To create biomimetic cilia movement, we produced a hinged servo robotic arm, consisting of three servos. Three Tower Pro SG50 Micro Digital Servos (Servo Shop, Frodsham, UK) were used to copy the whipping power and recovery stroke, with two servos mounted axially to create a two-segment robotic arm, and the third servo placed orthogonally at the base of the arm, to facilitate rotation of the cilia to stroke in different directions; this setup is shown in
Figure 4. The servos are connected to each other with the manufacturer-provided servo arms and screws. Each servo is controlled by an Atmega 328 8-bit AVR RISC-based microcontroller (RS Components, Corby, UK), with pulse-width-modulation angular control for each servo. The ciliary beating action was created by a sequence of angular controls to each servo in the armature in order to create the typical asymmetrical ciliary beating action. Each ciliary unit in the form of servo armature and relative microcontroller is mounted on a printed circuit board. The overall force of the three-servo cilia was recorded 12 times using a Sauter FK50 force gauge (RS Components, Corby, UK) attached to the tip of the cilia during the power stroke phase. All the files required to reproduce and reprogram the cilia can be obtained and are provided as open-source material for anyone to use provided citations and credit are given where appropriate (
https://github.com/UnconventionalComputingUWE/Paramecium).
2.2.1. Assembly
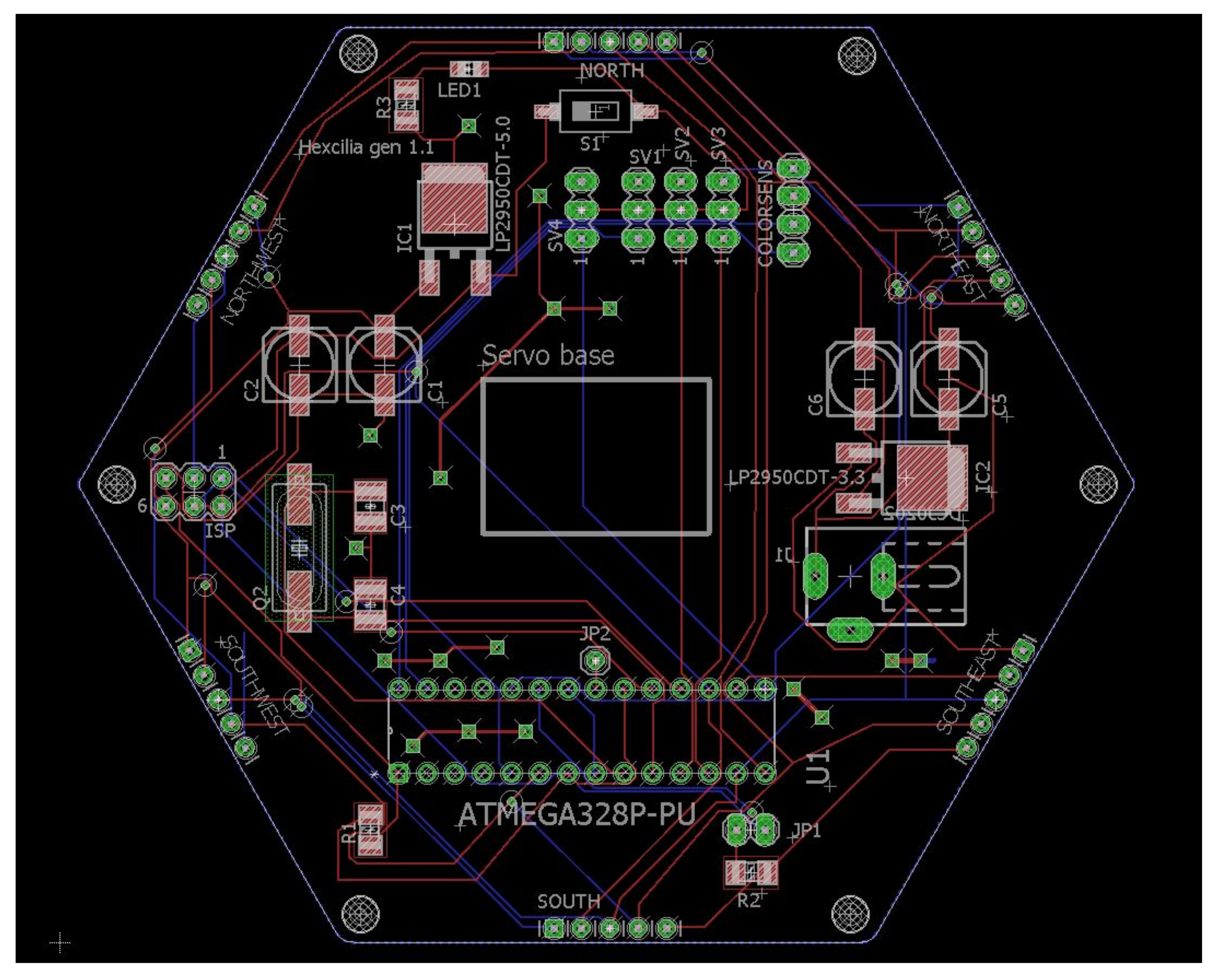
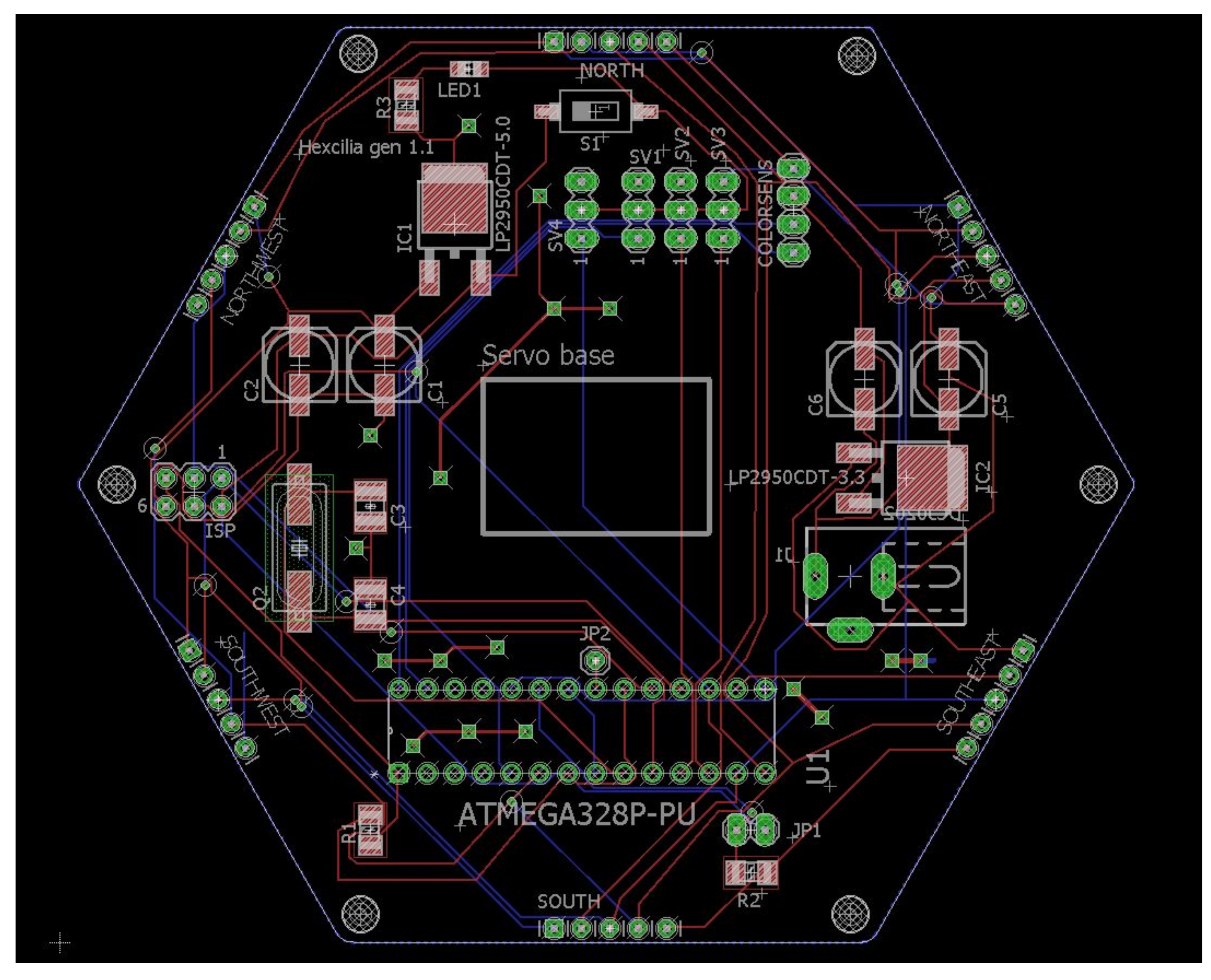
The circuit board is a mixture of surface mount devices (SMD) and through-hole components (
Figure 5); while surface mount components are often assembled by a pick-and-place machine, the components used in this project are easily assembled manually by hand by someone with reasonable soldering experience, as larger components have been purposefully chosen. Servos were mounted on to the board using super glue gel, in order to hold them in place during manipulation of heavier objects. Servo armatures were attached to the servo above using cable ties or super glue gel, and both provided acceptable strength of attachment. The ciliary tip was an extra long servo armature assembled by connecting two linkages together using screws provided with the servos. The color sensor board was again attached to the top using super glue gel. Each servo and the color sensor board were connected to the board using pin headers to facilitate simple swapping of servos.
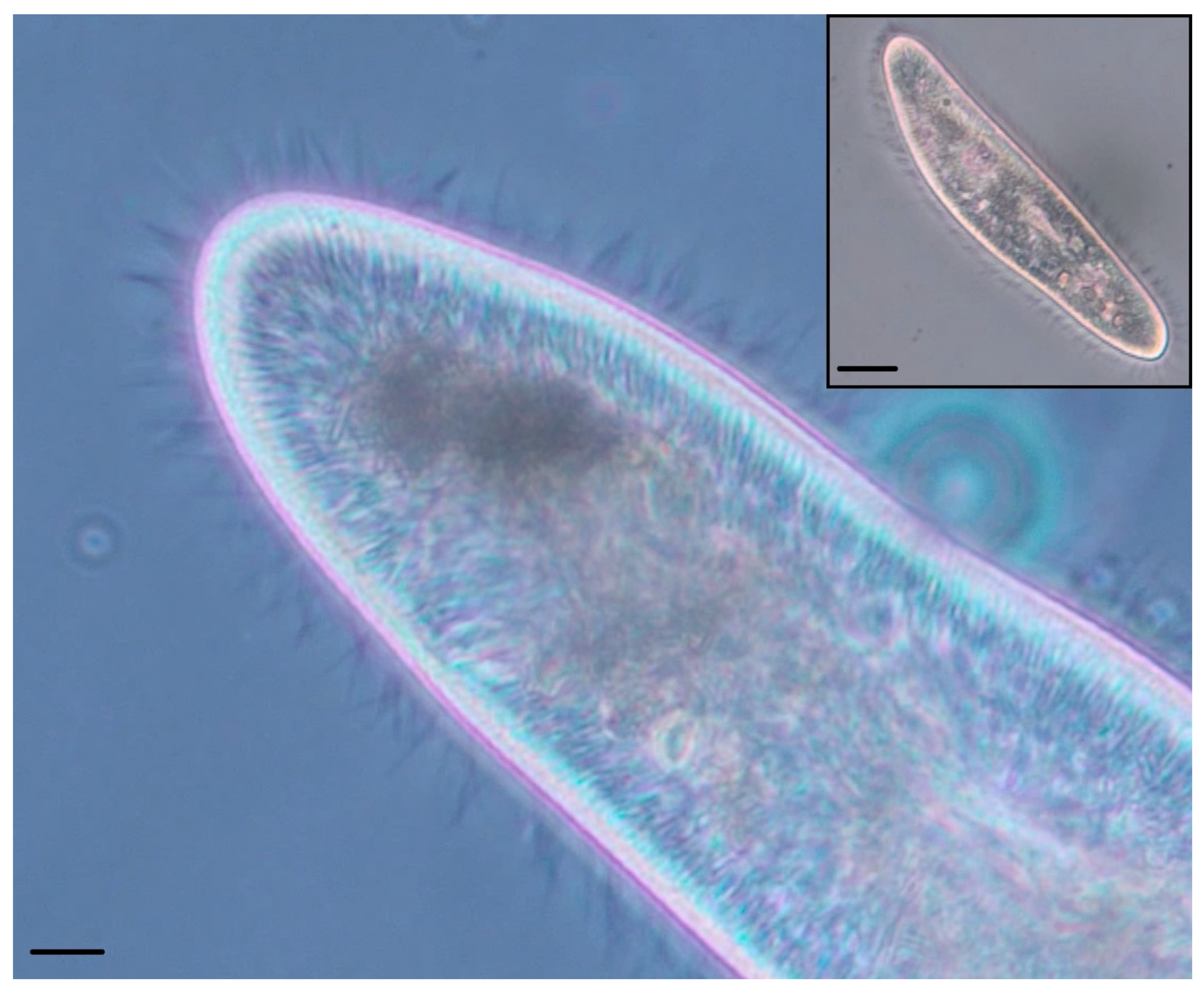
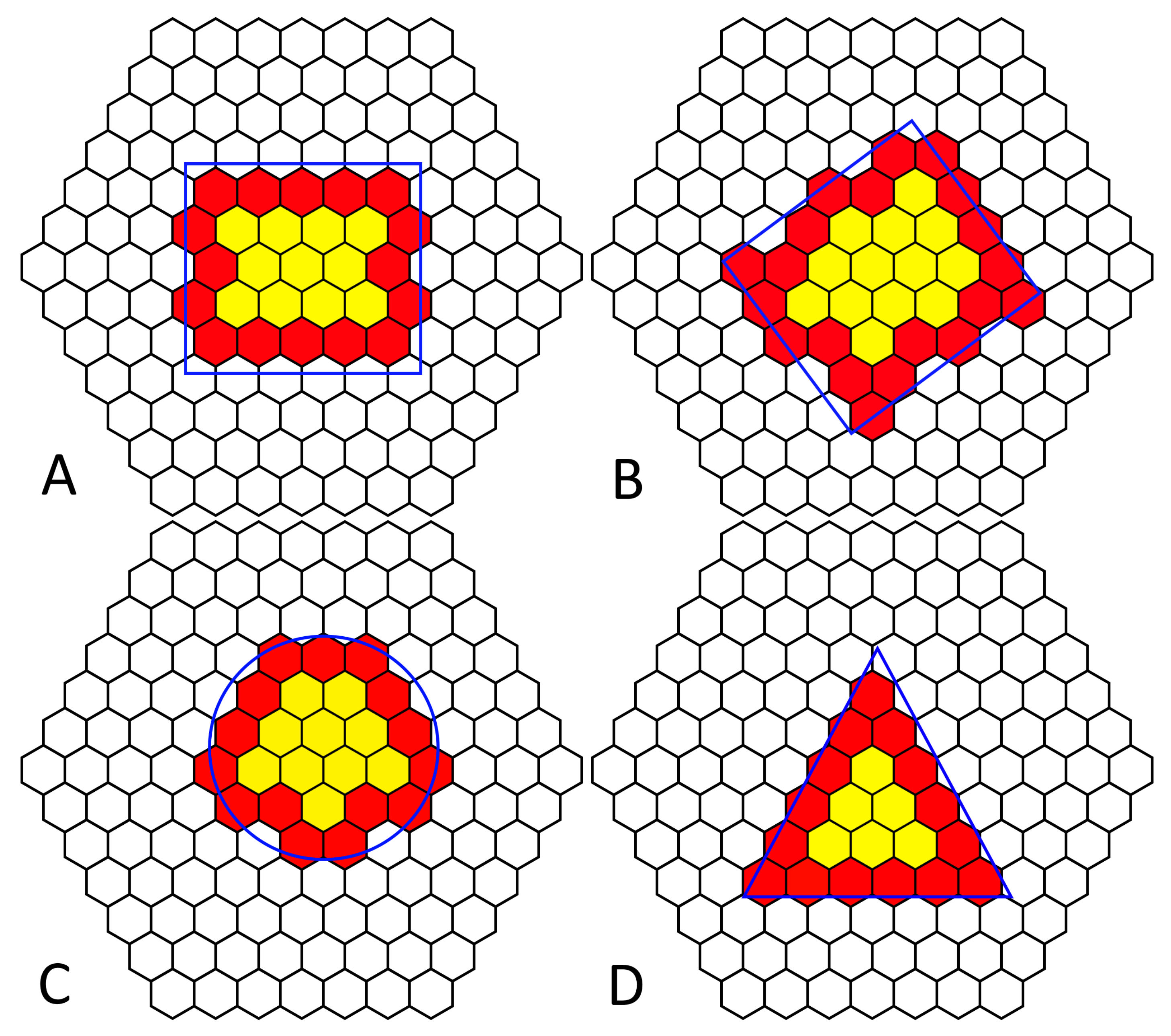
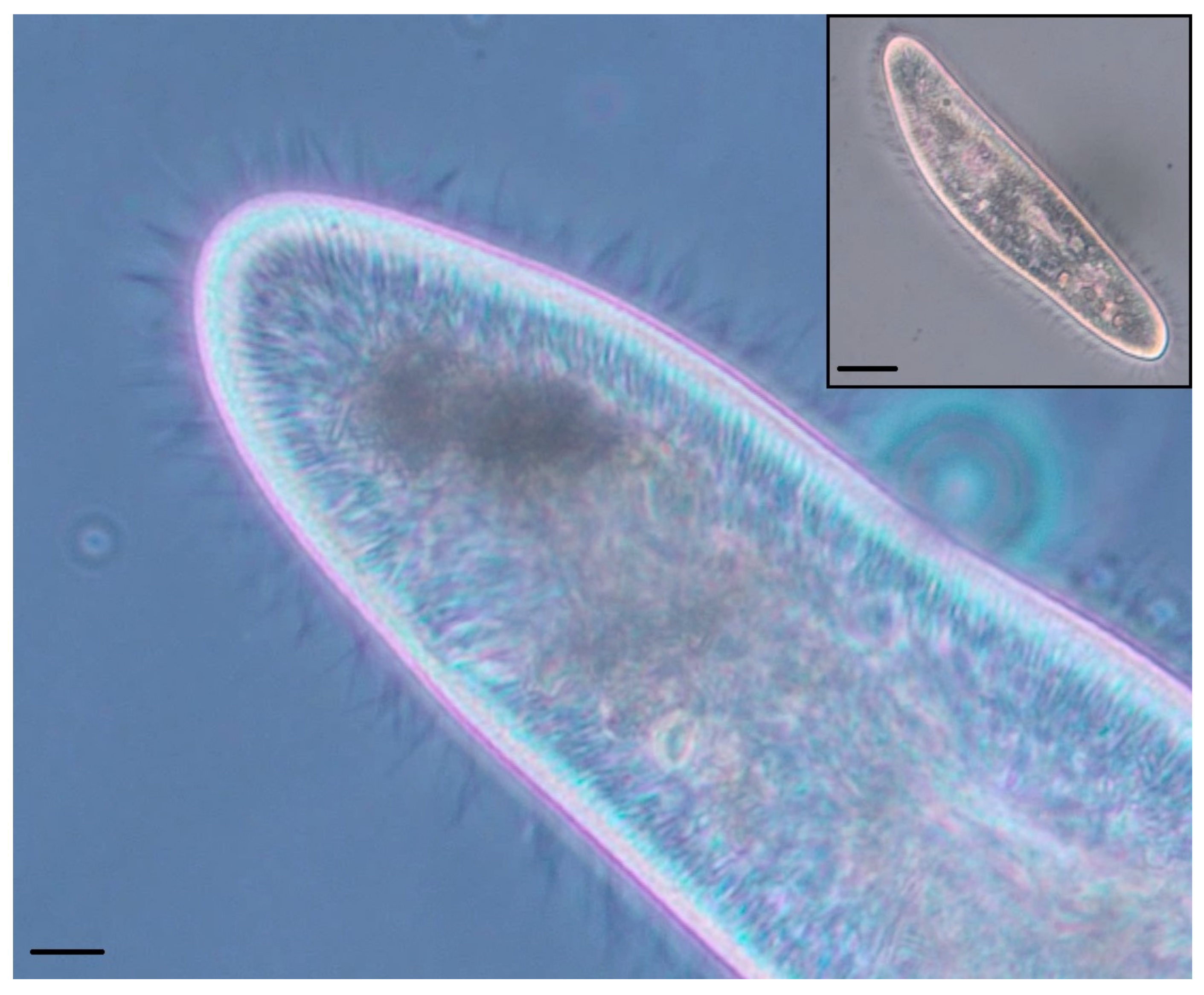
The organism’s membrane holds the cilia in alignment in the continuous outer membrane; under sufficient magnification, a hexagonal tessellation of longitudinal rows is visible (
Figure 2), with one cilia protruding from each hexagonal structure. It is because of this hexagonal tessellation that each ciliary unit’s circuit board has hexagonal outer dimensions. This tessellation provides opportunities for sharing power, providing a system that is entirely reconfigurable in morphology; additionally, it is significantly biomimetic in geometry [
14]. Each Printed Circuit Board (PCB) measured 95 × 83 mm, and a total of 35 boards was used to cover an area of approximately 50 × 39 cm. Due to the power sharing design of the boards, the number of boards that can be supported is limited to the sole power supply, rather than the number of individual power supplies to each board. In order to share data with its six neighboring cells, the boards have five pin headers along each edge of the board, and these align with pins on adjacent boards. Five pin jumper modules connect to digital Input/Output (I/O) pins on the microprocessor, which facilitates neighbor communication. Recording of the objects’ position and speed was performed in real time using a webcam mounted on a clamp stand above the assembled platform, with video being fed to a custom MATLAB (Mathworks, Natick, MA, USA) script for tracking object position. In order to facilitate smooth movement, the cilia shared data to coordinate beating. The Matlab script can be supplied by the Corresponding Author.
2.2.2. Color Detection
Paramecium grazes on food particles of different sizes and actively hunts for bacteria and minute protozoa, funneling them into the oral grove where they are sorted into edible and inedible and are passed into food vacuoles via the buccal cavity or rejected, respectively. As the focus of this work is to create a bioinspired sorting platform, a variety of sensors could be used, dependant on the practical application, of which there is a huge variety; the microcontroller can interact with a huge array of analogue or digital sensors, so desired sensing systems can be easily integrated for each application. In this instance, however, color sensors were employed to differentiate between objects. For simplicity, an Adafruit TCS34725 RGB color sensor (Adafruit, New York, NY, USA) was employed. The sensor has an inbuilt light-emitting diode (LED) and infrared (IR) filter to accurately detect a 3,800,000:1 dynamic range of color. The sensor board and supporting electronics were mounted atop the distal servo for optimum color detection, but did not extend above the servo’s working armature.
2.2.3. Software
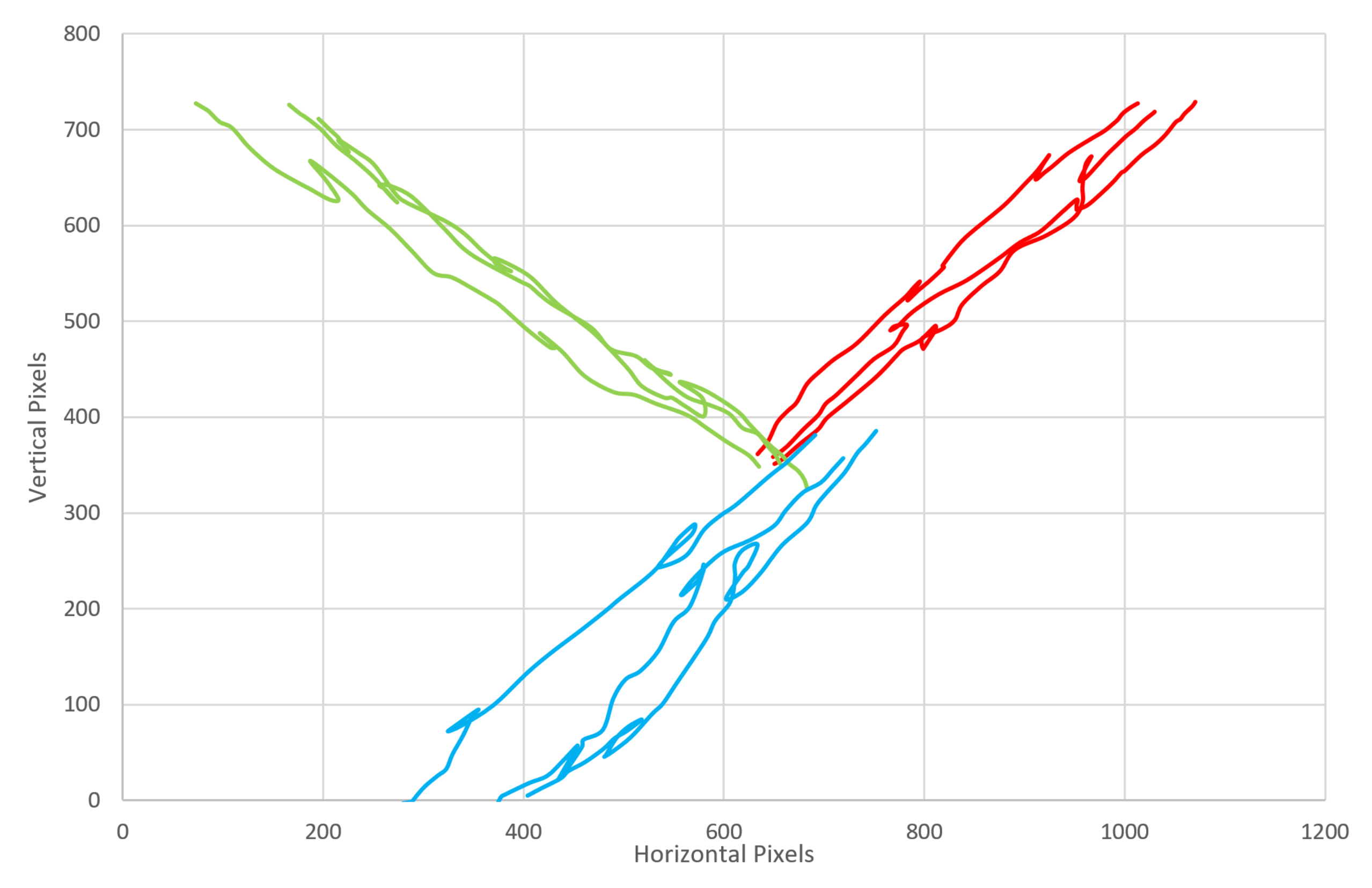
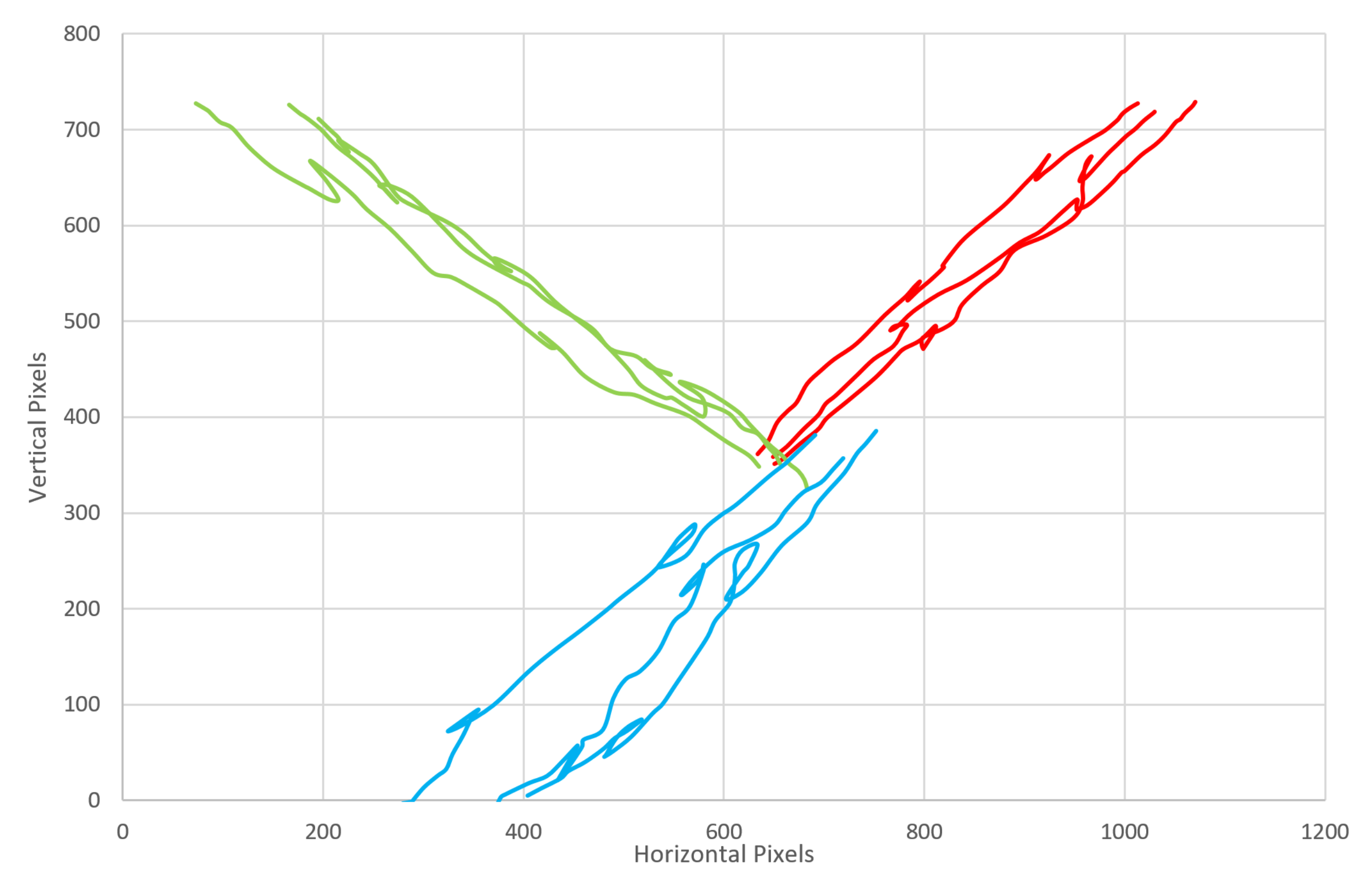
The embedded code on the microprocessor controls the system’s behavior; like the emergent intelligence of the Paramecia, a simple set of rules governs the organism’s response, leading to an apparently complex behavior. The software reads the value of the RGB sensor, and if a colored object is detected, the cilia will act; the base servo will rotate to an angle based on the relative RGB values and then perform a power and recovery stroke in that direction; this action is repeated until the RGB sensor no longer detects a colored object. Each cilia acts alone, but together, they are capable of transporting objects across the entire platform as each element knows its own behavior. It is possible, again if the application would desire it, to program each cilia or sections of cilia to act in different ways so as to sort objects in more complex manners, or with different sensors, to sort color and then weight, for example. In this instance, if the RGB sensor detects a red object, the cilia beats in a north-easterly direction (relative to the board); if a green object is identified, the cilia beats in a north-westerly direction; and if a blue object is detected, then the cilia beats in a south-westerly direction. It is possible to beat in any direction given a range of color values; however, for this proof-of-concept design, only three colors were used.
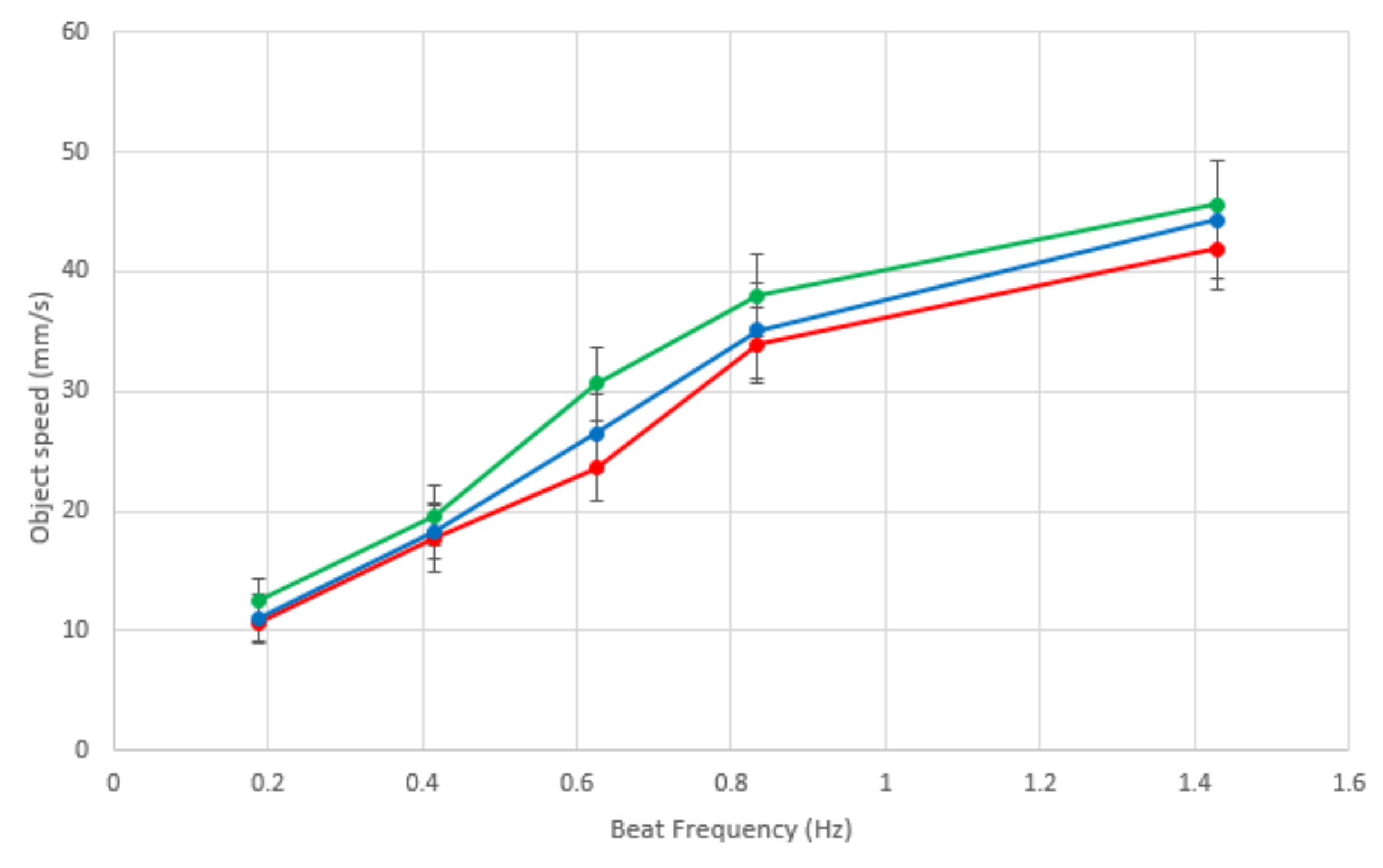
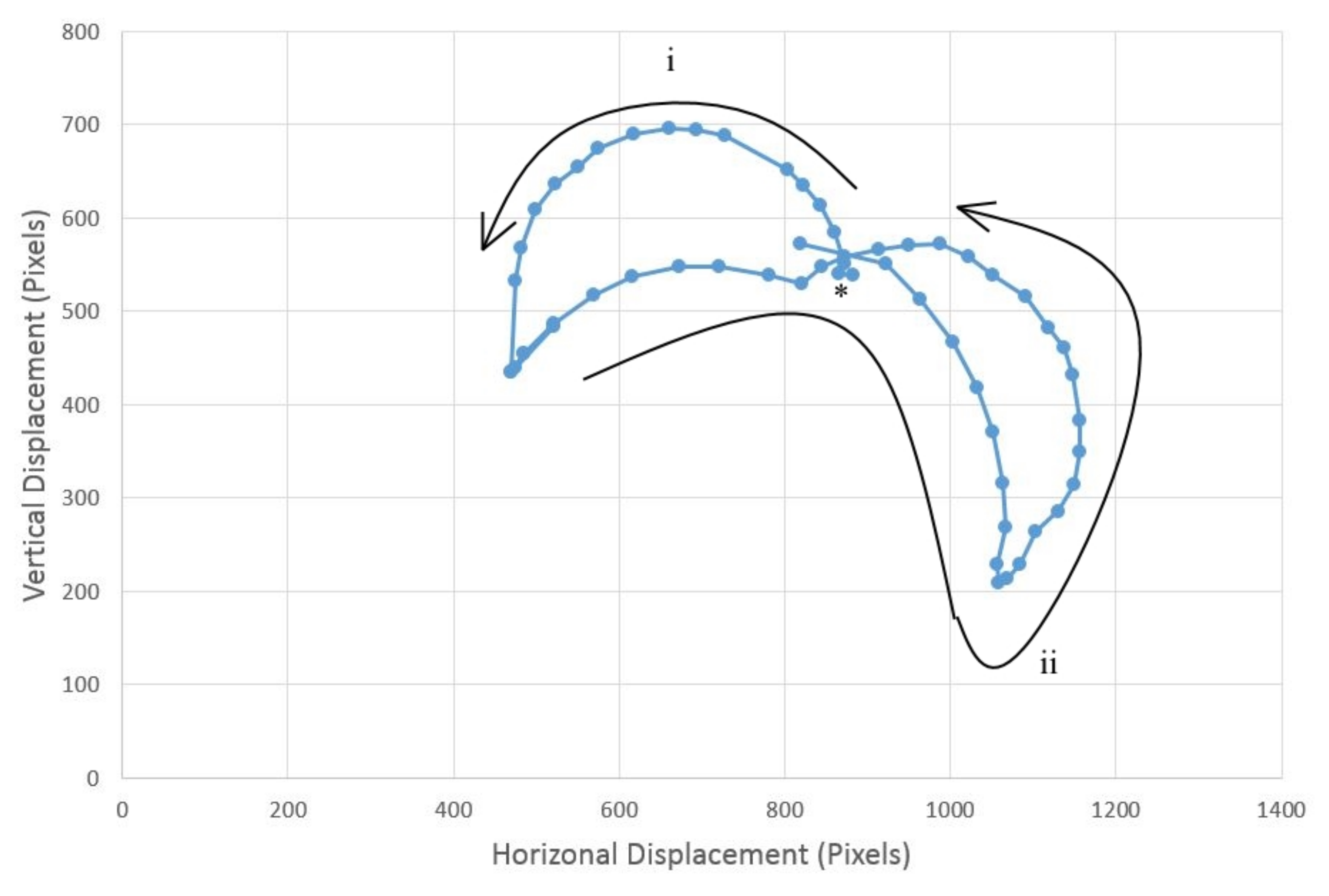
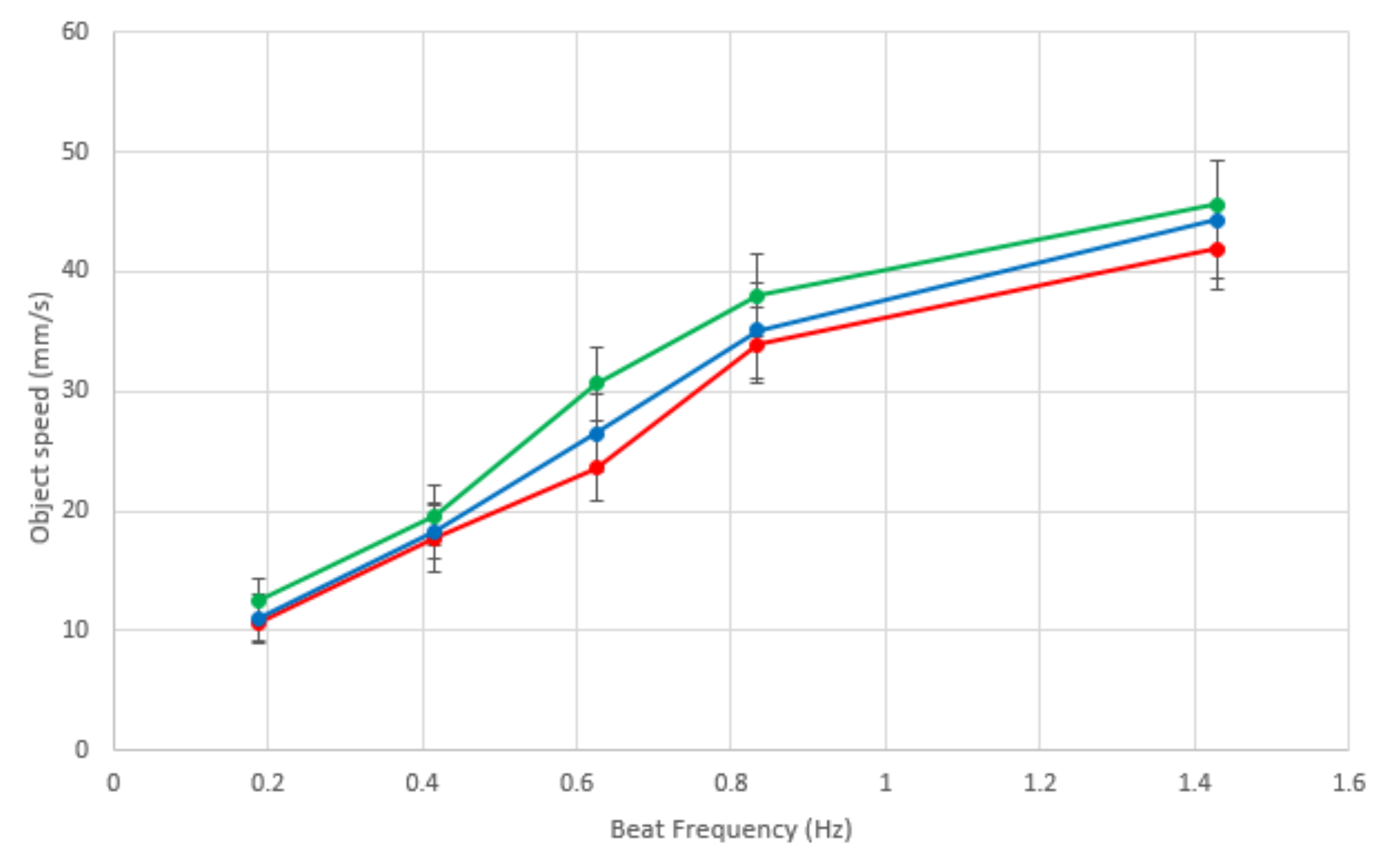
Investigating the beat frequency and the speed of locomotion of sorted objects, we tested objects moving at four different ciliary beat frequencies, those with periods of 0.7, 1.2, 1.6, 2.4 and 5.3 s. The object that was moved was a 20 × 15 cm jiffy bag with small electronic components inside, something typical of a mail sorting room situation; three different colors of jiffy bags were used to enable sorting in different directions, and all weighed the same (127 g) to eliminate bias between parcels. Each color parcel sorting was repeated five times, and the parcel was placed in the middle of the ciliary surface for each repeat.
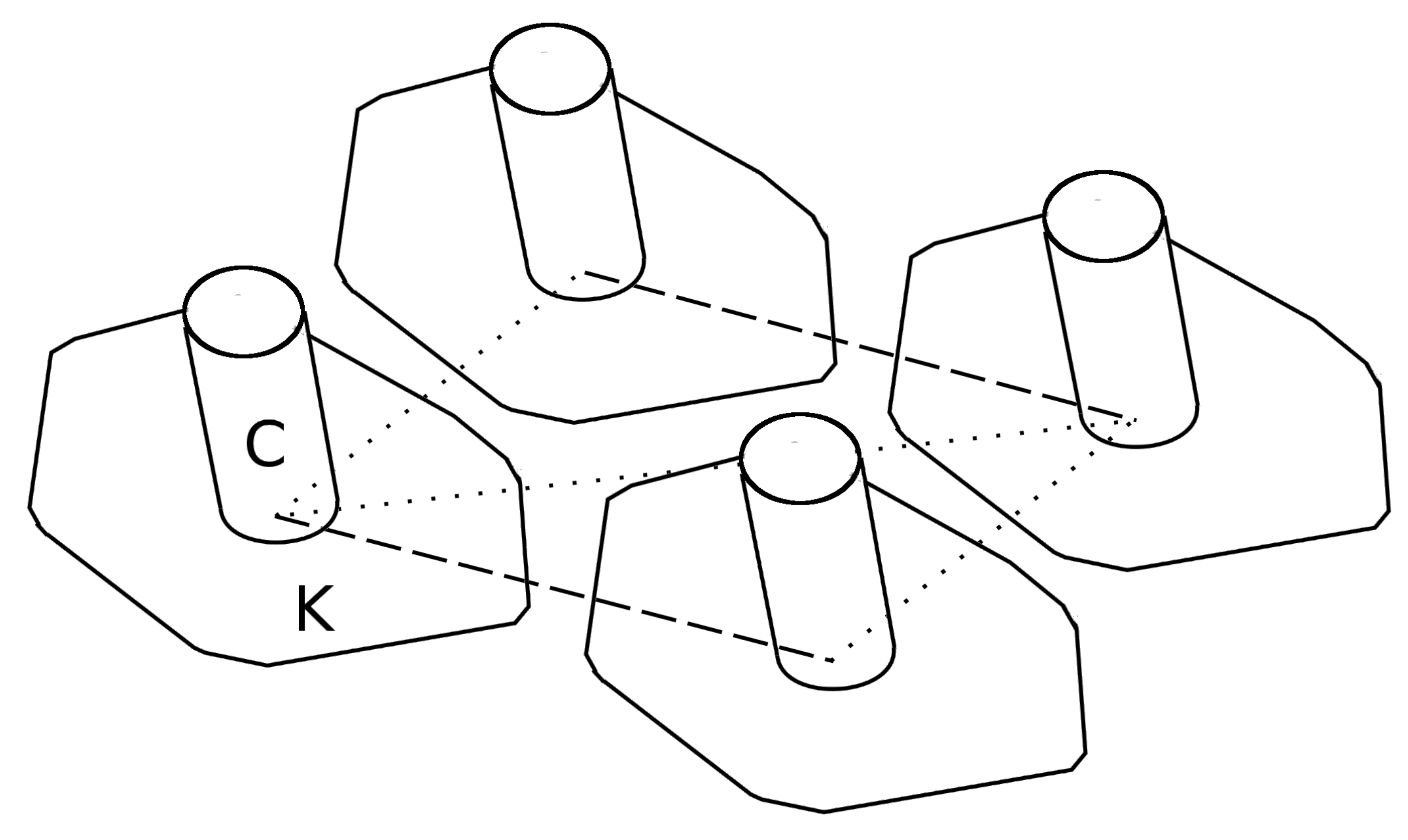
In addition to sharing power, each circuit board has a connection to each of its neighbors. Much like cellular automaton, which uses neighbor states to sequentially evolve behavior, the states of each neighbor cilia can be detected. This feature facilitates the ability to produce the familiar ciliary metachronal wave. The nature of cilia-to-cilia connections is incompletely elucidated, although they are physically linked by parallel fibrils running through the cell cortex, kinetodesma, chemically linked propagating ionotropic signals and also hydrodynamically coupled to each other via the fluid movements they induce [
16,
17]. We mimicked these forms of connectivity with local neighbor connections. Each cilia Printed Circuit Board (PCB) has an output, to announce its state to all six neighbors, and six inputs with which it detects each of its neighbor states. We have programed the embedded software in such a way to reproduce the metachronal waves; using the detection of an active neighbor cilia to initiate a phase lagged power stroke. This lagged ciliary beating action creates the metachronal wave.
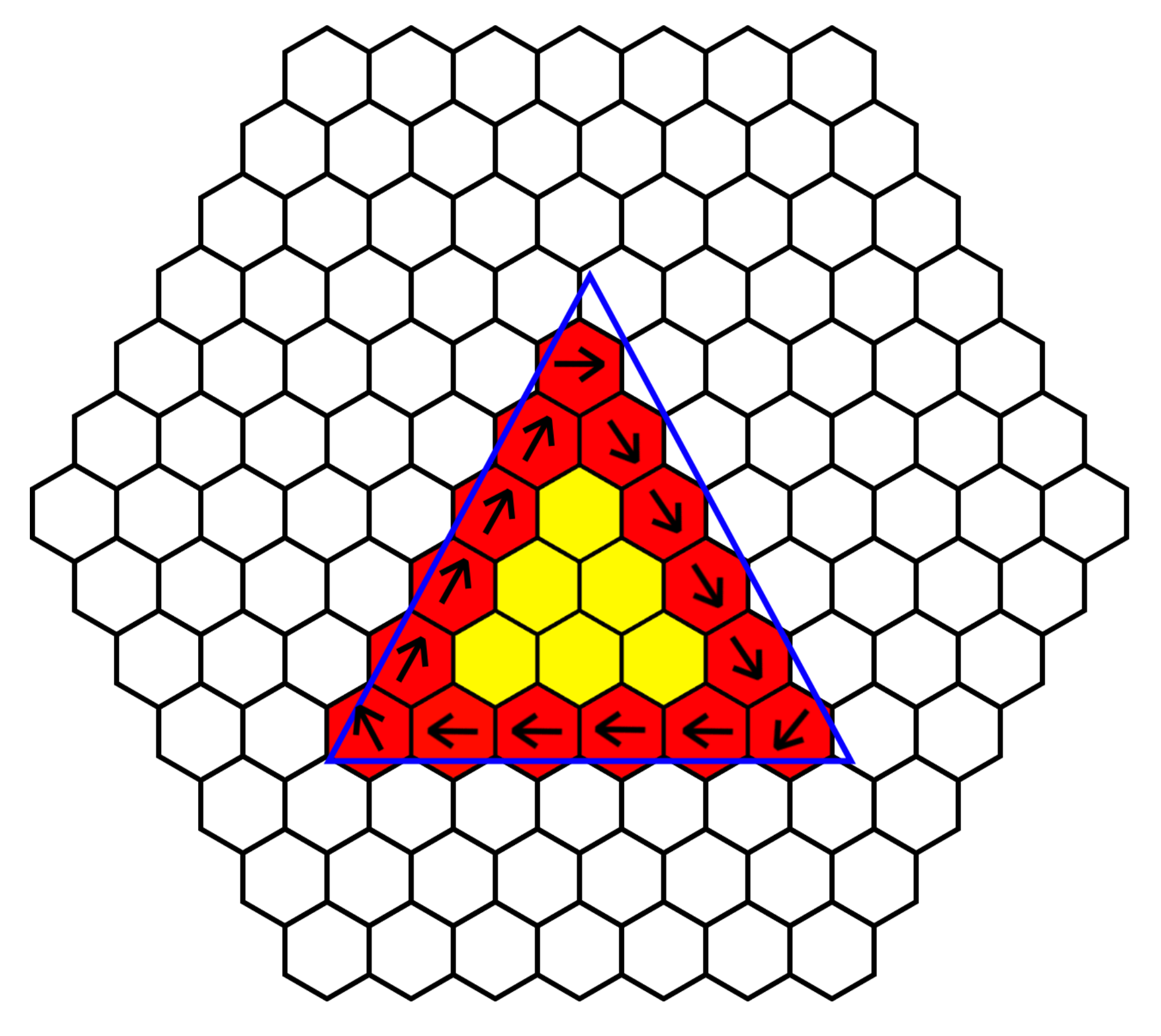
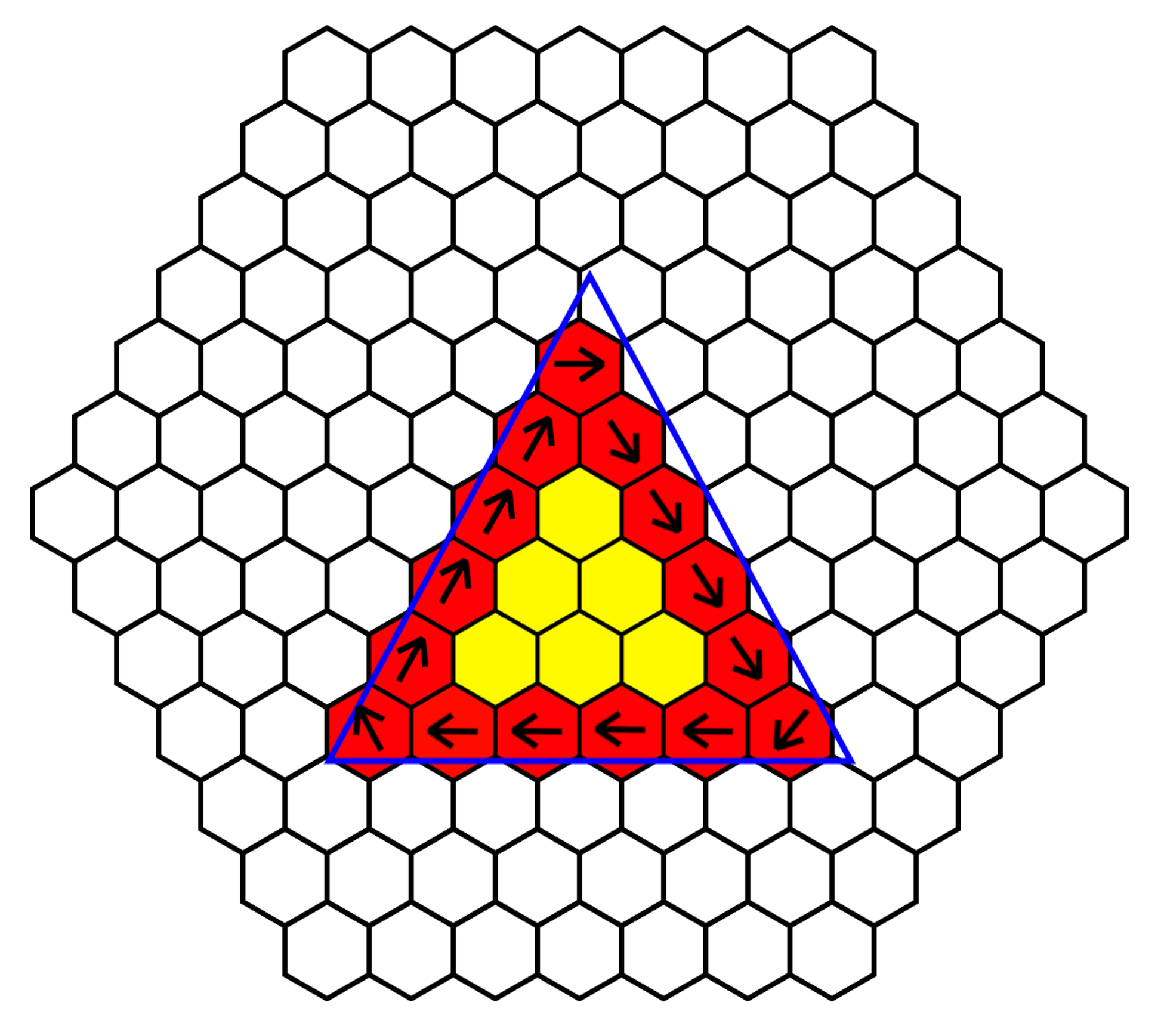
In order to detect and sort based on object shape, the sorting platform must be able to identify different shapes. This can be most accurately performed with edge detection. As each cilia cell is able to communicate and share information with its local neighbor, it is able to determine which of its surrounding cells are “occupied”. Using the ability to detect colored objects above its own headspace, each cell knows if an object is above it, then by sharing this information with its neighbor as shown in
Figure 6, it is able to determine if it is part of an edge of an object or part of an inner cell. Cells with at least one or more neighbors that are not “occupied” form part of the edge. If all cells in the platform perform this check, then the surface is able to identify the outline of an object.
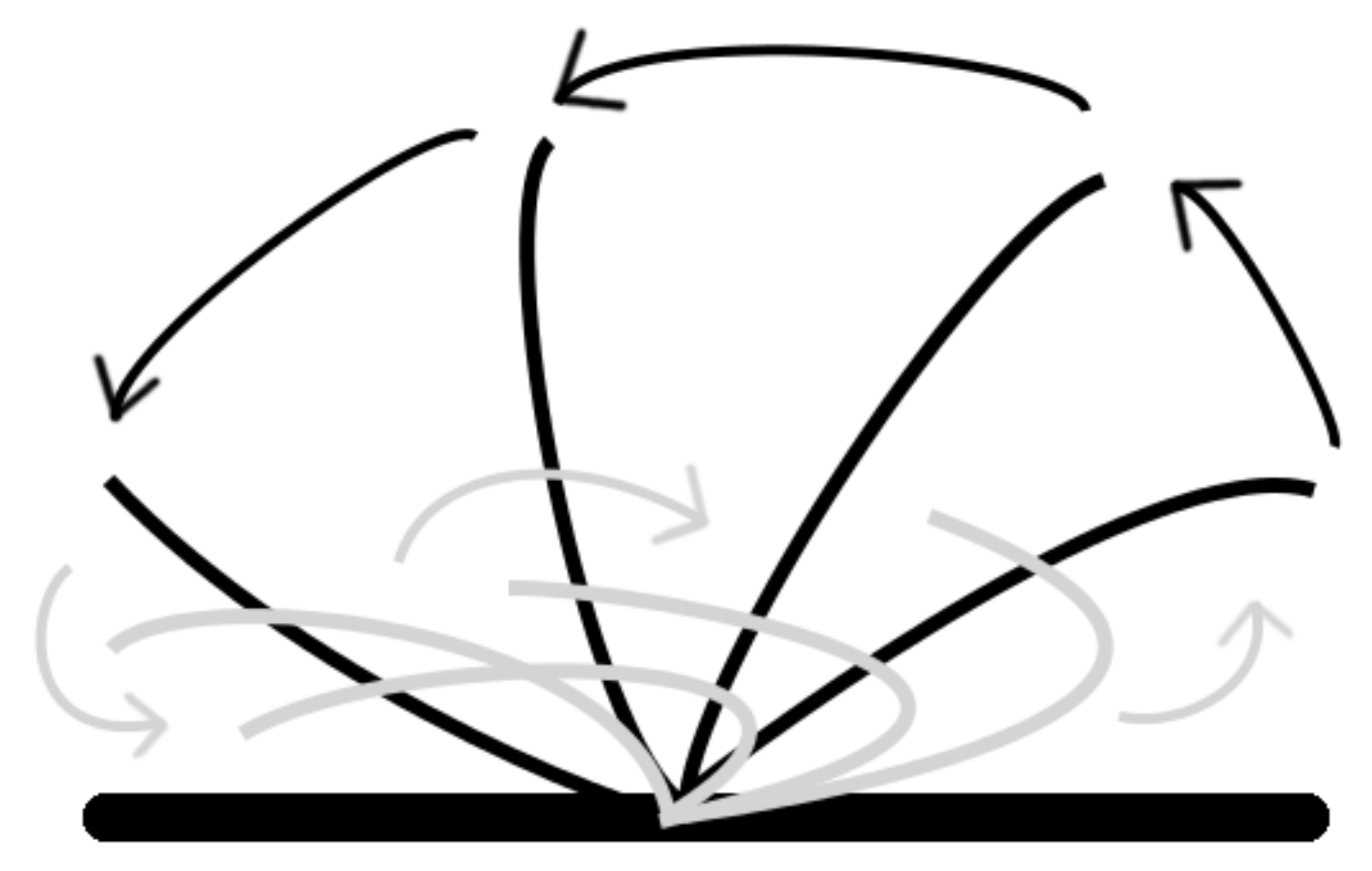
Rotation of an object is another task that can be performed by this platform. In order to perform rotation of an object about its origin, computation must be performed in order to determine either the outer edge or the center mass. Since we have neighbor communication and the center mass of a larger object could be buried several cells from the edge, we must determine the edge of an object. As we have discussed, we are able to determine the edge of a shape, by using the “occupied” status of each cell and its local neighbor, and we then take this one step further by programing in ciliary behavior when the cell is part of an edge. To rotate an object, all edge cell’s ciliary action must be coordinated; an edge cell was programed to beat the ciliary arm in a direction perpendicular to the edge and at clockwise orientation to the edge. Edge angle is calculated by averaging the local “occupied” neighbors.
Figure 7 demonstrates the process, giving several examples of this behavior. Cells in the center of the object do not respond, as the edge cells’ ciliary beating is sufficient to rotate the object.
2.2.4. Shape Detection
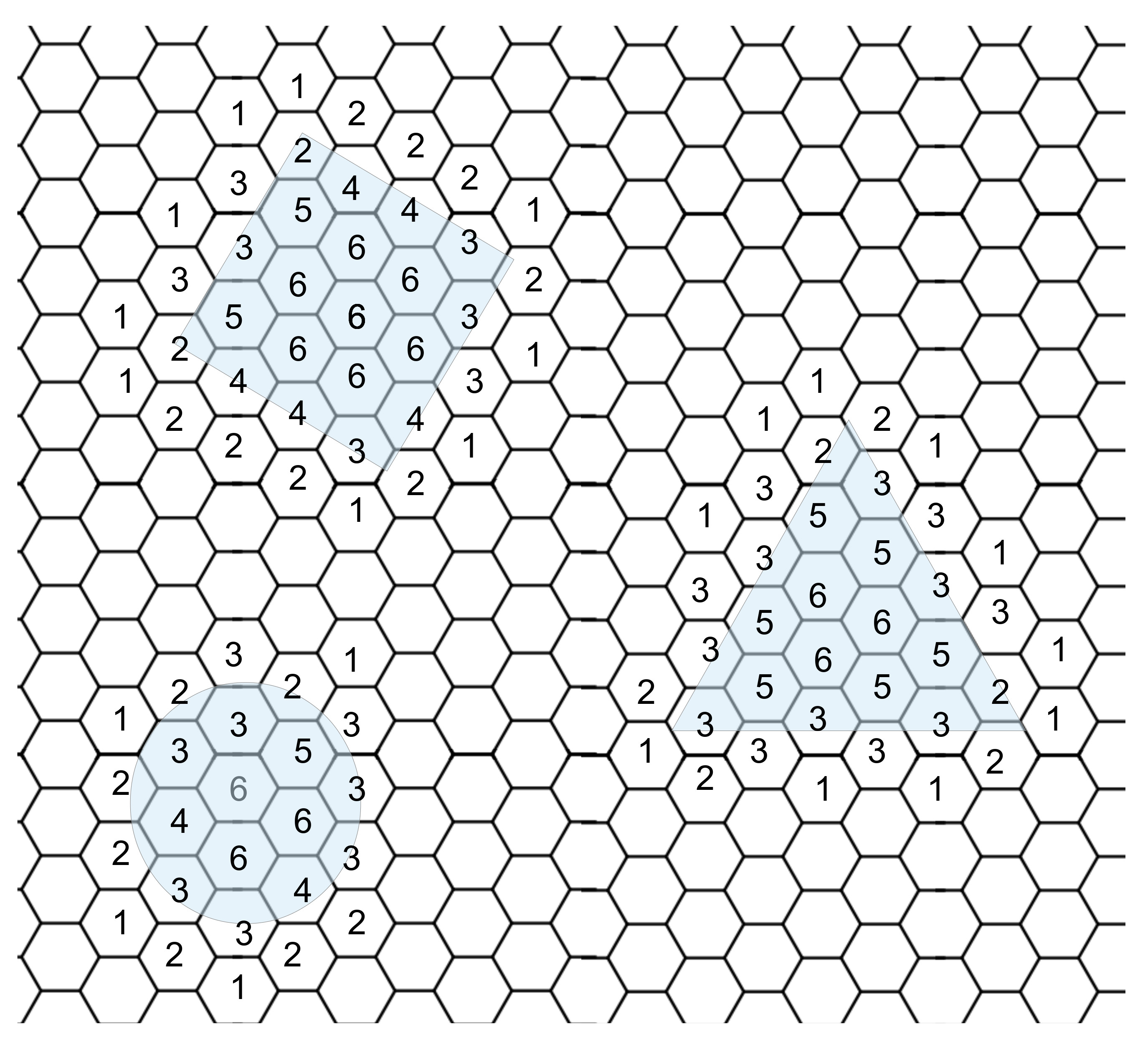
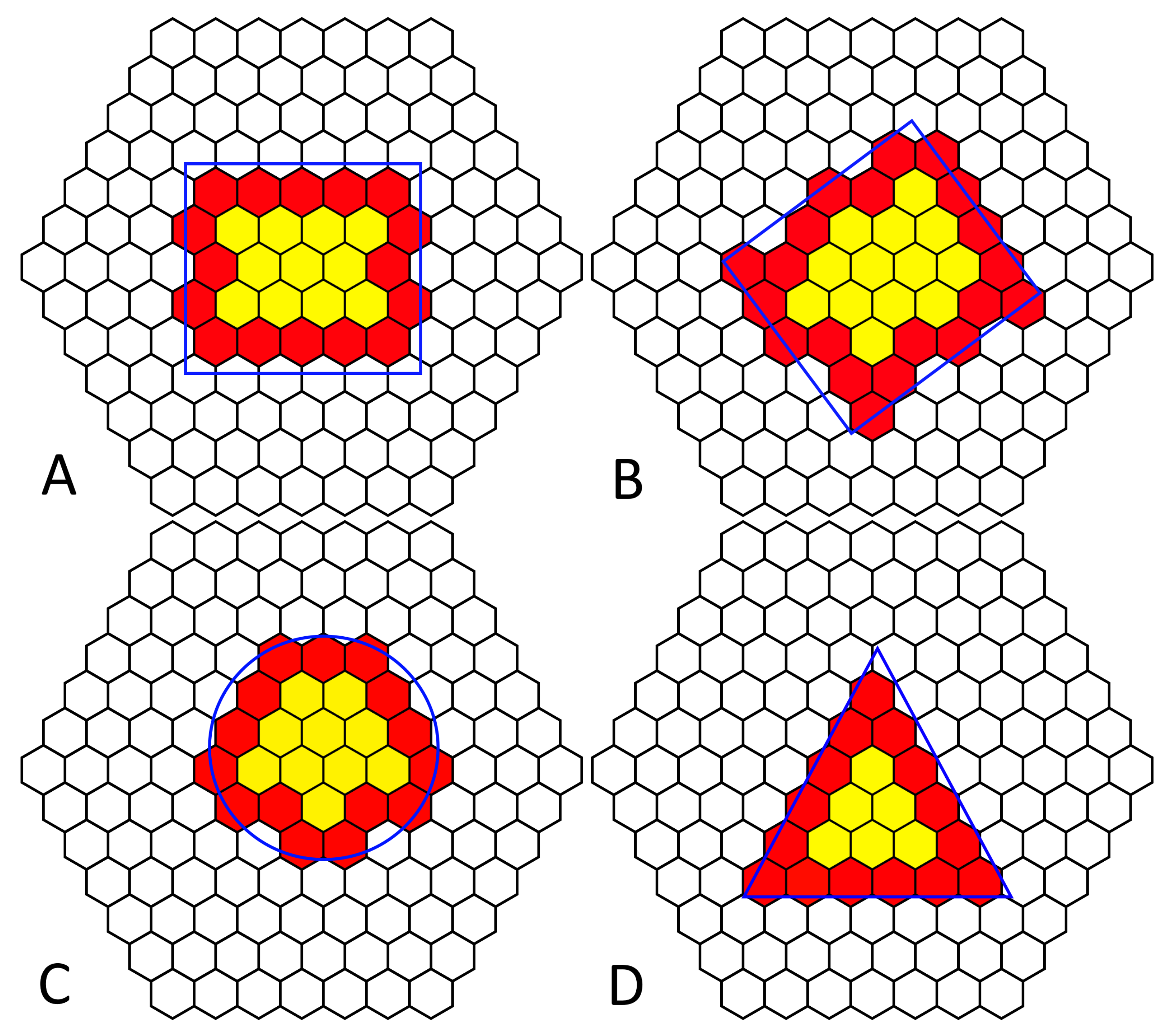
We attempted to identify an object’s shape using local communication and neighbors’ states. While we can detect the edge of an object, determining the shape provided much more of a challenge. Since each cell knows what state its neighbor is in and, importantly, what direction each neighbor is in, we can determine the number of active cells around each cell; using this information, we created a heat map of cells with the color proportional to the number of active neighbor cells. However, there was a limitation of sharing the neighbor states with anything but the local neighbors; increasing the local communication from each neighbor to include the neighboring cell’s neighbors increases the number of cells from 6 to 18, a three-fold increase in communication and geometric computational complexity. In another attempt to determine the shape, we developed a method to identify an object’s corner and, using local communication, share the number of corners in a local area. Corners or at least significant changes in shape can be identified using the number and position of neighboring active cells.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}