
Assessment of Optimal Conditions for Marine Invertebrate Cell-Mediated Mineralization of Organic Matrices


Abstract
1. Introduction
2. Materials and Methods
2.1. Collagen Preparation
2.2. Collagen Mineralization
2.3. pH Analysis
2.4. Scanning Electron Microscopy (SEM) and Energy Dispersive Spectroscopic (EDS) Analysis
2.5. X-ray Diffraction (XRD) Analysis
3. Results
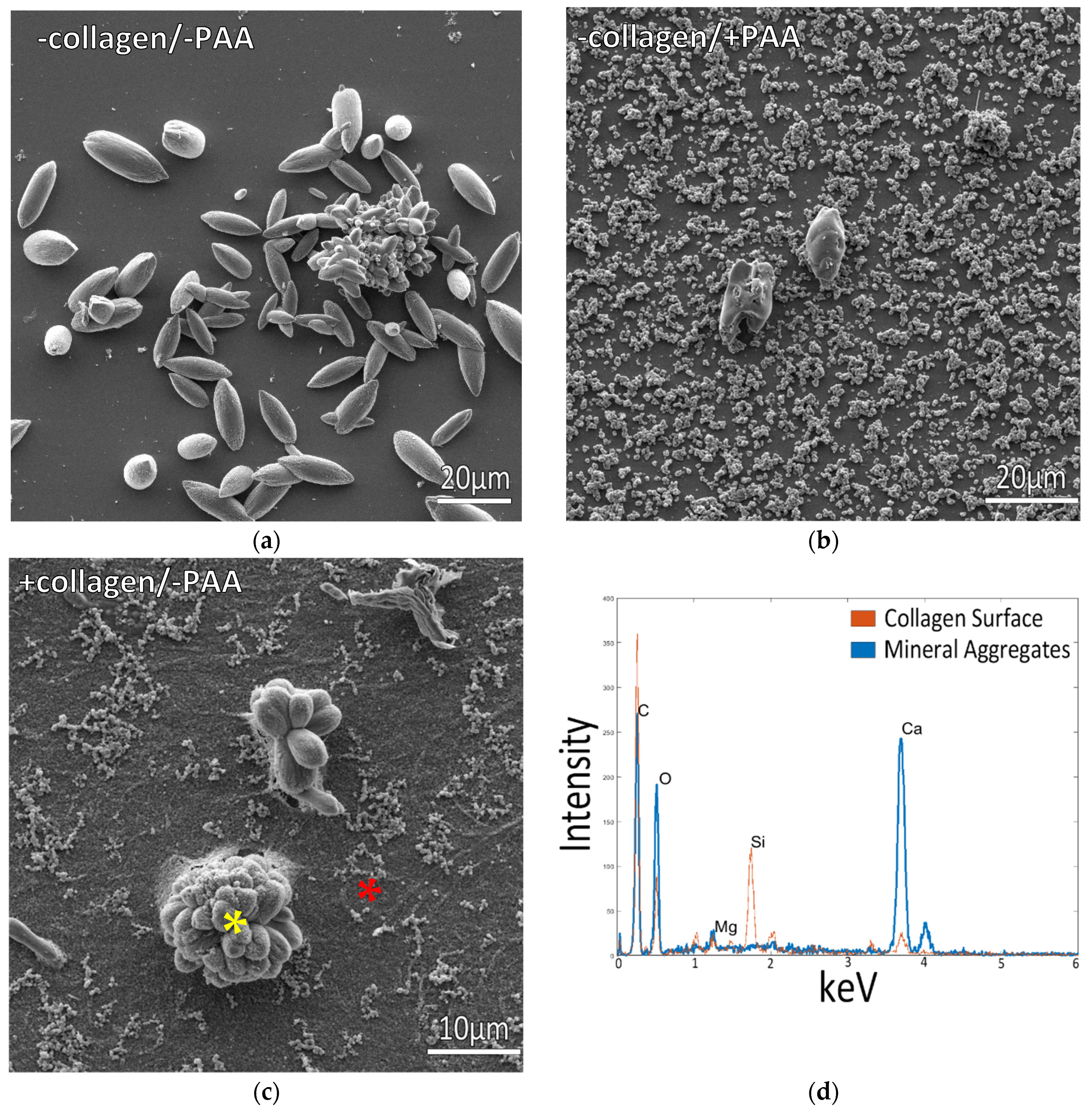
3.1. Mineralization at Room Temperature
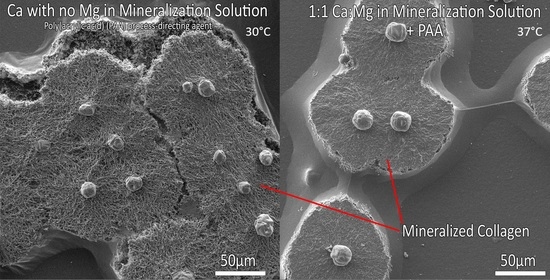
3.2. Mineralization at 37 °C
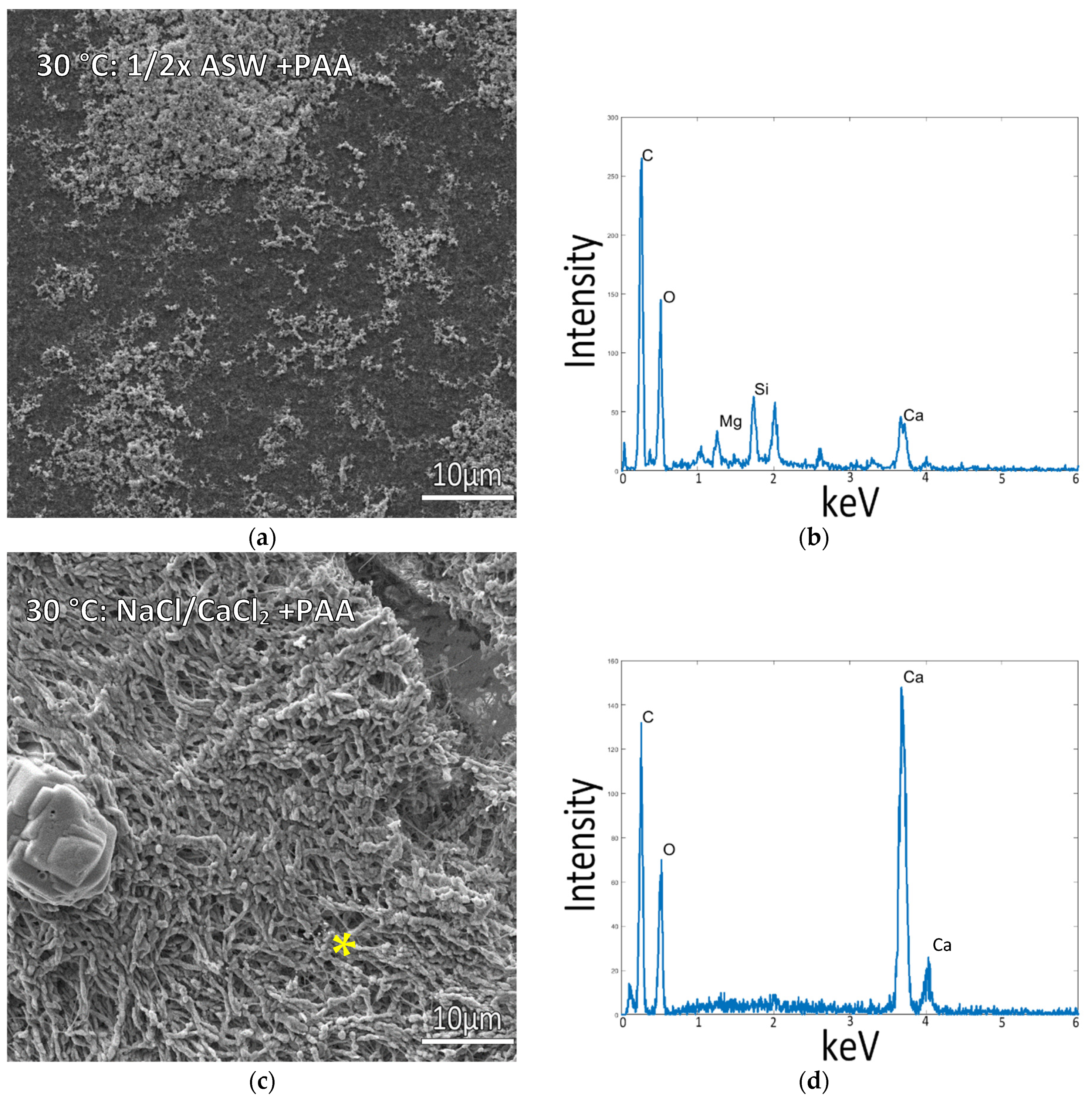
3.3. Mineralization at 30 °C
3.4. Influence of Calcium:Magnesium Ratio
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fratzl, P.; Weinkamer, R. Nature’s hierarchical materials. Prog. Mater. Sci. 2007, 52, 1263–1334. [Google Scholar] [CrossRef]
- Launey, M.E.; Ritchie, R.O. On the fracture toughness of advanced materials. Adv. Mater. 2009, 21, 2103–2110. [Google Scholar] [CrossRef]
- Wegst, U.G.K.; Bai, H.; Saiz, E.; Tomsia, A.P.; Ritchie, R.O. Bioinspired structural materials. Nat. Mater. 2015, 14, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, M.A.A.; Pack, S.P. Biomimetic and bioinspired silicifications: Recent advances for biomaterial design and applications. Acta Biomater. 2021, 120, 38–56. [Google Scholar] [CrossRef]
- Kul’chin, Y.N.; Bagaev, S.N.; Bukin, O.A.; Voznesenskiǐ, S.S.; Drozdov, A.L.; Zinin, Y.A.; Nagornyǐ, I.G.; Pestryakov, E.V.; Trunov, V.I. Photonic crystals based on natural oceanic biominerals. Tech. Phys. Lett. 2008, 34, 633–635. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Peres, B.U.; Carvalho, R.M.; MacLachlan, M.J. Photonic Hydrogels from Chiral Nematic Mesoporous Chitosan Nanofibril Assemblies. Adv. Funct. Mater. 2016, 26, 2875–2881. [Google Scholar] [CrossRef]
- Prozorov, T.; Bazylinski, D.A.; Mallapragada, S.K.; Prozorov, R. Novel magnetic nanomaterials inspired by magnetotactic bacteria: Topical review. Mater. Sci. Eng. R Rep. 2013, 74, 133–172. [Google Scholar] [CrossRef]
- Weiner, S.; Traub, W.; Lowenstam, H.A. Organic Matrix in Calcified Exoskeletons. Biominer. Biol. Met. Accumul. 1983, 205–224. [Google Scholar] [CrossRef]
- Landis, W.J.; Silver, F.H. Mineral Deposition in the Extracellular Matrices of Vertebrate Tissues: Identification of Possible Apatite Nucleation Sites on Type I Collagen. Cells Tissues Organs 2009, 189, 20–24. [Google Scholar] [CrossRef]
- Xu, Y.F.; Nudelman, F.; Eren, E.D.; Wirix, M.J.M.; Cantaert, B.; Nijhuis, W.H.; Hermida-Merino, D.; Portale, G.; Bomans, P.H.H.; Ottmann, C.; et al. Intermolecular channels direct crystal orientation in mineralized collagen. Nat. Commun. 2020, 11, 5068. [Google Scholar] [CrossRef]
- Bin San Chan, V.; Johnstone, M.B.; Wheeler, A.P.; Mount, A.S. Chitin Facilitated Mineralization in the Eastern Oyster. Front. Mar. Sci. 2018, 5, 347. [Google Scholar] [CrossRef]
- Ehrlich, H.; Luczak, M.; Ziganshin, R.; Mikšík, I.; Wysokowski, M.; Simon, P.; Baranowska-Bosiacka, I.; Kupnicka, P.; Ereskovsky, A.; Galli, R.; et al. Arrested in Glass: Actin within Sophisticated Architectures of Biosilica in Sponges. Adv. Sci. 2022, 9, 2105059. [Google Scholar] [CrossRef]
- Ehrlich, H.; Deutzmann, R.; Brunner, E.; Cappellini, E.; Koon, H.; Solazzo, C.; Yang, Y.; Ashford, D.; Thomas-Oates, J.; Lubeck, M.; et al. Mineralization of the metre-long biosilica structures of glass sponges is templated on hydroxylated collagen. Nat. Chem. 2010, 2, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Meyers, M.A.; Lim, C.T.; Li, A.; Hairul Nizam, B.R.; Tan, E.P.S.; Seki, Y.; McKittrick, J. The role of organic intertile layer in abalone nacre. Mater. Sci. Eng. C 2009, 29, 2398–2410. [Google Scholar] [CrossRef]
- Checa, A.G.; Cartwright, J.H.E.; Willinger, M.G. Mineral bridges in nacre. J. Struct. Biol. 2011, 176, 330–339. [Google Scholar] [CrossRef]
- Gebauer, D.; Völkel, A.; Cölfen, H. Stable prenucleation calcium carbonate clusters. Science 2008, 322, 1819–1822. [Google Scholar] [CrossRef]
- Gebauer, D.; Kellermeier, M.; Gale, J.D.; Bergström, L.; Cölfen, H. Pre-nucleation clusters as solute precursors in crystallisation. Chem. Soc. Rev. 2014, 43, 2348–2371. [Google Scholar] [CrossRef]
- Bewernitz, M.A.; Gebauer, D.; Long, J.; Cölfen, H.; Gower, L.B. A metastable liquid precursor phase of calcium carbonate and its interactions with polyaspartate. Faraday Discuss. 2012, 159, 291. [Google Scholar] [CrossRef]
- Wolf, S.E.; Gower, L.B. Challenges and Perspectives of the Polymer-Induced Liquid-Precursor Process: The Pathway from Liquid-Condensed Mineral Precursors to Mesocrystalline Products. New Perspect. Miner. Nucl. Growth 2017, 3, 43–75. [Google Scholar] [CrossRef]
- Gower, L.; Elias, J. Colloid assembly and transformation (CAT): The relationship of PILP to biomineralization. J. Struct. Biol. X 2022, 6, 100059. [Google Scholar] [CrossRef]
- Gower, L.B. Biomimetic Model Systems for Investigating the Amorphous Precursor Pathway and Its Role in Biomineralization. Chem. Rev. 2008, 108, 4551–4627. [Google Scholar] [CrossRef] [PubMed]
- Boskey, A.L.; Villarreal-Ramirez, E. Intrinsically disordered proteins and biomineralization. Matrix Biol. 2016, 52–54, 43–59. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.P.; Russ, J.A.; Verch, A.; Kröger, R.; Estroff, L.A.; Evans, J.S. The Intrinsically Disordered C-RING Biomineralization Protein, AP7, Creates Protein Phases That Introduce Nanopatterning and Nanoporosities into Mineral Crystals. Biochemistry 2014, 53, 4317–4319. [Google Scholar] [CrossRef]
- Rao, A.; Drechsler, M.; Schiller, S.; Scheffner, M.; Gebauer, D.; Cölfen, H. Stabilization of Mineral Precursors by Intrinsically Disordered Proteins. Adv. Funct. Mater. 2018, 28, 1802063. [Google Scholar] [CrossRef]
- Pendola, M.; Jain, G.; Evans, J.S. Skeletal development in the sea urchin relies upon protein families that contain intrinsic disorder, aggregation-prone, and conserved globular interactive domains. PLoS ONE 2019, 14, e0222068. [Google Scholar] [CrossRef]
- Wojtas, M.; Dobryszycki, P.; Ożyhar, A. Intrinsically Disordered Proteins in Biomineralization. Adv. Top. Biominer. 2012, 1, 3–32. [Google Scholar] [CrossRef]
- Fisher, L.W.; Torchia, D.A.; Fohr, B.; Young, M.F.; Fedarko, N.S. Flexible structures of SIBLING proteins, bone sialoprotein, and osteopontin. Biochem. Biophys. Res. Commun. 2001, 280, 460–465. [Google Scholar] [CrossRef]
- Gower, L.B.; Odom, D.J. Deposition of calcium carbonate films by a polymer-induced liquid-precursor (PILP) process. J. Cryst. Growth 2000, 210, 719–734. [Google Scholar] [CrossRef]
- Olszta, M.J.; Cheng, X.; Jee, S.S.; Kumar, R.; Kim, Y.-Y.; Kaufman, M.J.; Douglas, E.P.; Gower, L.B. Bone structure and formation: A new perspective. Mater. Sci. Eng. R Rep. 2007, 58, 77–116. [Google Scholar] [CrossRef]
- Nagasawa, H.; Luquet, G. The crustacean cuticle: Structure, composition and mineralization. Front. Biosci.-Elite 2012, 4, 711–720. [Google Scholar] [CrossRef]
- Aizenberg, J.; Addadi, L.; Weiner, S.; Lambert, G. Stabilization of amorphous calcium carbonate by specialized macromolecules in biological and synthetic precipitates. Adv. Mater. 1996, 8, 222–226. [Google Scholar] [CrossRef]
- Luquet, G. Biomineralizations: Insights and prospects from crustaceans. Zookeys 2012, 176, 103–121. [Google Scholar] [CrossRef] [PubMed]
- Belcher, A.M.; Wu, X.H.; Christensen, R.J.; Hansma, P.K.; Stucky, G.D.; Morse, D.E. Control of crystal phase switching and orientation by soluble mollusc-shell proteins. Nature 1996, 381, 56. [Google Scholar] [CrossRef]
- Kim, Y.-Y.; Douglas, E.P.; Gower, L.B. Patterning Inorganic (CaCO3) Thin Films via a Polymer-Induced Liquid-Precursor Process. Langmuir 2007, 23, 4862–4870. [Google Scholar] [CrossRef]
- Cheng, X.; Gower, L.B. Molding mineral within microporous hydrogels by a polymer-induced liquid-precursor (PILP) process. Biotechnol. Prog. 2006, 22, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Gower, L.A.; Tirrell, D.A. Calcium carbonate films and helices grown in solutions of poly(aspartate). J. Cryst. Growth 1998, 191, 153–160. [Google Scholar] [CrossRef]
- Amos, F.F.; Sharbaugh, D.M.; Talham, D.R.; Gower, L.B.; Fricke, M.; Volkmer, D. Formation of Single-Crystalline Aragonite Tablets/Films via an Amorphous Precursor. Langmuir 2007, 23, 1988–1994. [Google Scholar] [CrossRef]
- Olszta, M.J.; Douglas, E.P.; Gower, L.B. Scanning Electron Microscopic Analysis of the Mineralization of Type I Collagen via a Polymer-Induced Liquid-Precursor (PILP) Process. Calcif. Tissue Int. 2003, 72, 583–591. [Google Scholar] [CrossRef]
- Ping, H.; Xie, H.; Wan, Y.; Zhang, Z.; Zhang, J.; Xiang, M.; Xie, J.; Wang, H.; Wang, W.; Fu, Z. Confinement controlled mineralization of calcium carbonate within collagen fibrils. J. Mater. Chem. B 2016, 4, 880–886. [Google Scholar] [CrossRef]
- Niu, L.-N.; Jiao, K.; Qi, Y.-P.; Yiu, C.K.Y.; Ryou, H.; Arola, D.D.; Chen, J.-H.; Breschi, L.; Pashley, D.H.; Tay, F.R.; et al. Infiltration of Silica Inside Fibrillar Collagen. Angew. Chemie Int. Ed. 2011, 50, 11688–11691. [Google Scholar] [CrossRef]
- Nudelman, F.; Pieterse, K.; George, A.; Bomans, P.H.H.; Friedrich, H.; Brylka, L.J.; Hilbers, P.A.J.; De With, G.; Sommerdijk, N.A.J.M. The role of collagen in bone apatite formation in the presence of hydroxyapatite nucleation inhibitors. Nat. Mater. 2010, 9, 1004–1009. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H. Chitin and collagen as universal and alternative templates in biomineralization. Int. Geol. Rev. 2010, 52, 661–699. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, L.; Qi, H.; Zhao, Q.; Liu, Y.; Zhang, Y.; Zhang, J.; Qi, H.; Zhao, Q.; Zhang, Y.; et al. Dual Function of Magnesium in Bone Biomineralization. Adv. Healthc. Mater. 2019, 8, 1901030. [Google Scholar] [CrossRef] [PubMed]
- Ng, A.H.M.; Hercz, G.; Kandel, R.; Grynpas, M.D. Association between fluoride, magnesium, aluminum and bone quality in renal osteodystrophy. Bone 2004, 34, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Hand, C. Another sea anemone from California and the types of certain Californian anemones. J. Washingt. Acad. Sci. 1957, 47, 411–414. [Google Scholar]
- Schlüter, L.; Lohbeck, K.T.; Gutowska, M.A.; Gröger, J.P.; Riebesell, U.; Reusch, T.B.H. Adaptation of a globally important coccolithophore to ocean warming and acidification. Nat. Clim. Chang. 2014, 4, 1024–1030. [Google Scholar] [CrossRef]
- Nishino, Y.; Oaki, Y.; Imai, H. Magnesium-mediated nanocrystalline mosaics of calcite. Cryst. Growth Des. 2009, 9, 223–226. [Google Scholar] [CrossRef]
- Macías-Sánchez, E.; Tarakina, N.V.; Ivanov, D.; Blouin, S.; Berzlanovich, A.M.; Fratzl, P. Spherulitic Crystal Growth Drives Mineral Deposition Patterns in Collagen-Based Materials. Adv. Funct. Mater. 2022, 2200504. [Google Scholar] [CrossRef]
- Olszta, M.J.; Gajjeraman, S.; Kaufman, M.; Gower, L.B. Nanofibrous Calcite Synthesized via a Solution-Precursor-Solid Mechanism. Chem. Mater. 2004, 16, 2355–2362. [Google Scholar] [CrossRef]
- Homeijer, S.J.; Olszta, M.J.; Barrett, R.A.; Gower, L.B. Growth of nanofibrous barium carbonate on calcium carbonate seeds. J. Cryst. Growth 2008, 310, 2938–2945. [Google Scholar] [CrossRef]
- Politi, Y.; Batchelor, D.R.; Zaslansky, P.; Chmelka, B.F.; Weaver, J.C.; Sagi, I.; Weiner, S.; Addadi, L. Role of Magnesium Ion in the Stabilization of Biogenic Amorphous Calcium Carbonate: A Structure−Function Investigation. Chem. Mater. 2010, 22, 161–166. [Google Scholar] [CrossRef]
- Cheng, X.; Varona, P.L.; Olszta, M.J.; Gower, L.B. Biomimetic synthesis of calcite films by a polymer-induced liquid-precursor (PILP) process: 1. Influence and incorporation of magnesium. J. Cryst. Growth 2007, 307, 395–404. [Google Scholar] [CrossRef]
- Loste, E.; Wilson, R.M.; Seshadri, R.; Meldrum, F.C. The role of magnesium in stabilising amorphous calcium carbonate and controlling calcite morphologies. J. Cryst. Growth 2003, 254, 206–218. [Google Scholar] [CrossRef]
- Niu, L.; Jee, S.E.; Jiao, K.; Tonggu, L.; Li, M.; Wang, L.; Yang, Y.; Bian, J.; Breschi, L.; Jang, S.S.; et al. Collagen intrafibrillar mineralization as a result of the balance between osmotic equilibrium and electroneutrality. Nat. Mater. 2017, 16, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Bewernitz, M.A.; Lovett, A.C.; Gower, L.B. Liquid–Solid Core-Shell Microcapsules of Calcium Carbonate Coated Emulsions and Liposomes. Appl. Sci. 2020, 10, 8551. [Google Scholar] [CrossRef]
- Antao, S.M. Temperature dependence of the structural parameters in the transformation of aragonite to calcite, as determined from in situ synchrotron powder x-ray-diffraction data. Can. Mineral. 2010, 48, 1225–1236. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Concentration (mM) | |||
|---|---|---|---|---|
| Instant Ocean Seawater Mix | 1/2x ASW | NaCl + 10 mM Ca | NaCl + 10 mM Ca + 50 mM Mg | |
| Chloride | 544 | 272 | 160 | 160 |
| Sodium | 469 | 235 | 190 | 190 |
| Sulfate | 28 | 14 | 20 | 20 |
| Magnesium | 54 | 27 | 0 | 50 |
| Potassium | 10 | 5 | 10 | 10 |
| Calcium | 10 | 5 | 10 | 10 |
| Carbonate/bicarbonate | 3 | 2 | 0 * | 0 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elias, J.; Angelini, T.; Martindale, M.Q.; Gower, L. Assessment of Optimal Conditions for Marine Invertebrate Cell-Mediated Mineralization of Organic Matrices. Biomimetics 2022, 7, 86. https://doi.org/10.3390/biomimetics7030086
Elias J, Angelini T, Martindale MQ, Gower L. Assessment of Optimal Conditions for Marine Invertebrate Cell-Mediated Mineralization of Organic Matrices. Biomimetics. 2022; 7(3):86. https://doi.org/10.3390/biomimetics7030086
Chicago/Turabian StyleElias, Jeremy, Thomas Angelini, Mark Q. Martindale, and Laurie Gower. 2022. "Assessment of Optimal Conditions for Marine Invertebrate Cell-Mediated Mineralization of Organic Matrices" Biomimetics 7, no. 3: 86. https://doi.org/10.3390/biomimetics7030086
APA StyleElias, J., Angelini, T., Martindale, M. Q., & Gower, L. (2022). Assessment of Optimal Conditions for Marine Invertebrate Cell-Mediated Mineralization of Organic Matrices. Biomimetics, 7(3), 86. https://doi.org/10.3390/biomimetics7030086