Genealogy: The Tree Where History Meets Genetics

 ,
,  , and
, and 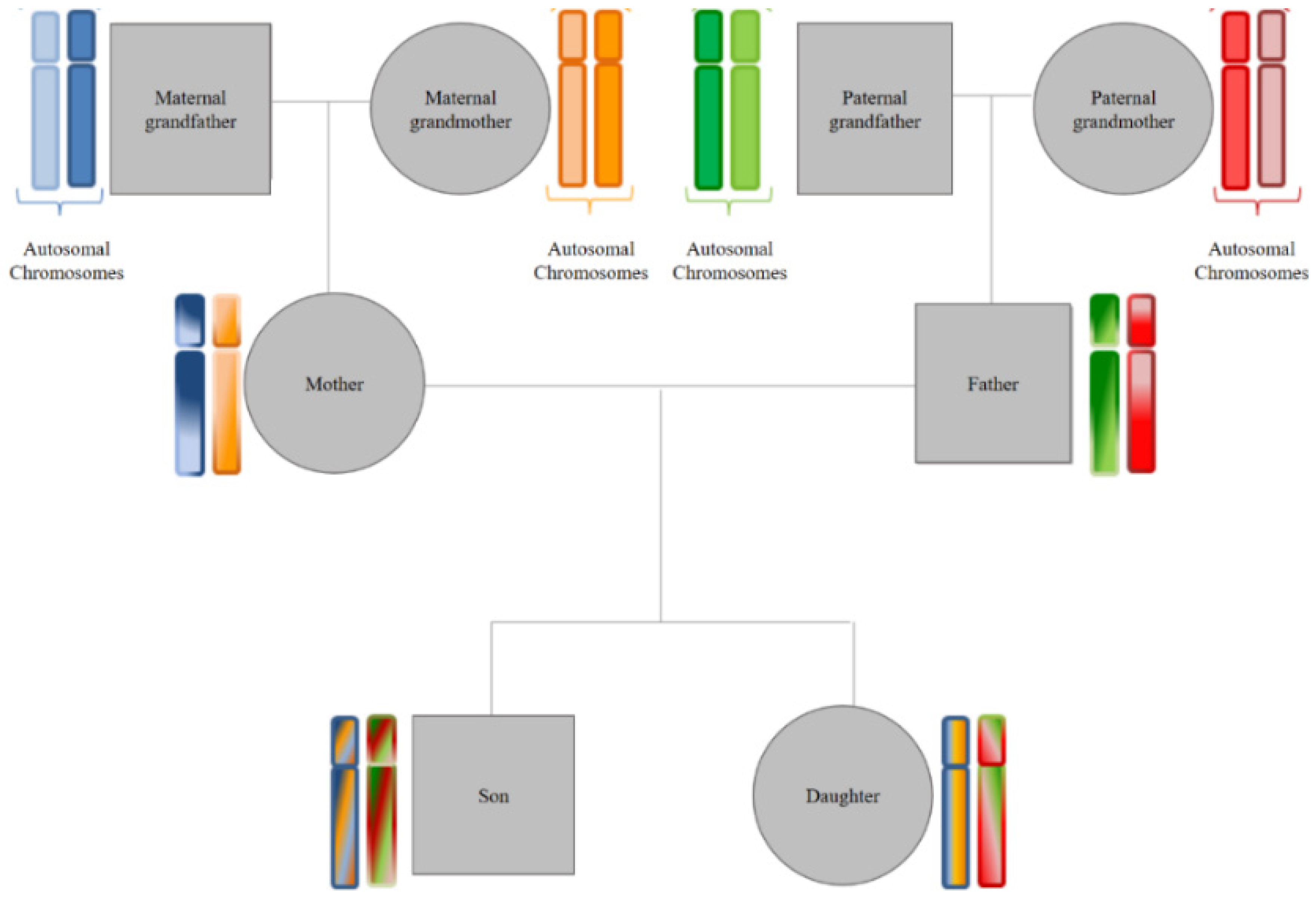{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. “Family”: The Evolution of the Concept
2.1. The Concept of Family and Relationship throughout Western European History
2.1.1. Prehistory
- (a)
- The case of Çatalhöyük (Konya, Anatolia, Turkey)—the family society
2.1.2. Ancient Period
- (a)
- Ancient Egypt (3100–30 BC)
- (b)
- Ancient Greece (1200–146 BC)
- (c)
- Jewish Culture
- (d)
- Ancient Rome (753 BC–476 AD)
- (e)
- Islamic Culture (632 AD)
2.1.3. Western Medieval Period (476 AD–1453 AD)
- (a)
- Christian Western Europe-The “Old Europe”
2.1.4. Modern Period
3. The Concept of Family, Clan, and Kinship
The Concept of Family, Kinship, and Burial Place
4. The Genetic Approach
4.1. Nuclear Information
- (a)
- Transmission of markers located on autosomal chromosomes
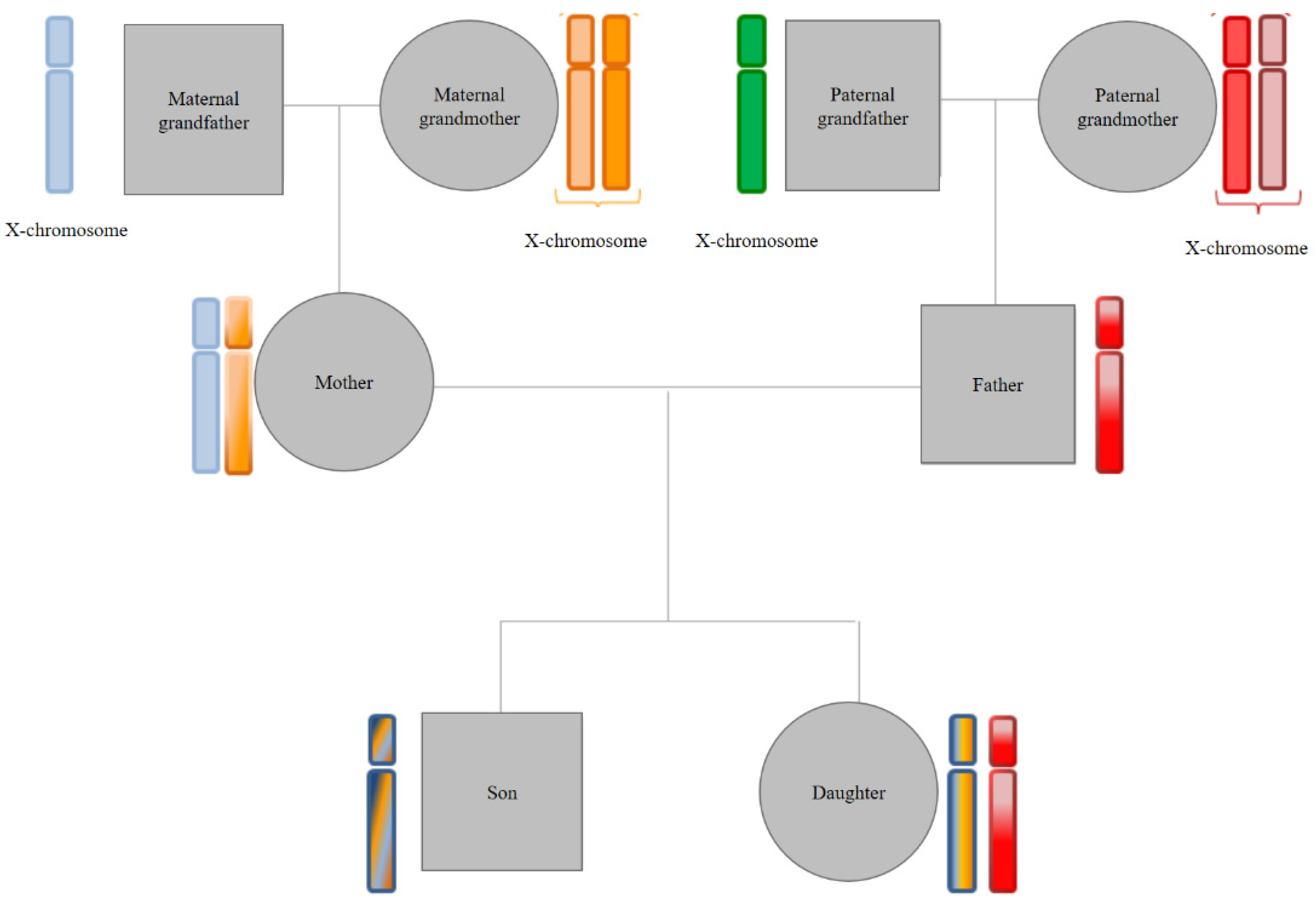
- (b)
- X chromosome transmission
4.2. Lineage Markers
- -
- The study of families
- -
- The study of lineages
- -
- The analysis of the biogeographical origin
- -
- The possibility of obtaining information from degraded samples or minimal signs; in the worst cases, these markers could provide the only available information when DNA is in very poor conditions.
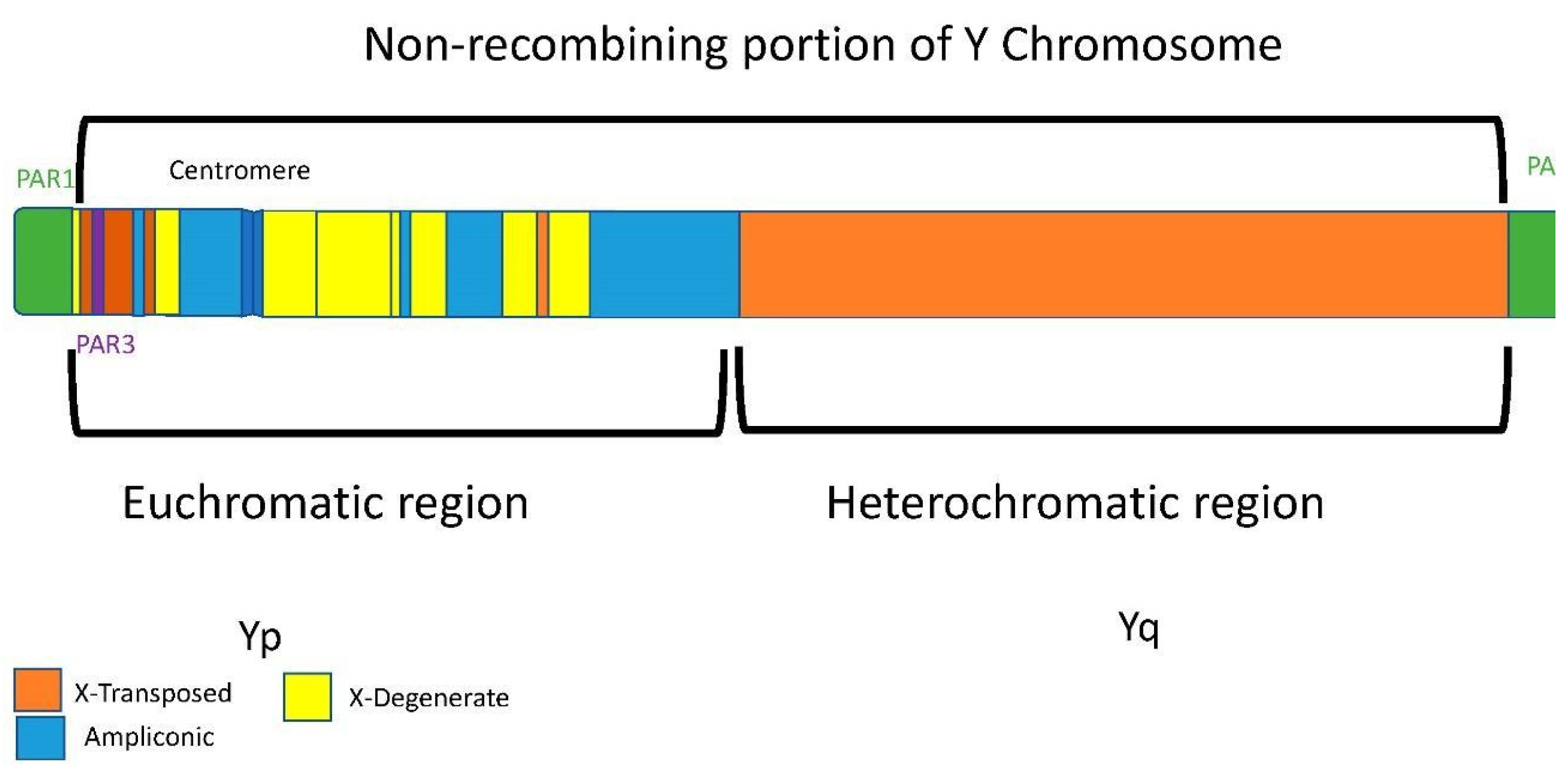
- (a)
- Y chromosome
- The Y chromosome is present exclusively in males
- The Y chromosome has no homologous chromosome
- The Y chromosome does not present recombination.
- (a1)
- Y chromosome transmission
- (a2)
- Applications of Y-Chromosome testing
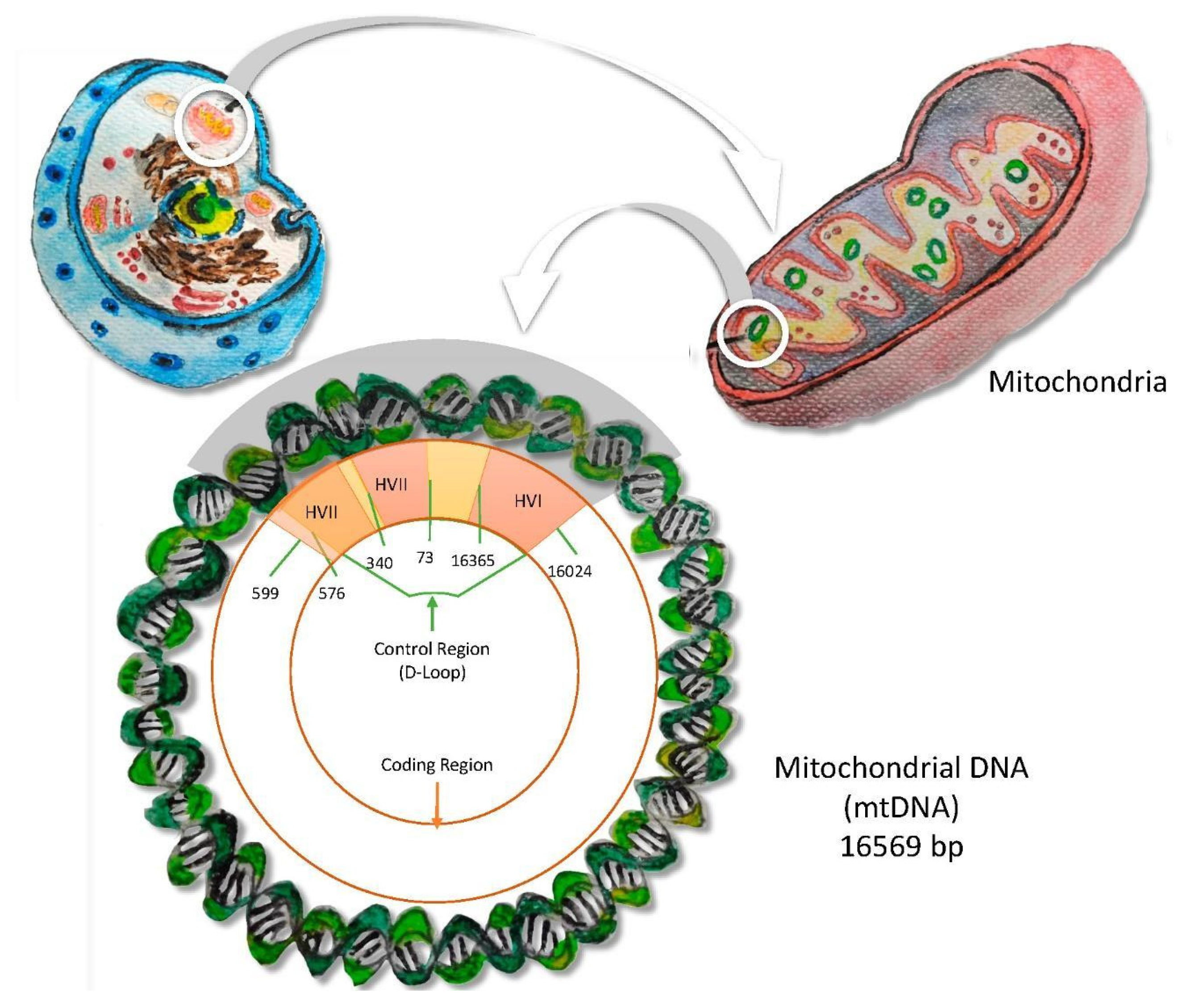
- (b)
- mtDNA
- mtDNA is inherited through the maternal lineage, from the mother to all her descendants.
- This molecule presents a recombination absence.
- The cell contains a high number of mtDNA molecules, which allow a greater probability of results obtained from problematic samples, such as human skeletal remains.
- The mtDNA shows specific geographical and ethnic distributions, involving the possibility of assigning a particular origin to an individual.
- The small size and the circular structure of mtDNA support less degradation of the molecule. Furthermore, the higher copy number compared with nuclear DNA means there are many more mtDNA molecules available for replication. This characteristic is fundamental when working with critical samples with low template DNA.
Applications of mtDNA Testing
- -
- When facing low template DNA samples, such as ancient human skeletal remains.
- -
- When the objective is to detect kinship through the maternal lineage.
- -
- To approximate the biogeographical origin.
5. Adoption and Genealogy
6. Surnames Heritage
7. Familial DNA Genealogy and Criminal Investigation
7.1. Genealogical Databases
7.1.1. GEDmatch
7.1.2. FamilyTreeDNA
7.1.3. 23AndMe
7.1.4. The Golden State Killer Case
7.1.5. Privacy and Security Implications
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Allen, Ann Taylor. 1999. Feminism, Social Science and the Meanings of Modernity: The Debate on the Origin of the Family in Europe and the United States. American Historical Review 104: 1085–113. [Google Scholar] [CrossRef]
- Allen, Troy D. 2009. The Ancient Egyptian Family. Kinship and Social Structure. New York: Taylor & Francis. ISBN 0-203-89022-1. [Google Scholar]
- Alt, Kurt, and Werner Vach. 1995. Odontological Kinship Analysis in Skeletal Remains. Forensic Science International: Genetics 74: 99–113. [Google Scholar] [CrossRef]
- Alt, Kurt, and Werner Vach. 1998. Kinship Studies in Skeletal Remains: Concepts and Examples. Dental Anthropology. London: Springer. [Google Scholar] [CrossRef]
- Anderson, Kermyt G. 2006. How Well Does Paternity Confidence Match Actual Paternity? Evidence from Worldwide Nonpaternity Rates. Current Anthropology 47: 513–20. [Google Scholar] [CrossRef]
- Andrews, Richard M., Iwona Kubacka, Patrick F. Chinnery, Robert N. Lightowlers, Douglass M. Turnbull, and Neil Howell. 1999. Reanalysis and revision of the Cambridge reference sequence for human mitochondrial DNA. Nature Genetics 23: 147. [Google Scholar] [CrossRef]
- Astuti, Rita. 2009. Revealing and obscuring rivers’s pedigrees: Biological inheritance and kinship in Madagascar. In Kinship and Beyond: The Genealogical Model Reconsidered. Fertility, Reproduction, and Sexuality. New York and Oxford: Berghahn Books. [Google Scholar]
- Bamford, Sandra. 2004. Conceiving relatedness: Non-substantial relations among the Kamea of Papua New Guinea. Journal of the Royal Anthropological Institute 10: 287–306. [Google Scholar] [CrossRef]
- Beauchamp, Tom, and James Childress. 2001. Principles of Biomedical Ethics, 5th ed. Oxford: Oxford University Press. [Google Scholar]
- Bieber, Frederick R., Charles Brenner, and David Lazer. 2006. Finding Criminals Through DNA of Their Relatives. Science 312: 1315–16. [Google Scholar] [CrossRef]
- Boattini, Alessio, Stefania Sarno, Paola Pedrini, Chiara Medoro, Carta Marilisa, Serena Tucci, Gianmarco Ferri, Milena Alù, Donata Luiselli, and Davide Pettener. 2015. Traces of medieval migrations in a socially stratified population from Northern Italy. Evidence from uniparental markers and deep-rooted pedigrees. Heredity 114: 155–62. [Google Scholar] [CrossRef]
- Brewer, Douglas J., Douglas James Brewer, and Emily Teeter. 2004. Ancient Egyptian Society and Family Life. In Egypt and the Egyptians. Cambridge: Cambridge University Press. [Google Scholar]
- Brodzinsky, David M., and Marshall D. Schechter. 1994. The Psychology of Adoption. Oxford: Oxford University Press. ISBN 978-0195082739. [Google Scholar]
- Burnham, Philip. 2015. Clan. In International Encyclopedia of the Social & Behavioral Sciences. Amsterdam: Elsevier, pp. 730–32. [Google Scholar]
- Butler, John. 2005. Forensic DNA Typing: Biology, Technology, and Genetics of STR Markers, 2nd ed. Amsterdam: Elsevier. [Google Scholar]
- Calafell, Francesc, and Maarten H. D. Lamurseen. 2017. The Y chromosome as the most popular marker in genetic genealogy benefits interdisciplinary research. Human Genetic 136: 559–73. [Google Scholar] [CrossRef]
- Carr, Karen. 2017. Ancient Greek Family–Family Relationships in Ancient Greece: Quatr.us Study Guides. Available online: https://quatr.us/greeks/women-ancient-greece.htm (accessed on 20 October 2021).
- Casey, James, and Juan Hernández-Franco. 1997. Familia, Parentesco y Linaje. Murcia: Universidad de Murcia. ISBN 8476848633. [Google Scholar]
- Claerhout, Sofie, Jennifer Roelens, Michiel Van der Haegen, Paulien Verstraete, Maarten H. D. Larmuseau, and Ronny Decorte. 2020. Ysurnames? The patrilineal Y-chromosome and surname correlation for DNA kinship research. Forensic Science International: Genetics 44: 102204. [Google Scholar] [CrossRef]
- Coble, Michael, Odile M. Loreille, Mark J. Wadhams, Suni M. Edson, and Kerry Maynard. 2009. Mystery Solved: The Identification of the Two Missing Romanov Children Using DNA Analysis. PLoS ONE 4: e4838. [Google Scholar] [CrossRef] [PubMed]
- Crespillo, Márquez, and Pedro A. Barrio. 2018. Genética Forense: Del Laboratorio a Los Tribunales. Edited by Díaz de Santos. Madrid: Ediciones Díaz de Santos, S.A. ISBN 978-84-9052-213-4. [Google Scholar]
- Debus-Sherrill, Sara, and Michael B. Field. 2018. Familial DNA searching—An emerging forensic investigative tool. Science & Justice 59: 20–28. [Google Scholar] [CrossRef]
- Diop, Cheikh Anta. 1971. Parente Genetique de L’Egyptien Pharaonique et des Lanques Negro-Africaines. Ifan-Dakar: Les Nouvelles Editions Africaines, ISBN-10: 2917400013. [Google Scholar]
- Esparza, Ángel, Sara Palomo-Díez, Javier Velasco-Vázquez, Germán Delibes, Eduardo Arroyo-Pardo, and Domingo C. Salazar-García. 2017. Familiar Kinship? Palaeogenetic and isotopic evidence from a triple burial of the Cogotas I Archaeological Culture (Bronze Age, Iberian Peninsula). Oxford Journal of Archaeology 36: 223–42. [Google Scholar] [CrossRef]
- Fielder, Christine, and Chris King. 2004. Sexual Paradox: Complementarity, Reproductive Conflict and Human Emergence. Morrisville: Lulu.com, ISBN-10: 141165532X. [Google Scholar]
- Fortier, Alyssa Lyn, Jaehee Kim, and Noah A. Rosenberg. 2020. Human-Genetic Ancestry Inference and False Positives in Forensic Familial Searching. G3 (Bethesda) 10: 2893–902. [Google Scholar] [CrossRef] [PubMed]
- Foster, Eugene A., Mark A. Jobling, Peter G. Taylor, Peter Donnelly, Peter de Knijff, Rene Mieremet, Tatiana Zerjal, and Chris Tyler-Smith. 1998. Jefferson fathered slave’s last child. Nature 396: 27–28. [Google Scholar] [CrossRef]
- Friš, Eva Lina E., Sara Grdina, Eva Podovšovnik, Tomaž Zupanc, and Irena Zupanič Pajnič. 2019. Comparison of DNA yield after longterm storage of Second World War bone samples. Forensic Science International: Genetics Supplement Series 7: 117–19. [Google Scholar]
- Gilgenkrantz, Simone. 2009. Unambiguous identification of the remains of the Romanov family. Medecine Sciences 25: 637–40. [Google Scholar] [CrossRef][Green Version]
- Gill, Peter, Pavel L. Ivanov, Colin Kimpton, Romelle Piercy, and Nicola Benson. 1994. Identification of the remains of the Romanov family by DNA analysis. Nature Genetics 6: 130–35. [Google Scholar] [CrossRef]
- Gomes, Cláudia. 2020. Investigación de Parentesco Biológico en Muestras Críticas-Utilidad en Casos de Investigación Histórica, Antropológica y/o Forense. Ph.D. thesis, Facultad de Medicina, Universidad Complutense de Madrid, Madrid, Spain. [Google Scholar]
- Gomes, Cláudia, María Fondevila, Concepción Magaña-Loarte, Juan Fernández-Jiménez, José Fernández-Serrano, and Sara Palomo-Díez. 2019. An unusual kinship case from the Spanish Civil War (1936–1939): Ancient versus degraded sample’s investigation. Forensic Science International: Genetics Supplement Series 7: 690–91. [Google Scholar] [CrossRef]
- Goucher, Candice, and Linda Walton. 1998. Ordering the World: Family and Household, in the Balance: Themes in Global History. Boston: McGraw-Hill. ISBN 0070241791. [Google Scholar]
- Graves, Jennifer A. M. 1998. Interactions between SRY and SOX genes in mammalian sex determination. Bioessays 20: 264–69. [Google Scholar] [CrossRef]
- Greeff, Jaco M., and Christoff Erasmus. 2015. Three hundred years of low non-paternity in a human population. Heredity 115: 396–404. [Google Scholar] [CrossRef] [PubMed]
- Greely, Henry T., Daniel P. Riordan, Garrison Nanibaa’A, and Joanna L. Mountain. 2006. Family ties: The use of DNA offender databases to catch offenders’ kin. Journal of Law, Medicine & Ethics 34: 248–62. [Google Scholar]
- Gutrie, R. Dale. 2005. The Nature of Paleolithic art. Chicago, 1st ed. Chicago: University of Chicago Press. ISBN 0226311260. [Google Scholar]
- Haag, Matthew. 2019. FamilyTreeDNA Admits to Sharing Genetic Data with FBI. The New York Times, July 9. [Google Scholar]
- Haas, Randall, James Watson, Tammy Buonasera, John Southon, Jennifer C. Chen, Sarah Noe, Kevin Smith, Carlos Viviano Llave, Jelmer Eerkens, and Glendon Parker. 2020. A 9000-year-old hunter burial and meta-analysis reveal nongendered labor in the early Americas with females as big-game hunters. Science Advances 6: eabd0310. [Google Scholar] [CrossRef]
- Harper, Nathan, and Tiffiny Tung. 2012. Burial Treatment Based on Kinship? The Hellenistic–Roman and Venetian- Period Tombs in the Malloura Valley. In Crossroads and Boundaries: The Archaeology of Past and Present in the Malloura Valley, Cyprus, 2nd ed. Alexandria: American Schools of Oriental Research. [Google Scholar]
- Herper, Matthew. 2013. 23andMe Stops Offering Genetic Tests Related To Health. Forbes. Available online: https://www.forbes.com/sites/matthewherper/2013/12/05/23andme-stops-offering-genetic-tests-related-to-health/ (accessed on 15 July 2021).
- Holmes, Teresa. 2009. When blood matters: Making kinship in colonial Kenya. In Kinship and Beyond: The Genealogical Model Reconsidered. Fertility, Reproduction and Sexuality. New York: Berghahn Books. [Google Scholar]
- Hummel, Susanne. 2003. Ancient DNA Typing. Methods, Strategies and Applications, 1st ed. New York: Springer. [Google Scholar]
- Jehaes, Els, Ronny Decorte, Alain Peneau, and Johan H. Petrie. 1998. Mitochondrial DNA analysis on remains of a putative son of Louis XVI, King of France and Marie-Antoinette. European Journal of Human Genetics 6: 383–95. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jobling, Mark A., and Chris Tyler-Smith. 2003. The human Y chromosome: An evolutionary marker comes of age. Nature Reviews Genetics 4: 598–612. [Google Scholar] [CrossRef]
- Jobling, Mark A., Matthew Hurles, and Chris Tyler-Smith. 2004. Human Evolutionary Genetics: Origins, Peoples and Disease. New York: Garland Science. ISBN 0815341857. [Google Scholar]
- Johnson, Kent M., and Kathleen S. Paul. 2016. Bioarchaeology and Kinship: Integrating Theory, Social Relatedness, and Biology in Ancient Family Research. Journal of Archaeological Research 24: 75–123. [Google Scholar] [CrossRef]
- Keyser-Tracqui, Christine, Eric Crubezy, and Bertrand Ludes. 2003. Nuclear and mitochondrial DNA analysis of a 2000-year-old necropolis in the Egyin Gol Valley of Mongolia. The American Journal of Human Genetics 73: 247–60. [Google Scholar] [CrossRef]
- King, Turi E., and Mark A. Jobling. 2009. Founders, drift, and infidelity: The relationship between Y chromosome diversity and patrilineal surnames. Molecular Biology Evolution 26: 1093–102. [Google Scholar] [CrossRef]
- King, Turi E., Stéphane J. Ballereau, Kevin E. Schurer, and Mark A. Jobling. 2006. Genetic signatures of coancestry within surnames. Current Biology 16: 384–88. [Google Scholar] [CrossRef]
- Klapisch-Zuber, Christiane. 1991. The Genesis of the Family Tree. I Tatti Studies: Essays in the Renaissance. Chicago: The University of Chicago Press, vol. 4, pp. 105–29. [Google Scholar]
- Klapisch-Zuber, Christiane. 2000. L’ombre des Ancêtres: Essai sur L’imaginaire Médiéval de la Parenté. Paris: Fayard, ISBN-10: 2213604274. [Google Scholar]
- Kling, Daniel, Christopher Phillips, Debbie Kennett, and Andreas Tillmar. 2021. Investigative genetic genealogy: Current methods, knowledge and practice. Forensic Science International: Genetics 52: 102474. [Google Scholar] [CrossRef] [PubMed]
- Lambek, Michael. 2011. Kinship as gift and theft: Acts of succession in Mayotte and Ancient Israel. American Ethnologist 38: 2–16. [Google Scholar] [CrossRef]
- Larmuseau, Maarten H. D., Nancy Vanderheyden, Anneleen Van Geystelen, Mannis Van Oven, Peter de Knijff, and Ronny Decorte. 2014. Recent radiation within Y-chromosomal haplogroup R-M269 resulted in high Y-STR haplotype resemblance. Annals of Human Genetics 78: 92–103. [Google Scholar] [CrossRef]
- Larmuseau, Maarten H. D., Koen Matthijs, and Tom Wenseleers. 2016. Cuckolded fathers rare in human populations. Trends in Ecology & Evolution 31: 327–29. [Google Scholar] [CrossRef]
- Leach, James. 2004. Creative Land: Place and Procreation on the Rai Coast of Papua New Guinea. Oxford: Berghahn Books, ISBN-10: 1571816933. [Google Scholar]
- Lévi-Strauss, Claude. 1962. Le Totemisme Aujourd’hui. Paris: Presses Universitaires de France. [Google Scholar]
- Lévi-Strauss, Claude. 1984. Paroles Données. Paris: Plon, ISBN-10: 2259011373. [Google Scholar]
- Lévi-Strauss, Claude. 1987. Anthropology and Myth: Lectures 1951–1982. New York: Wiley-Blackwell. ISBN 978-0-631-14474-8. [Google Scholar]
- Lévi-Strauss, Claude. 1999. The Way of the Masks. Vancouver: University of British Columbia Press. ISBN 9780774807616. [Google Scholar]
- Lindsay, Hugh. 2009. Adoption in the Roman World. Cambridge: Cambridge University Press. [Google Scholar] [CrossRef]
- López-Parra, Ana María, Maria S. Mesa, Leonor Gusmão, António Amorim, and Eduardo Arroyo-Pardo. 2005. Isonimia y Genética: Variabilidad geográfica de los individuos apellidados Castilla. Revista Española de Antropología Física 25: 87. [Google Scholar]
- Marshall, Charla, Rebecca Taylor, Kimberly Sturk-Andreaggi, Suzanne Barritt-Ross, Gregory E. Berg, and Timothy P. McMahon. 2020. Mitochondrial DNA haplogrouping to assist with the identification of unknown service members from the World War II Battle of Tarawa. Forensic Science International: Genetics 47: 102291. [Google Scholar] [CrossRef]
- Martínez-Cadenas, Conrado, Alejandro Blanco-Verea, Barbara Hernando, George B. J. Busby, María Brion, and Ángel Carracedo. 2016. The relationship between surname frequency and Y chromosome variation in Spain. European Journal of Human Genetics 24: 120–28. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Gonzalez, Luis J., Esther Martinez-Espín, Juan C. Alvarez, Francesc Albardaner, Olga Rickards, and Cristina Martinez-Labarga. 2012. Surname and Y chromosome in Southern Europe: A case study with Colom/Colombo. European Journal of Human Genetics 20: 211–16. [Google Scholar] [CrossRef][Green Version]
- Mateen, Rana Muhammad, Muhammad Farooq Sabar, Safdar Hussain, Rukhsana Parveen, and Manzoor Hussain. 2020. Familial DNA analysis and criminal investigation: Usage, Downsides and Privacy concerns. Forensic Science International 318: 110576. [Google Scholar] [CrossRef]
- Maynes, Mary Jo, and Ann Waltner. 2012. The Family A World History (New Oxford World History). New York: Oxford University Press, ISBN-10: 0195338146. [Google Scholar]
- McEvoy, Brian, and Daniel Bradley. 2006. Y-chromosome and the extent of patrilineal ancestry in Irish surnames. Human Genetics 119: 212–9. [Google Scholar] [CrossRef]
- McGlynn, Kayleigh E. 2019. Remedying wrongful convictions through DNA testing: Expanding post-conviction litigants’ access to DNA database searches to prove innocence. BCL Review 60: 709–51. [Google Scholar]
- Mitchell, Linda Elizabeth. 2007. Family Life in Middle Ages. Westport: Greenwood Publishing Group. ISSN 1558-6286. [Google Scholar]
- Moulton, Carroll. 1998. Ancient Greece and Rome: An Encyclopedia for Students, 1st ed. New York: Charles Scribners & Sons, p. 169. ISBN 0684805073. [Google Scholar]
- Mulligan, Connie J. 2006. Anthropological applications of ancient DNA: Problems and prospects. American Antiquity 71: 365–80. [Google Scholar] [CrossRef]
- Murray, Jacqueline. 2017. The Family in the Medieval West. Oxford: Oxford Bibliographies. [Google Scholar] [CrossRef]
- O’Connor, Stephen. 2004. Orphan Trains: The Story of Charles Loring Brace and the Children He Saved and Failed. Chicago: University of Chicago Press. ISBN 9780226616674. [Google Scholar]
- Obal, Marcel, Irena Zupanič Pajnič, Barbara Gornjak Pogorelc, and Tomaž Zupanc. 2019. Different skeletal elements as a source of DNA for genetic identification of Second World War victims. Forensic Science International: Genetics Supplement Series 7: 27–29. [Google Scholar] [CrossRef]
- Ochiai, Eriko, Motoki Osawa, Shiori Satoh, Tomonori Tamura, Masato Nakatome, Yu Kaneko, Yu Kakimoto, and Kiyoshi Minaguchi. 2021. Y chromosome analysis for common surnames in the Japanese male population. Japanese Journal of Human Genetics 66: 731–38. [Google Scholar] [CrossRef]
- Ohno, Susumu. 1967. Monographs on Endocrinology. In Sex Chromosomes and Sex-Linked Genes, 1st ed. Heidelberg: Springer. [Google Scholar]
- Olalde, Iñigo, Federico Sánchez-Quinto, Debayan Datta, Urko M. Marigorta, Charleston W. K. Chiang, and Juan Antonio Rodríguez. 2014. Genomic analysis of the blood attributed to Louis XVI (1754–1793), king of France. Scientific Reports 4: 1–7. [Google Scholar] [CrossRef]
- Palomo-Díez, Sara. 2015. Caracterización genética de las poblaciones de las Edades del Cobre y del Bronce en la Submeseta Norte de la Península Ibérica. Ph.D. thesis, Universidad Complutense de Madrid, Madrid, Spain. [Google Scholar]
- Palomo-Díez, Sara, Ana María López-Parra, Cláudia Gomes, Carlos Baeza-Richer, Ángel Esparza-Arroyo, and Eduardo Arroyo-Pardo. 2015. Kinship analysis in mass graves: Evaluation of the Blind Search tool of the Familias 3.0 software in critical samples. Forensic Science International: Genetics Supplement Series 5: e547–e550. [Google Scholar] [CrossRef]
- Palomo-Díez, Sara, Ángel Esparza Arroyo, Mirian Tirado-Vizcaíno, Javier Velasco Vázquez, Ana María López-Parra, Cláudia Gomes, Carlos Baeza-Richer, and Eduardo Arroyo-Pardo. 2018. Kinship analysis and allelic dropout: A forensic approach on an archaeological case. Annals of Human Biology 45: 365–68. [Google Scholar] [CrossRef] [PubMed]
- Palomo-Díez, Sara, Cláudia Gomes, Ana María López-Parra, Carlos Baeza-Richer, and Ivón Cuscó. 2019. Genetic identification of Spanish civil war victims. The state of the art in Catalonia (Northeastern Spain). Forensic Science International: Genetics Supplement Series 7: 419–21. [Google Scholar] [CrossRef]
- Pinheiro, María de Fátima Terra. 2010. Genética Forense: Perspectivas da Identificação Genética. Porto: Edições Universidad Fernando Pessoa. ISBN 978-989-643-050-4. [Google Scholar]
- Pollack, Andrew. 2015. 23andMe Will Resume Giving Users Health Data. The New York Times. Available online: https://www.nytimes.com/2015/10/21/business/23andme-will-resume-giving-users-health-data.html (accessed on 15 July 2021).
- Sasse, George, Hansjakob Muller, Ranajit Chakrabory, and Jürg Ott. 1994. Estimating the frequency of nonpaternity in Switzerland. Human Heredity 44: 337–43. [Google Scholar] [CrossRef]
- Schneider, David. 1972. What Is Kinship All About? Washington, DC: The Anthropological Society of Washington. [Google Scholar]
- Schneider, David. 1980. American Kinship: A Cultural Account. Chicago: University of Chicago Press. ISBN 9780226227092. [Google Scholar]
- Skaletsky, Helen, Tomoko Kuroda-Kawaguchi, Patrick J. Minx, Holland S. Cordum, and LaDeana Hillier. 2003. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 423: 825–37. [Google Scholar] [CrossRef]
- Stojanowski, Christopher M., and Michael A. Schillaci. 2006. Phenotypic Approaches for Understanding Patterns of Intracemetery Biological Variation. American Journal of Physical Anthropology 43: 49–88. [Google Scholar] [CrossRef]
- Sykes, Bryan, and Catherine Irven. 2000. Surnames and the Y chromosome. American Journal of Human Genetics 66: 1417–19. [Google Scholar] [CrossRef]
- TIME. 2008. Best Inventions of the Year. Available online: http://content.time.com/time/specials/packages/completelist/0,29569,1852747,00.html (accessed on 19 July 2021).
- Underhill, Peter A., Peidong Shen, Alice A. Lin, Li Jin, Giuseppe Passarino, and Wei H. Yang. 2000. Y chromosome sequence variation and the history of human populations. Nature Genetics 26: 358–61. [Google Scholar] [CrossRef] [PubMed]
- Van Oven, Mannis, and Mandred Kayser. 2009. Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation. Human Mutation 30: E386–E394. [Google Scholar] [CrossRef] [PubMed]
- Viveiros de Castro, Eduardo. 2009. The gift and the given: Three nano-essays on kinship and magic. In Kinship and Beyond: The Genealogical Model Reconsidered. Fertility, Reproduction and Sexuality. London: Berghahn Books. [Google Scholar]
- Whitehouse, Ruth. 2006. Gender Archaeology in Europe. Handbook of Gender in Archaeology. Lanham: Rowman Altamira, p. 744. ISBN 0759106789. [Google Scholar]
- Wickenheiser, Ray. 2019. Forensic genealogy, bioethics and the Golden State Killer case. Forensic Science International: Synergy 1: 114–25. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, John F. 2006. Unraveling male and female histories from human genetic data. Current Opinion in Genetics & Development 16: 611–17. [Google Scholar] [CrossRef]
- Williams, Robin, and Paul Johnson. 2005. Issues in the developing uses of DNA profiling in support of criminal. The Journal of Law Medicine & Ethics 33: 545–58. [Google Scholar]
- Zeng, Zhen, Jiaoyang Tian, Chuangui Jiang, Weijian Ye, Kaijun Liu, and Yuchun Li. 2019. Inferring the history of surname Ye based on Y chromosome high-resolution genotyping and sequencing data. Journal of Human Genetics 64: 703–9. [Google Scholar] [CrossRef]
- Zhang, Sarah. 2019. The Messy Consequences of the Golden State Killer Case. The Atlantic. Available online: https://www.theatlantic.com/science/archive/2019/10/genetic-genealogy-dna-database-criminal-investigations/599005/ (accessed on 10 July 2021).
- Zupanič Pajnič, Irena, Magdalena Debska, Barbara Gornjak Pogorelc, Katja Vodopivec Mohorčič, Jože Balažic, Tomaž Zupanc, Borut Štefanič, and Ksenija Geršak. 2016. Highly efficient automated extraction of DNA from old and contemporary skeletal remains. Journal of Forensic and Legal Medicine 37: 78–86. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, C.; Palomo-Díez, S.; López-Parra, A.M.; Arroyo-Pardo, E. Genealogy: The Tree Where History Meets Genetics. Genealogy 2021, 5, 98. https://doi.org/10.3390/genealogy5040098
Gomes C, Palomo-Díez S, López-Parra AM, Arroyo-Pardo E. Genealogy: The Tree Where History Meets Genetics. Genealogy. 2021; 5(4):98. https://doi.org/10.3390/genealogy5040098
Chicago/Turabian StyleGomes, Cláudia, Sara Palomo-Díez, Ana María López-Parra, and Eduardo Arroyo-Pardo. 2021. "Genealogy: The Tree Where History Meets Genetics" Genealogy 5, no. 4: 98. https://doi.org/10.3390/genealogy5040098
APA StyleGomes, C., Palomo-Díez, S., López-Parra, A. M., & Arroyo-Pardo, E. (2021). Genealogy: The Tree Where History Meets Genetics. Genealogy, 5(4), 98. https://doi.org/10.3390/genealogy5040098