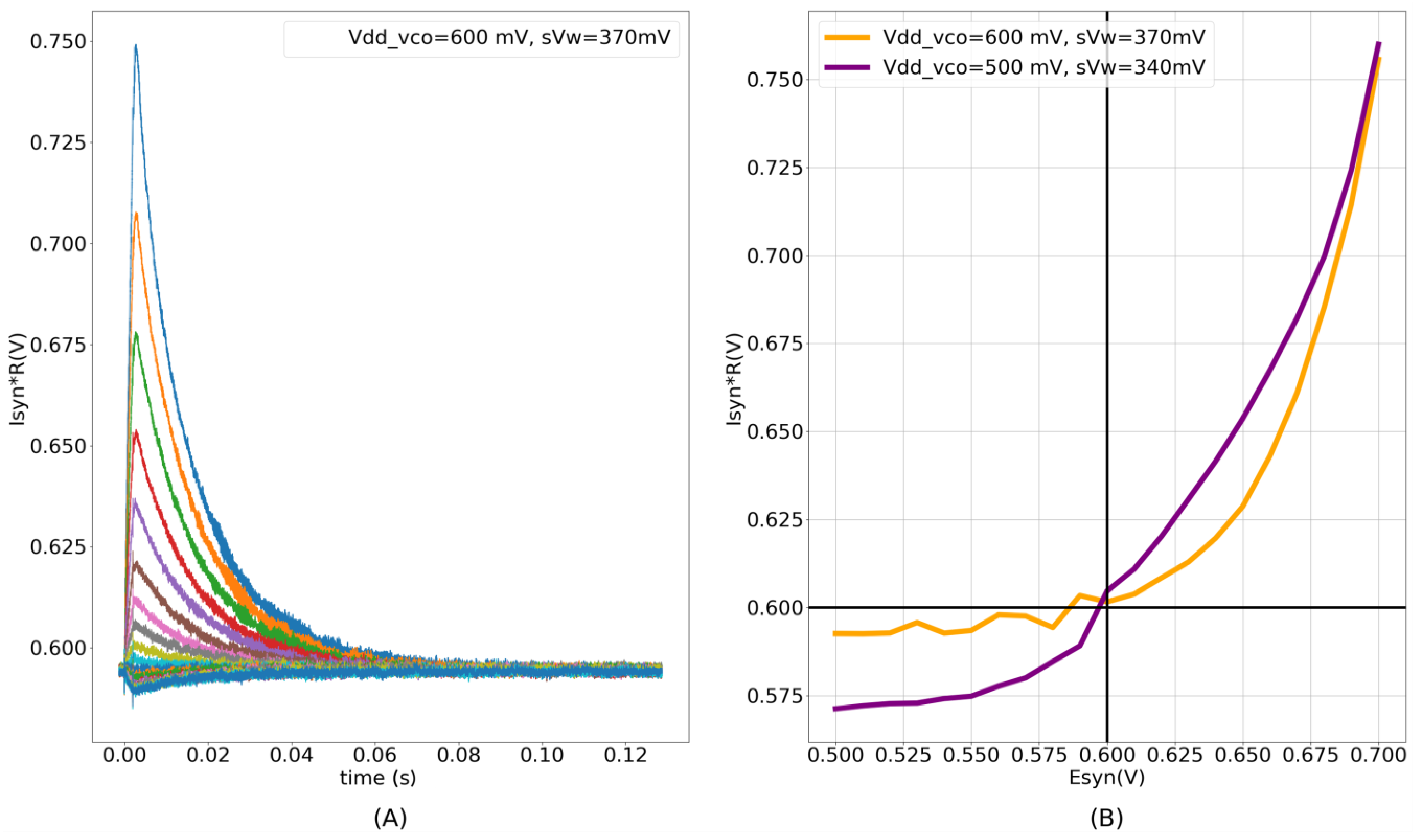
Figure 4E plots the moving average profile of the induced synaptic current for
700 mV and
600 mV.
This circuit functions as a non-linear resistor. Unlike an ideal resistor, the induced current has an exponential dependence on the difference between
and
. The current in a PMOS device increases with its source-gate overdrive voltage, and for a fixed
, the overdrive in M13 and M14 is higher for
than for
. Owing to the exponential current–voltage (I–V) relationship in the subthreshold domain, the resistance emulated is exponentially larger for values of
<
(in comparison with values of
>
), leading to an “asymmetric” I–V relationship (See
Section 3.1). The transconductance stage of the proposed circuit can also be used as a non-linear resistor between terminals
and
if the node V
syn is fixed at a constant value.
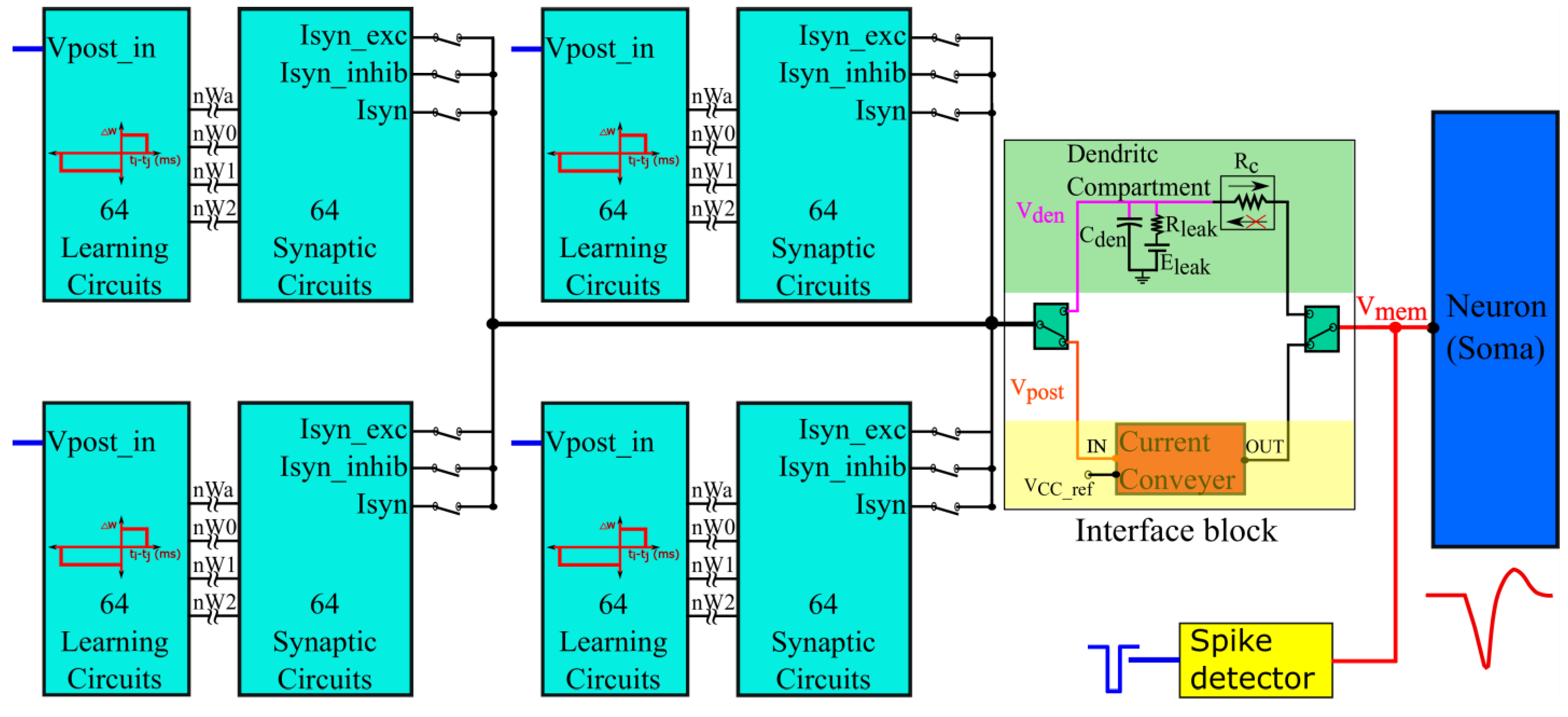
2.3. Architecture of Silicon Neuron Circuit
The synapse circuit described in
Section 2.2 is incorporated into a silicon neuron circuit fabricated in the Taiwan Semiconductor Manufacturing Company (TSMC) 250 nm technology node. The block diagram is shown in
Figure 5. It has 256 synapse circuits in groups of four (64 circuits per group) for activating a qualitatively modeled soma circuit [
16,
35]. The polarities of the synapse circuits can be configured as a group. In an excitatory or inhibitory configuration (I
syn_exc or I
syn_inhib terminals chosen as outputs in
Figure 2 and
Figure 5), the current-based synapse model is evoked. Here, the induced synaptic current does not depend on the postsynaptic potential, whereas it does in the conductance-based configuration (terminal I
syn is chosen as the output in
Figure 2 and
Figure 5). Upon activation, the synapse circuits induce a current into the soma circuit via an interface circuit, causing it to either depolarize or hyperpolarize. The spiking behavior and current polarity of the neuronal soma circuit are opposite to the convention. An excitatory (inhibitory) synapse circuit has
lower (higher) than the resting
and depolarizes (hyperpolarizes) the soma by sinking (sourcing) current out of (into) it, causing the postsynaptic membrane potential to drop (rise) below (above) its resting value. This is because the soma circuit is designed primarily using PMOS transistors with much smaller leakage currents than their NMOS counterparts. This minimizes the power consumption of the circuit. In this study, the soma circuit is configured in the fast-spiking Class 1 mode of Hodgkin’s classification (no spike-frequency adaptation). Its spikes are converted into pulses using a spike detector circuit (see [
30] for details). Subsequently, these pulses are fed back to the learning circuitry (representing the postsynaptic spike, V
post_in in
Figure 5). All synapse circuits have a learning circuitry to implement adaptive spike-timing-dependent plasticity (STDP) learning, which updates the synaptic efficacy based on the spike timings of the pre- and postsynaptic spikes. To perform pattern detection on-chip, input spike trains are transmitted from a PC to the chip via a field-programmable gate array. An on-chip spike address decoder circuit is used to activate the synapse circuits. The details of this spike transfer module can be found in a previous study [
30].
2.3.1. Interface Block
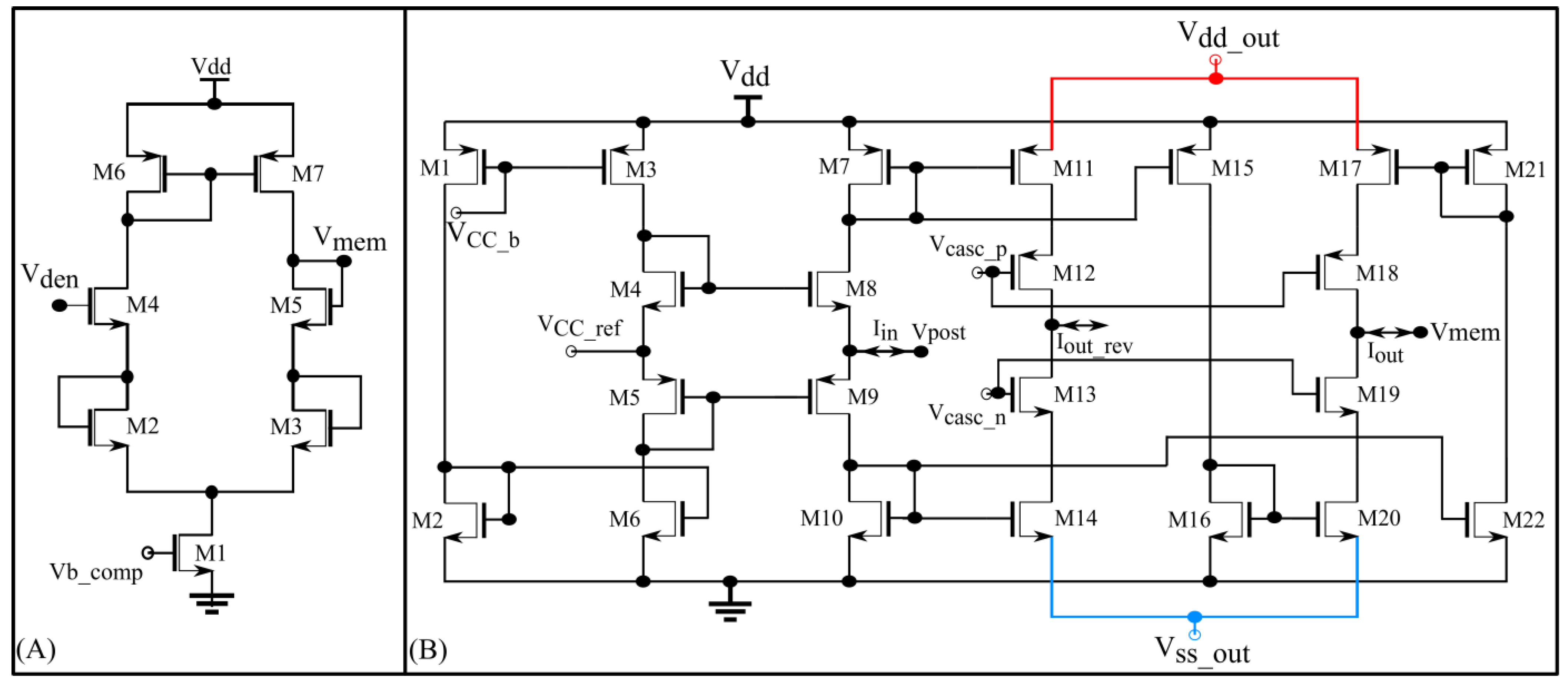
The interface block has two circuits available as a link between the synaptic and soma circuits: a unidirectional resistor (green part in
Figure 5) composed of a transconductance circuit (
Figure 6A), and a bidirectional current conveyor circuit (yellow part in
Figure 5) whose circuitry is shown in
Figure 6B. The former configures the neuron as a unidirectional two-compartment neuron circuit, whereas the latter does so as a single-compartment point neuron circuit.
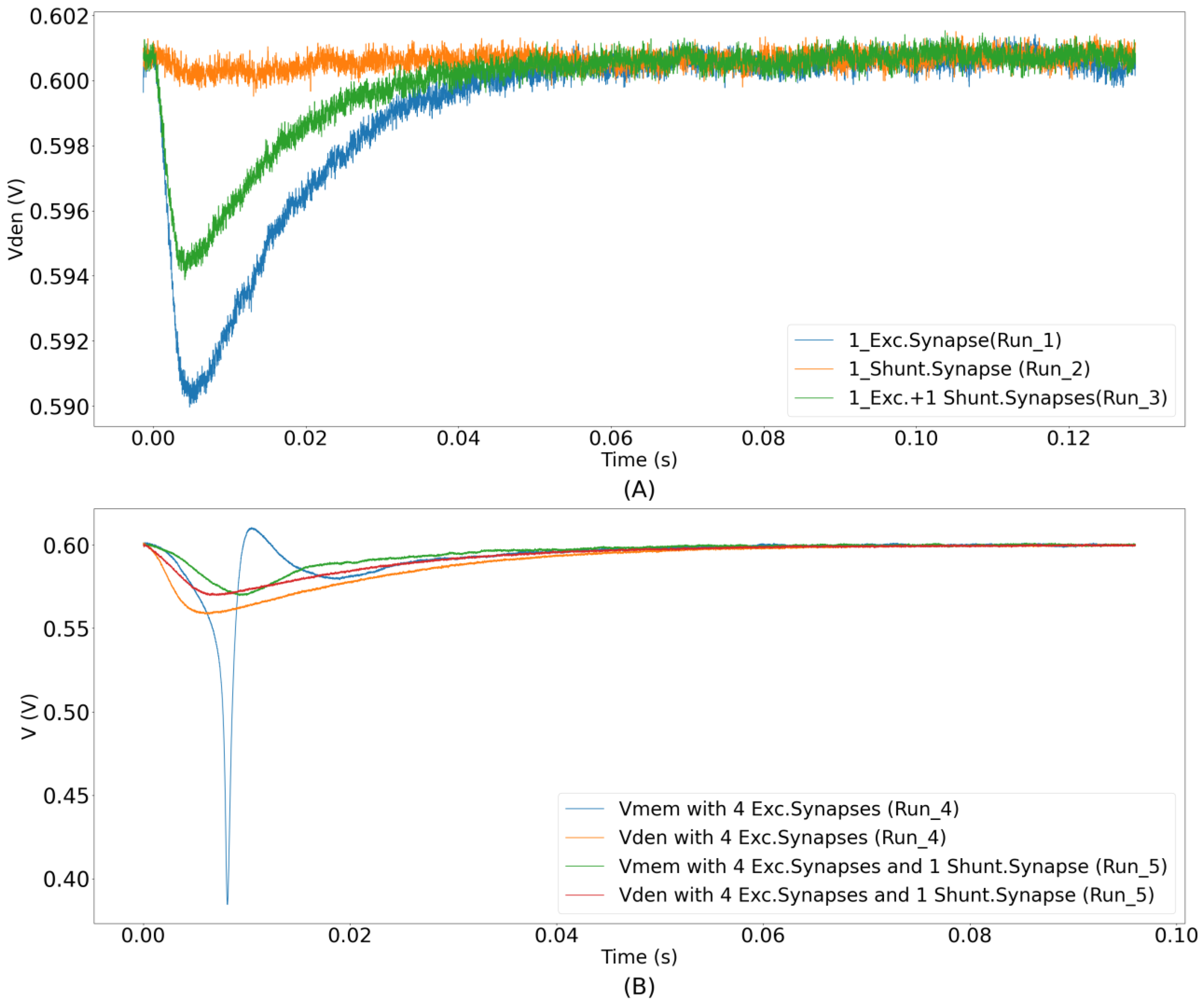
The unidirectional two-compartment neuron configuration was described in detail in a previous study [
29]. It has a somatic compartment comprising a soma circuit and dendritic compartment comprising a leak resistor (
) and dendritic capacitor (
) for integrating the synaptic current induced by the synapse circuits. The membrane potentials of the somatic and dendritic compartments are represented by V
mem and V
den, respectively. Based on their potential difference, current flows into or out of the somatic compartment via a unidirectional resistor (
). As the name implies, no current flows into or out of the dendritic compartment via the unidirectional resistor as would occur in an ideal two-compartment neuron model. The dendritic capacitor
is approximately 8.5 pF. The leak resistor
is implemented using one synapse circuit in the conductance-based configuration. This neuron configuration is used to demonstrate the shunting inhibition.
In the single-compartment neuron configuration, the membrane capacitance of the soma integrates the synaptic current. However, in the circuit implementation, if synapse circuits are connected directly to the soma circuit, their parasitic capacitance and leakage current disturb the spiking dynamics of the soma circuit. Hence, a bidirectional current conveyor circuit that replicates the current induced by the synapse circuits into the soma circuit is used as a link between them. Its two output branches induce currents with opposite polarities. I
out has the same polarity as I
in and I
out_rev has the opposite polarity. The current conveyor circuit is a current-controlled current source that fixes the node voltage common to the output terminals of all 256 synapse circuits (V
post in
Figure 5 and
Figure 6B) at a fixed value, approximately
= 600 mV. Thus, in this single-compartment configuration, the induced synaptic current depends only on the voltage parameters configuring the synapse circuit. As the node V
post is fixed to a constant value, the conductance-based synapse circuits act as excitatory synapse circuits (when connected to the soma circuit via the output terminal I
out_rev and with
fixed higher than
or V
post). In the experiments, the voltage bias
was 630 mV (370 mV below
). For relatively weaker bias voltages (
= 700 mV), the precise shapes and timings of the current induced by the synapse circuits were not conveyed to the soma circuit. However, with a stronger bias voltage
= 630 mV) the circuit consumes relatively higher power and induces significant noise. The noise is due to thermal noise in silicon and the bias voltage source. The ripple noise of the power line was extremely low as ultralow ripple power supplies were used. The induced thermal noise caused the soma circuit’s resting membrane potential (when around 800 mV) to vary randomly by approximately 50 mV. This random variation was higher for resting membrane potential values close to the spiking threshold of the soma circuit (700 mV in single-compartment configuration). To minimize this noise,
Vdd out and
were fixed at 949 mV and 50 mV, respectively, i.e., smaller and larger than their ideal values of 1 and 0 V, respectively. This reduced the random variation to approximately 30 mV (at a resting membrane potential of 800 mV). Furthermore, to minimize the effect of thermal noise, the resting membrane potential of the soma circuit was increased to approximately 850 mV. The power consumed by this circuit was approximately 90 nW (measured in the Spectre simulation). In this configuration, the 256 conductance-based synapse circuits were connected to the soma circuit via the current conveyor’s output terminal I
out_rev. The spike pattern detection was performed in this configuration to demonstrate that the switching nature of the conductance stage did not affect the performance of the task.
In terms of the complexity and biological plausibility, the unidirectional two-compartment model lies between the single- and two-compartment neuron models. The single-compartment neuron configuration cannot be used to demonstrate shunting inhibition, because the postsynaptic node (output terminals of the synapse circuits, V
post) is fixed at a constant value by the feedback action of the current conveyor circuit. In this study, we present three experimental results, as summarized in
Table 1.
2.3.2. Learning Circuit
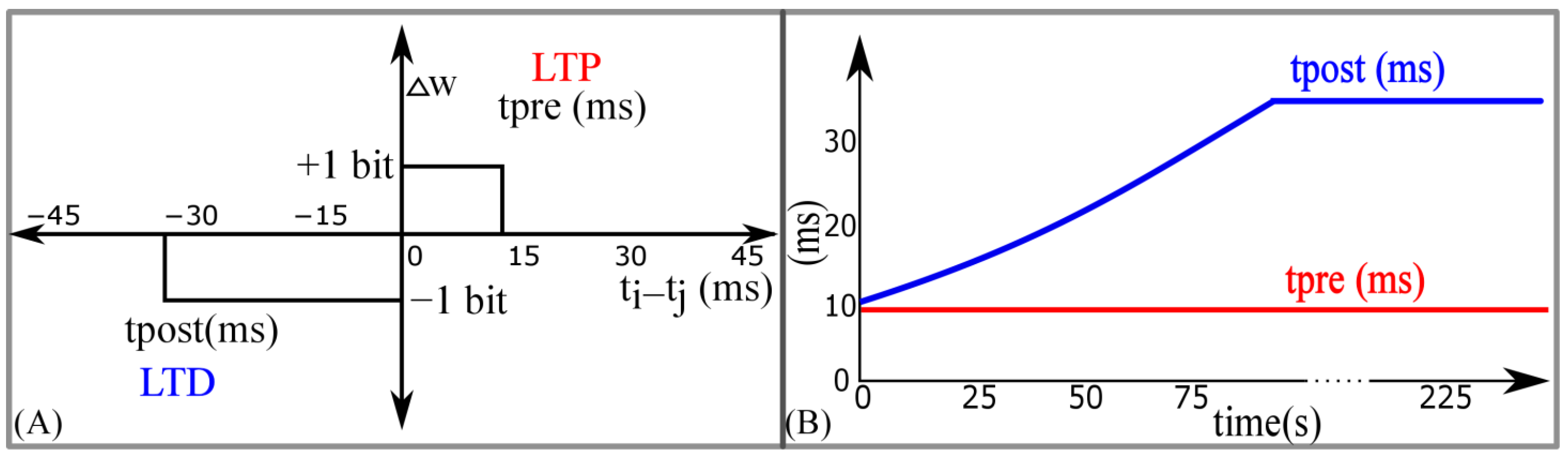
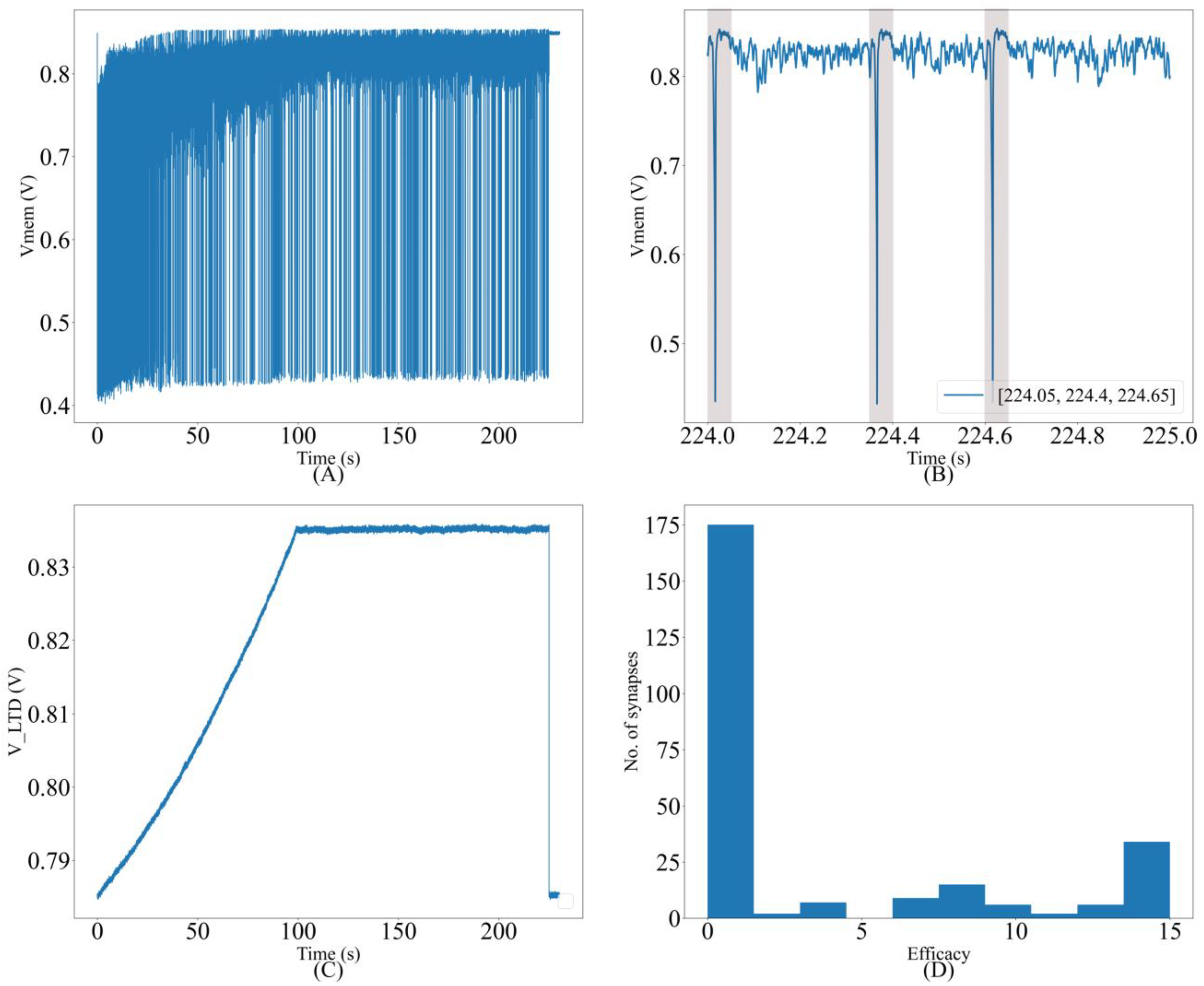
Similar to STDP learning, the adaptive STDP learning rule updates the synaptic efficacy based on the time difference between the pre- and postsynaptic spikes. However, the update in the efficacy is restricted to ±1 bits. This ±1 bit update is encoded by the rectangular STDP learning function (
Figure 7A) and is mathematically expressed as follows:
where
is the maximum delay of the postsynaptic spike after the presynaptic spike leading to potentiation (LTP).
is the maximum delay of the presynaptic spike after the postsynaptic spike leading to depression (LTD);
and
represent the timing of the pre- and postsynaptic spikes, respectively. The efficacy saturates at its maximum (
) and minimum (
) values. The learning parameter
is kept constant during learning and
is increased, as shown in
Figure 7B. The details of the adaptive STDP learning are provided in a previous study [
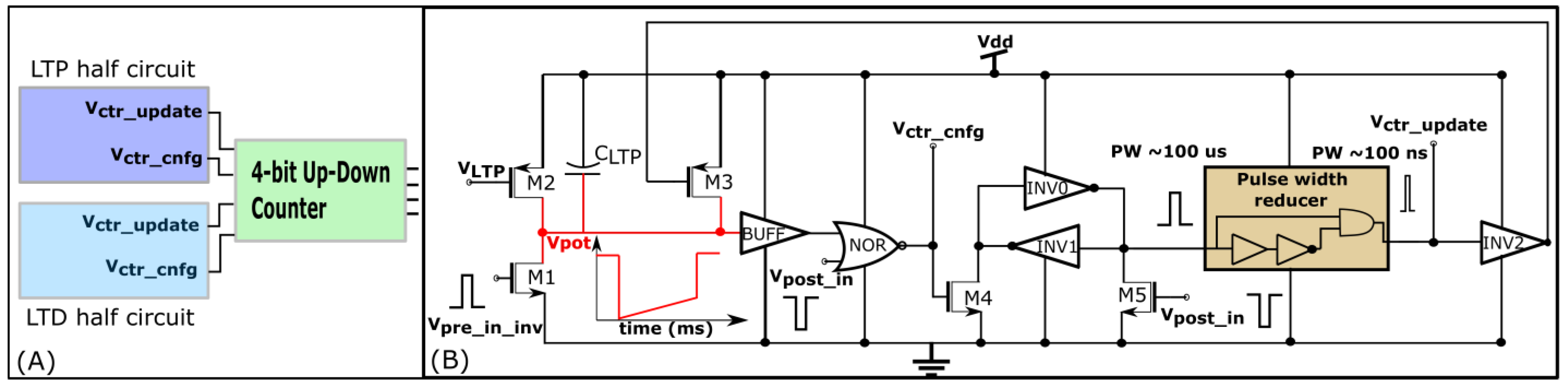
29]. Each synapse circuit has a learning circuit to implement the adaptive STDP learning. A block diagram is shown in
Figure 8A. The synaptic efficacy is stored in a four-bit up-down counter and updated by the circuits controlling its LTP and LTD. A conceptual schematic of the half-circuit controlling the LTP of the synaptic efficacy is shown in
Figure 8B. The details of the circuit operation are provided in another study [
30]. The value of
(which controls
) was fixed at 780 mV. The initial value of
(that controls
) was fixed at 783 mV and was adapted to higher values during learning (as shown in
Figure 7B). The chip does not contain an adaptation circuitry, and the adaptation of
was controlled via an external voltage source.
2.3.3. Spike Pattern Detection Task
The goal of the spike pattern detection task is to detect a 50 ms long spike pattern hidden within stochastic input spike trains at irregular intervals using a single neuron in an unsupervised manner. The neuron receives spike trains via synapses ( is the number of afferents). These spike trains are generated independently via an inhomogeneous Poisson process. The instantaneous firing rate ranges between 0 Hz and 90 Hz (the minimum time period for changing from 0 Hz to 90 Hz is 50 ms). Each afferent spikes at least once in a 50 ms duration, fixing 20 Hz as the minimum spiking frequency. Upon the generation of stochastic spike train (with a length of 225 s), a random 50 ms long segment (the target spike pattern) is chosen and copied. Subsequently, the original spike train is segmented into 50 ms long sections. Depending on the required spike pattern repetition frequency (chosen as 25 or 10%), certain randomly chosen sections are replaced by the target spike pattern. Consecutive 50 ms sections are avoided in this copy–paste process. This process ensures that only the specific spike time of the afferents distinguishes the spike pattern. The population average spike rate (measured in 10 ms time bins) is approximately the same inside and outside the spike patterns (approximately 54 Hz). These spike trains are used as inputs with 256 for the spike pattern detection experiment. The spike trains are 225 s long, and 50 runs were performed for each experimental setup.
The ideal STDP learning model has been shown to perform well in such spatiotemporal pattern detection tasks; however, its circuit implementation requires high-resolution synaptic efficacy [
5,
28]. In contrast, low-power circuits generally adopt memory devices under five bits for synaptic efficacy, owing to the silicon area and power constraints. Thus, we propose a hardware-friendly, bioinspired learning rule called adaptive STDP. In previous studies, the task described above was solved using adaptive STDP learning via simulations [
29] and circuit experiments [
30] with current-based four-bit excitatory synapses. In this study, we solved this task using the conductance-based four-bit synapse circuit described in
Section 2.2.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}