



Effects of Nutrition and Light Quality on the Growth of Southern Highbush Blueberry (Vaccinium corymbosum L.) in an Advanced Plant Factory with Artificial Lighting (PFAL)

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
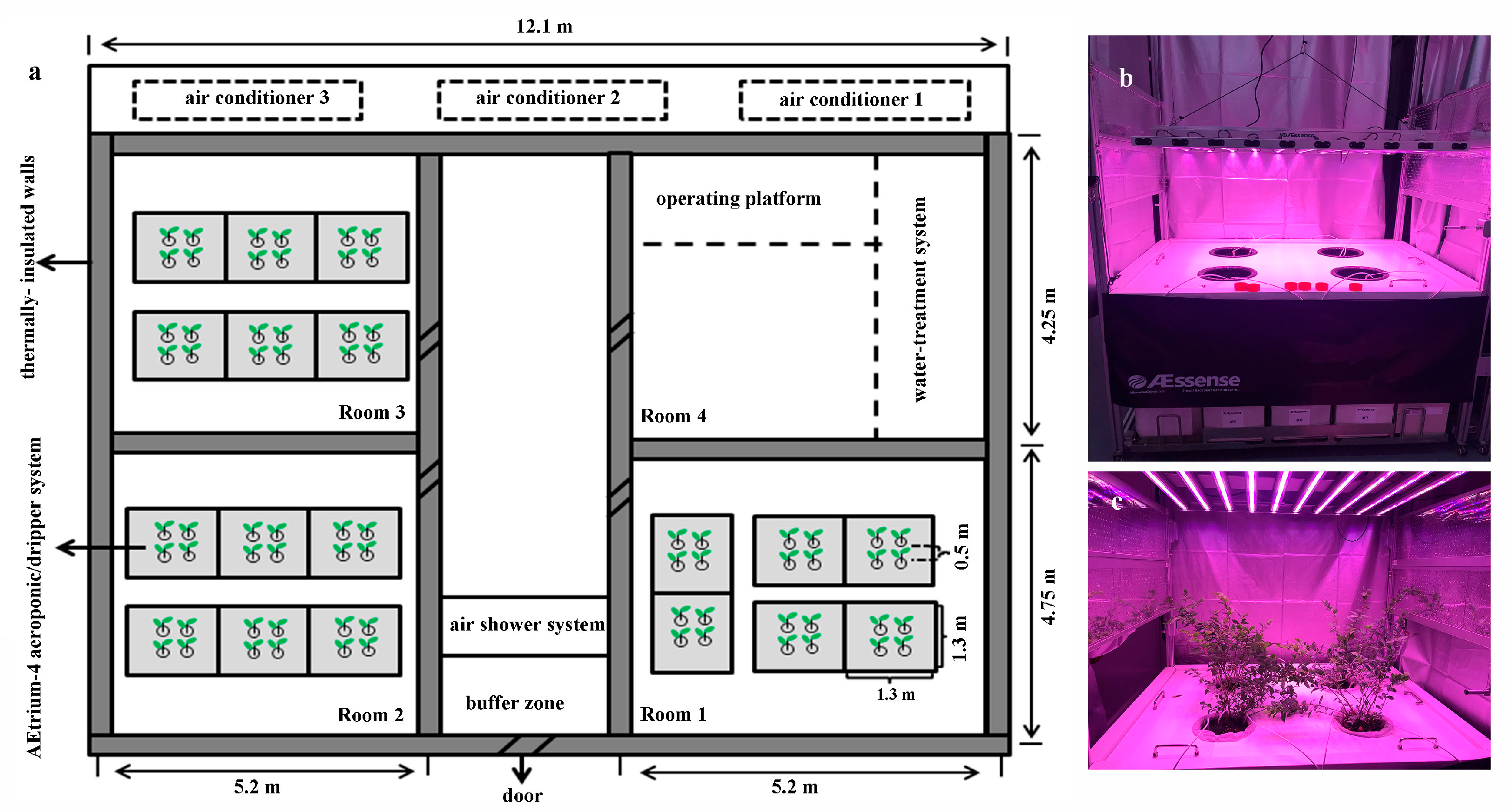
2.1. Plant Materials and Growth Conditions
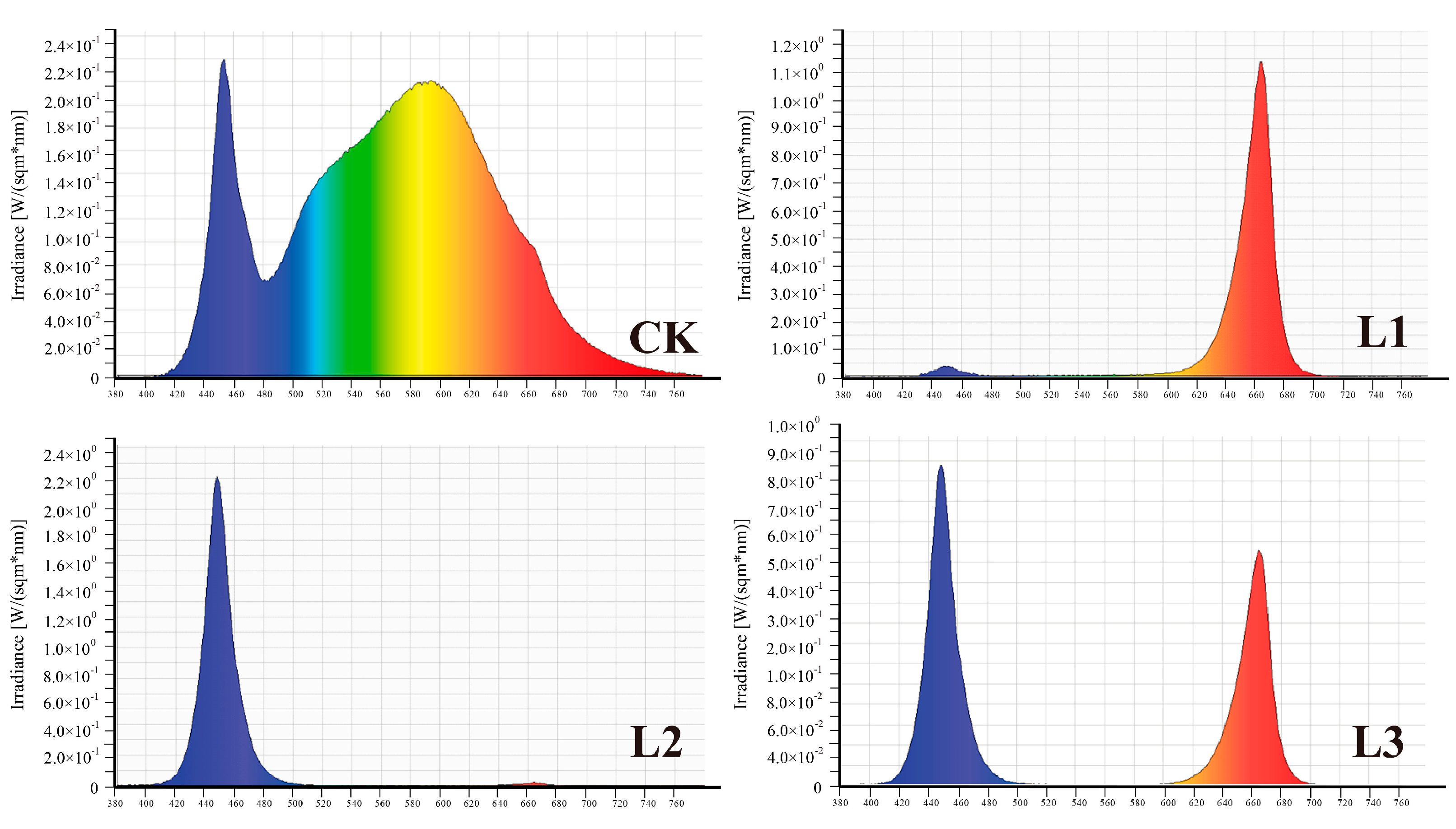
2.2. Experimental Designs
2.3. Observation of Phenotypic Traits
2.4. Analysis of Photosynthetic Characteristics
2.5. Determination of Nutrition Elements Uptake
2.6. Determination of Anthocyanin Content
2.7. Determination of Antioxidant Enzymes (CAT, POD, SOD), H2O2 Level, MDA, and Protein Content
2.8. Data Statistical Analysis
3. Results
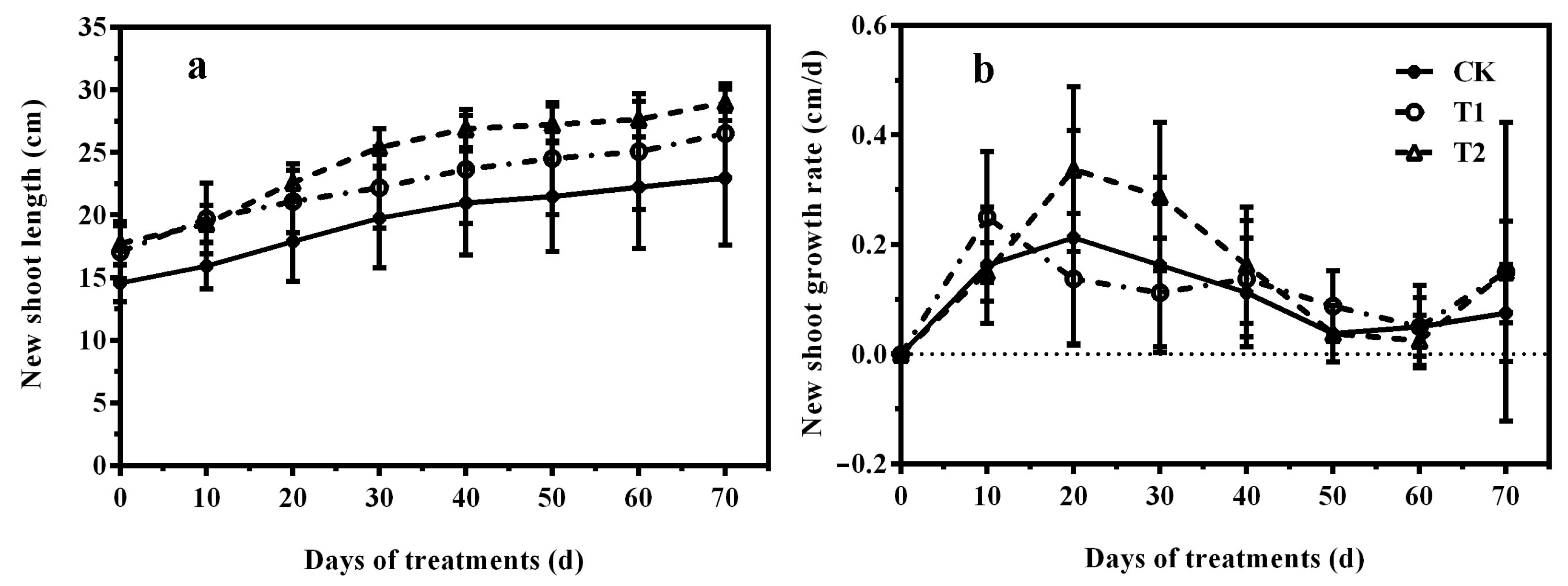
3.1. Effects of Nutrition on Plant Growth of ‘Misty’ in PFAL
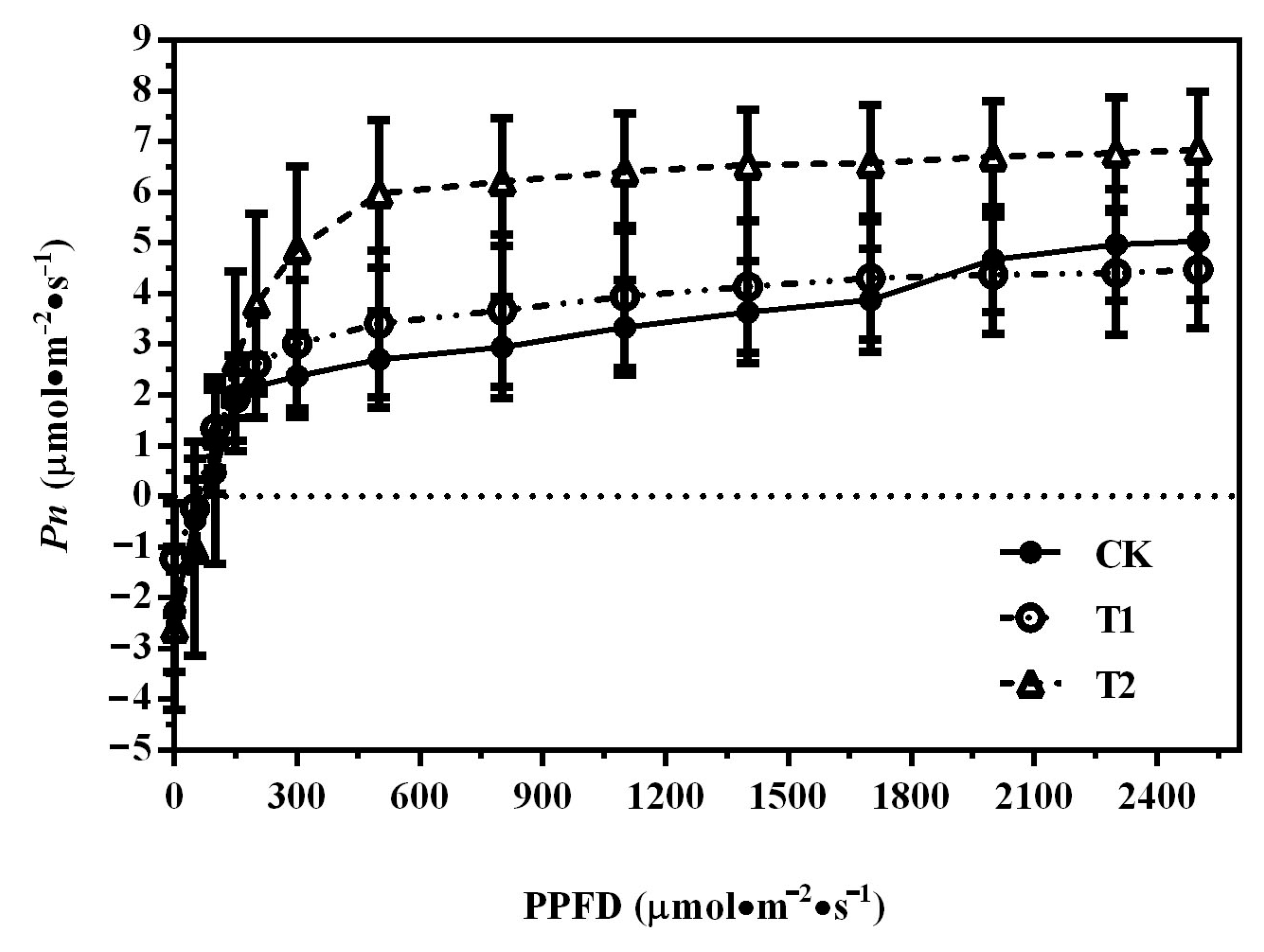
3.2. Photosynthetic Light-Response Curves of ‘Misty’ under Different Nutrition Treatments in PFAL
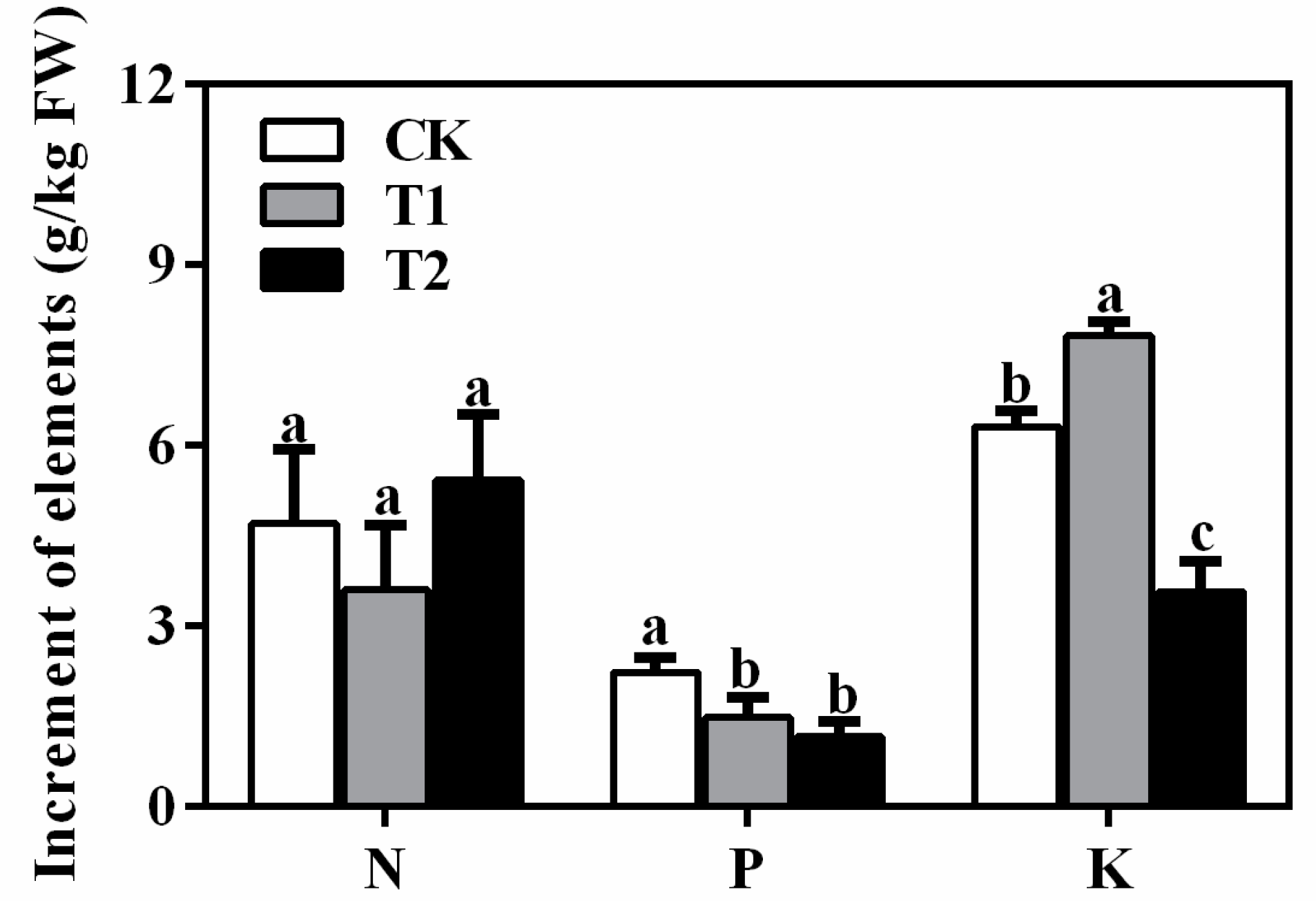
3.3. Effects of Different Nutrition Treatments on Elements Absorption of ‘Misty’ in PFAL
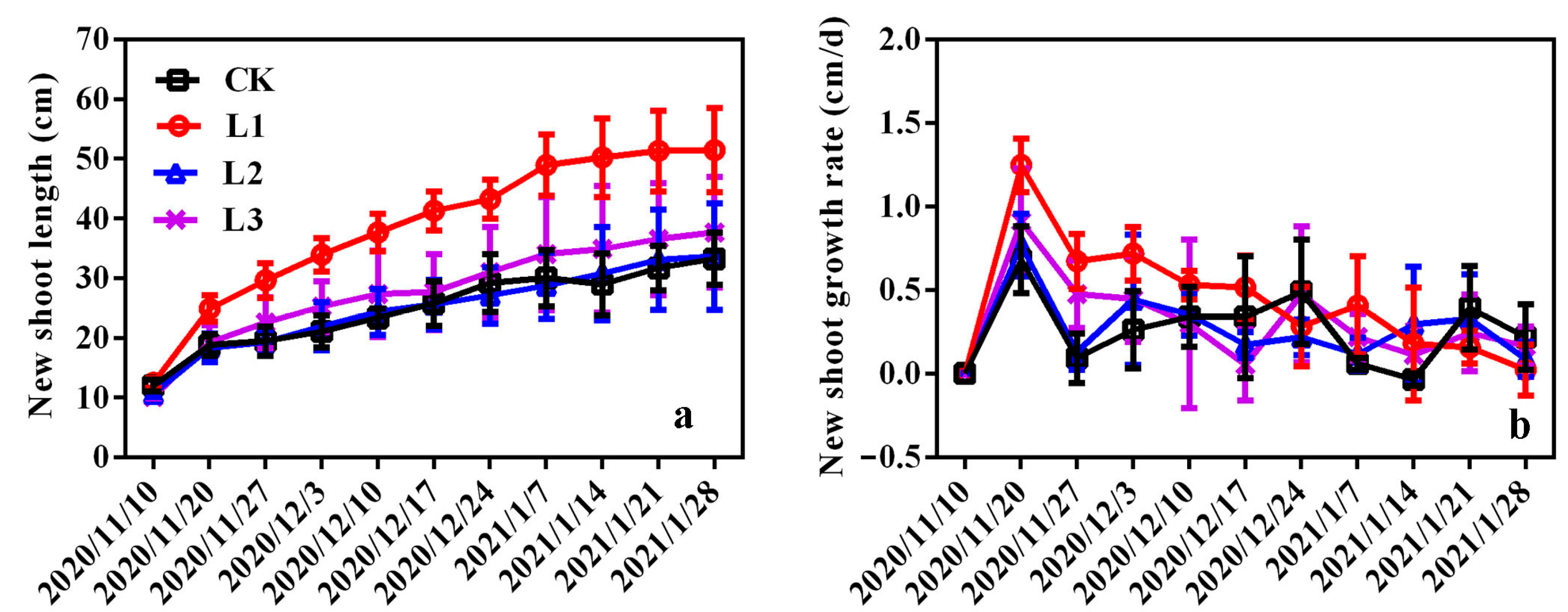
3.4. Effects of Different Light Qualities on Plant Growth of ‘Misty’ in PFAL
3.5. Effects of Different Light Qualities on Photosynthetic Characteristics of ‘Misty’ in PFAL
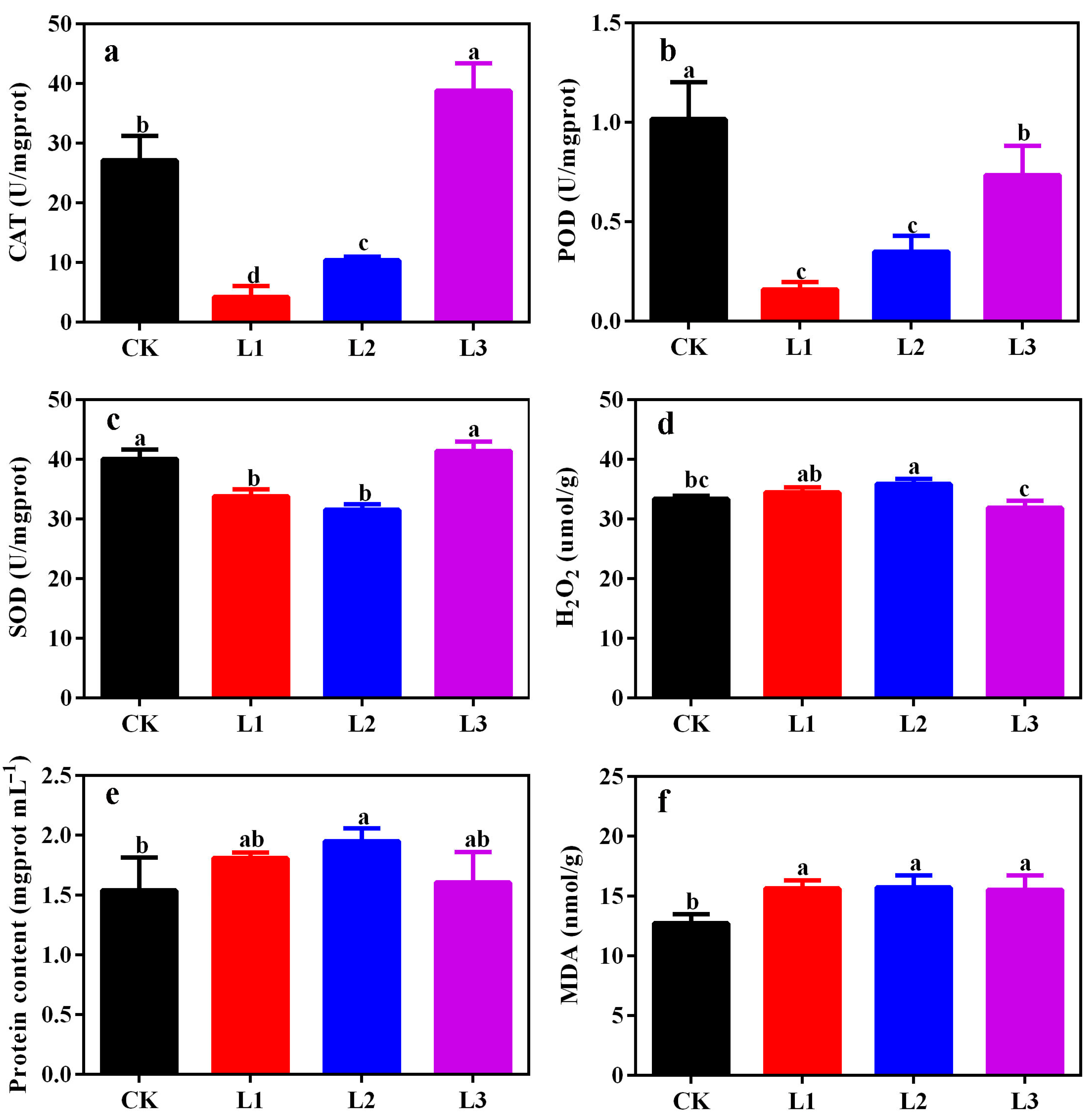
3.6. Changes in CAT, POD, SOD, H2O2, MDA, and Protein Content of ‘Misty’ Leaves in PFAL
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kozai, T. Resource use efficiency of closed plant production system with artificial light: Concept, estimation and application to plant factory. Proc. Jpn. Acad. Ser. B 2013, 89, 447–461. [Google Scholar] [CrossRef] [PubMed]
- Kozai, T. Plant factory in Japan—Current situation and perspectives. Chron. Hortic. 2013, 53, 8–11. [Google Scholar]
- Kozai, T. Towards sustainable plant factories with artificial lighting (PFALs) for achieving SDGs. Int. J. Agric. Biol. Eng. 2019, 12, 28–37. [Google Scholar] [CrossRef]
- Cho, H.Y.; Kadowaki, M.; Che, J.; Takahashi, S.; Horiuchi, N.; Ogiwara, I. Influence of light quality on flowering characteristics, potential for year-round fruit production and fruit quality of blueberry in a plant factory. Fruits 2019, 74, 3–10. [Google Scholar] [CrossRef]
- Kozai, T.; Niu, G. Plant factory as a resource—Efficient closed plant production system. Plant Fact. 2016, 2016, 69–90. [Google Scholar]
- Joshi, J.; Zhang, G.; Shen, S.; Supaibulwatana, K.; Watanabe, C.K.A.; Yamori, W. A combination of downward lighting and supplemental upward lighting improves plant growth in a closed plant factory with artificial lighting. HortScience 2017, 52, 831–835. [Google Scholar] [CrossRef]
- Niu, G.; Kozai, T.; Sabeh, N. Physical environmental factors and their properties. In Plant Factory: An Indoor Vertical Farming System for Efficient Quality Food Production, 1st ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 129–140. [Google Scholar]
- Kikuchi, Y.; Kanematsu, Y.; Yoshikawa, N.; Okubo, T.; Takagaki, M. Environmental and resource use analysis of plant factories with energy technology options: A case study in Japan. J. Clean. Prod. 2018, 186, 703–717. [Google Scholar] [CrossRef]
- Watanabe, H. Light-controlled plant cultivation system in Japan—Development of a vegetable factory using LEDs as a light source for plants. Acta Hortic. 2011, 907, 37–44. [Google Scholar] [CrossRef]
- Shimizu, H.; Saito, Y.; Nakashima, H.; Miyasaka, J.; Ohdoi, K. Light environment optimization for lettuce growth in plant factory. Int. Fed. Autom. Control. 2011, 44, 605–609. [Google Scholar] [CrossRef]
- Shimizu, H.; Nakashima, H.; Miyaska, J.; Ohdoi, K. Current state and research trend in light environment for plant factory. Sens. Technol. Biomater. Food Agric. 2013, 8881, 888104. [Google Scholar]
- Wang, Y.; Chu, Y.; Wan, Z.; Zhang, G.; Liu, L.; Yan, Z. Root architecture, growth and photon yield of cucumber seedlings as influenced by daily light integral at different stages in the closed transplant production system. Horticulturae 2021, 7, 328. [Google Scholar] [CrossRef]
- Ohishi-Yamazaki, M.; Watanabe, M.; Nakanishi, A.; Che, J.; Horiuchi, N.; Ogiwara, I. Shortening of the juvenile phase of the southern highbush blueberry (Vaccinium corymbosum L. interspecific hybrid) grown in controlled rooms under artificial light. Hortic. J. 2018, 87, 329–339. [Google Scholar] [CrossRef]
- Cho, H.Y.; Kadowaki, M.; Che, J.; Horiuchi, N.; Ogiwara, I. Plant morphological characteristics for year-round production and its fruit quality in response to different light wavelengths in blueberry. Acta Hortic. 2018, 1206, 201–209. [Google Scholar] [CrossRef]
- Gupta-Elera, G.; Garrett, A.; Martinez, A.; Kraus, R.D.; Robison, R.; O’Neill, K. A comparison of antioxidant properties in organic and conventional blueberries. J. Food Res. 2012, 1, 1–7. [Google Scholar] [CrossRef]
- An, H.; Zhang, J.; Xu, F.; Jiang, S.; Zhang, X.; 2020. Transcriptomic profiling and discovery of key genes involved in adventitious root formation from green cuttings of highbush blueberry (Vaccinium corymbosum L.). BMC Plant Biol. 2020, 20, 182. [Google Scholar] [CrossRef]
- Li, S.; Zhang, J.; Zhang, L.; Fang, X.; Luo, J.; An, H.; Zhang, X. Genome-wide identification and comprehensive analysis reveals potential roles of long non-coding RNAs in fruit development of southern highbush blueberry (Vaccinium corymbosum L.). Front. Plant Sci. 2022, 13, 1078085. [Google Scholar] [CrossRef]
- Li, Y.; Pei, J.; Chen, L.; Sun, H. China blueberry industry report 2020. J. Jilin Agric. Univ. 2021, 43, 1–8. (In Chinese) [Google Scholar]
- Valenzuela-Estrada, L.R.; Vera-Caraballo, V.; Ruth, L.E.; Eissenstat, D.M. Root anatomy, morphology and longevity among root orders in Vaccinium corymbosum (Ericaceae). Am. J. Bot. 2008, 95, 1506–1514. [Google Scholar] [CrossRef]
- An, H.; Meng, J.; Xu, F.; Jiang, S.; Wang, X.; Shi, C.; Zhou, B.; Luo, J.; Zhang, X. Rooting ability of hardwood cuttings in highbush blueberry (Vaccinium corymbosum L.) using different indole-butyric acid concentrations. HortScience 2019, 54, 194–199. [Google Scholar] [CrossRef]
- Pescie, M.A.; Fradkin, M.; Lavado, R.S.; Chiocchio, V.M. Endophytic fungi in blueberry cultivars, in three production areas of Argentina. Physiol. Mol. Plant P. 2021, 115, 101662. [Google Scholar] [CrossRef]
- Aung, T.; Muramatsu, Y.; Horiuchi, N.; Che, J.; Mochizuki, Y.; Ogiwara, I. Plant growth and fruit quality of blueberry in a controlled room under artificial light. J. Japan Soc. Hortic. Sci. 2014, 83, 273–281. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; An, H.; Zhang, X.; Zhou, B. Integrated transcriptome and metabolome analysis reveals anthocyanin biosynthesis mechanisms in blueberry (Vaccinium corymbosum L.) leaves under different light qualities. Front. Plant Sci. 2022, 13, 1073332. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tiyip, T.; Ding, J.; Zhang, D.; Liu, W.; Wang, F. Quantitative estimation of organic matter content in arid soil using vis-NIR spectroscopy preprocessed by fractional derivative. J. Spectrosc. 2017, 2017, 1375158. [Google Scholar] [CrossRef]
- Li, L.; Zhang, H.; Liu, Z.; Cui, X.; Zhang, T.; Li, Y.; Zhang, L. Comparative transcriptome sequencing and de novo analysis of Vaccinium corymbosum during fruit and color development. BMC Plant Biol. 2016, 16, 223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; An, H.; Zhang, X.; Xu, F.; Zhou, B. Transcriptomic analysis reveals potential gene regulatory networks under cold stress of loquat (Eriobotrya japonica Lindl.). Front. Plant Sci. 2022, 13, 944269. [Google Scholar] [CrossRef]
- Pennisi, G.; Pistillo, A.; Orsini, F.; Cellini, A.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Crepaldi, A.; Gianquinto, G.; Marcelis, L.F.M. Optimal light intensity for sustainable water and energy use in indoor cultivation of lettuce and basil under red and blue LEDs. Sci. Hortic. 2020, 272, 109508. [Google Scholar] [CrossRef]
- Wang, Y.; Chen. Y.F.; Wu. W.H. Potassium and phosphorus transport and signaling in plants. J. Integr. Plant Biol. 2021, 63, 34–52. [Google Scholar] [CrossRef]
- Cabrera, R.; Areces, A.J.; Diaz-Larrea, J.; Garcia, L.N.; Cruz-Avina, J.R. Influence of the macronutrients N, P and K on the agarophyte Alsidium triquetrum (S.G. Gmelin) trevisan, during experimental culture. Am. J. Plant Sci. 2021, 12, 573–585. [Google Scholar] [CrossRef]
- Doyle, J.W.; Nambeesan, S.U.; Malladi, A. Physiology of nitrogen and calcium nutrition in blueberry (Vaccinium sp.). Agronomy 2021, 211, 765. [Google Scholar] [CrossRef]
- Korcak, R.F. Nutrition of blueberry and other calcifuges. Hortic. Rev. 1988, 10, 183–227. [Google Scholar]
- Bryla, D.R.; Strik, B.C. Nutrient requirements, leaf tissue standards, and new options for fertigation of northern highbush blueberry. Horttechnology 2015, 25, 464–470. [Google Scholar] [CrossRef]
- Nestby, R.; Martinussen, I.; Krogstad, T.; Uleberg, E. Effects of fertilization, tiller cutting and environment on plant growth and yield of European blueberry (Vaccinium myrtillus L.) in Norwegian forest fields. J. Berry Res. 2014, 4, 79–95. [Google Scholar] [CrossRef]
- Wei, B.; Lou, X.; Wang, H.; Li, G. Effects of macroelements deficiency on the plant growth and gas exchange parameters of blueberries. Sci. Technol. Eng. 2021, 21, 1288–1294. (In Chinese) [Google Scholar]
- Li, Y.; Zhao, S.; Dai, H.; Guo, X. Effects of Nitrogen, phosphorus and potassium on growth, fruit production and leaf physiology in blueberry. Acta Hortic. 2009, 810, 759–764. [Google Scholar]
- Haynes, R.; Goh, K.M. Ammonium and nitrate nutrition of plants. Biol. Rev. 1978, 53, 465–510. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Tian, J.P.; Cui, J.; Hong, Y.H.; Luan, Y.S. Effects of different NH4+/NO3− ratios on the photosynthetic and physiology responses of blueberry (Vaccinium spp.) seedlings growth. J. Plant Nutr. 2021, 44, 854–864. [Google Scholar]
- Yanagi, T.; Okamoto, K.; Takita, S. Effects of blue, red, and blue/red lights of two different PPF levels on growth and morphogenesis of lettuce plants. Acta Hortic. 1996, 440, 117–122. [Google Scholar] [CrossRef]
- Dou, H.; Niu, G.; Gu, M.; Masabni, J.G. Effects of light quality on growth and phytonutrient accumulation of herbs under controlled environments. Horticulturae 2017, 3, 36. [Google Scholar] [CrossRef]
- Zhang, X.; He, D.; Niu, G.; Yan, Z.; Song, J. Effects of environment lighting on the growth, photosynthesis, and quality of hydroponic lettuce in a plant factory. Int. J. Agric. Biol. Eng. 2018, 2, 33–40. [Google Scholar] [CrossRef]
- Son, K.H.; Oh, M.M. Leaf shape, growth, and antioxidant phenolic compounds of two lettuce cultivars grown under various combinations of blue and red light-emitting diodes. HortScience 2013, 48, 988–995. [Google Scholar] [CrossRef]
- Di, Q.; Li, J.; Du, Y.; Wei, M.; Shi, Q.; Yang, F. Combination of red and blue lights improved the growth and development of eggplant (Solanum melongena L.) seedlings by regulating photosynthesis. J. Plant Growth Regul. 2021, 40, 1477–1492. [Google Scholar] [CrossRef]
- Kong, Y.; Nemali, K. Blue and far-red light affect area and number of individual leaves to influence vegetative growth and pigment synthesis in lettuce. Front. Plant Sci. 2021, 12, 667407. [Google Scholar] [CrossRef] [PubMed]
- Khoramtabrizi, M.; Aliniaeifard, S.; Chegini, G. Effects of different artificial light spectra on growth of lettuce in a continuous plant factory system. Acta Hortic. 2020, 1271, 101–106. [Google Scholar] [CrossRef]
- Tapingkae, T.; Taji, A. Light quality and quantity: Their effects on in vitro growth and development of two Australian plant species. Acta Hortic. 2000, 541, 281–288. [Google Scholar] [CrossRef]
- Ptushenko, V.V.; Avercheva, O.V.; Bassarskaya, E.M.; Berkovich, Y.A.; Erokhin, A.N.; Smolyanina, S.O.; Zhigalova, T.V. Possible reasons of a decline in growth of Chinese cabbage under a combined narrowband red and blue light in comparison with illumination by high-pressure sodium lamp. Sci. Hortic. 2015, 194, 267–277. [Google Scholar] [CrossRef]
- Hughes, N.M.; Lev-Yadun, S. Review: Why do some plants have leaves with red or purple undersides? Environ. Exp. Bot. 2023, 205, 105126. [Google Scholar] [CrossRef]
- Petridis, A.; van der Kaay, J.; Chrysanthou, E.; McCallum, S.; Graham, J.; Hancock, R.D. Photosynthetic limitation as a factor influencing yield in highbush blueberries (Vaccinium corymbosum) grown in a northern European environment. J. Exp. Bot. 2018, 69, 3069–3080. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Yu, D.J.; Kim, T.C.; Lee, H.J. Growth and photosynthetic characteristics of blueberry (Vaccinium corymbosum cv. Bluecrop) under various levels. Sci. Hortic. 2019, 129, 486–492. [Google Scholar] [CrossRef]
- Gechev, T.; Willekens, H.; Montagu, M.V.; Inze, D.; Van Camp, W.; Toneva, V.; Minkov, I. Different responses of tobacco antioxidant enzymes to light and chilling stress. J. Plant Physiol. 2003, 160, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Liu, W.; Zha, L.; Zhou, C.; Zhang, Y.; Li, B. Differential effects of high duration on growth, nutritional quality, and oxidative stress of hydroponic lettuce under red and blue LED irradiation. Sci. Hortic. 2020, 268, 109366. [Google Scholar] [CrossRef]
- Yan, Z.; Wang, L.; Wang, Y.; Chu, Y.; Lin, D.; Yang, Y. Morphological and physiological properties of greenhouse-grown cucumber seedlings as influenced by supplementary light-emitting diodes with same daily light integral. Horticulturae 2021, 7, 361. [Google Scholar] [CrossRef]
- Wang, S.; Verma, S.K.; Said, I.H.; Thomsen, L.; Ullrich, M.S.; Kuhnert, N. Changes in the fucoxanthin production and protein profiles in Cylindrotheca closterium in response to blue light-emitting diode light. Microb. Cell Fact. 2018, 17, 110. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wei, S.Q.; Liu, N.; Xu, L.J.; Yang, P. Growth of cucumber seedlings in different varieties as affected by light environment. Int. J. Agric. Biol. Eng. 2020, 13, 73–78. [Google Scholar]
- Wei, H.; Wang, M.Z.; Jeong, B.R. Effect of supplementary lighting duration on growth and activity of antioxidant enzymes in grafted watermelon seedlings. Agronomy 2020, 10, 337. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height (cm) | Stem Diameter (mm) | New Shoot Length (cm) | New Shoot Number | Chlorophyll Content (SPAD Value) |
|---|---|---|---|---|---|
| CK | 61.94 ± 16.45 a | 8.11 ± 1.89 b | 22.95 ± 5.35 b | 20.50 ± 6.63 b | 57.00 ± 5.91 a |
| T1 | 63.25 ± 9.82 a | 11.38 ± 2.42 a | 26.51 ± 5.3.56 ab | 18.75 ± 3.69 b | 52.90 ± 2.93 a |
| T2 | 72.50 ± 12.00 a | 11.95 ± 3.27 a | 29.00 ± 1.45 a | 29.79 ± 6.10 a | 56.93 ± 4.37 a |
| Treatments | Pn (µmol·m−2·s−1) | Gs (µmol·m−2·s−1) | Ci (µmol·m−2·s−1) | E (mmol·m−2·s−1) |
|---|---|---|---|---|
| CK | 5.54 ± 0.85 c | 52.45 ± 16.22 a | 271.68 ± 23.93 a | 1.59 ± 0.39 b |
| T1 | 5.95 ± 0.71 b | 51.38 ± 9.41 a | 247.04 ± 14.79 b | 1.61 ± 0.25 b |
| T2 | 6.38 ± 0.77 a | 62.34 ± 13.34 a | 228.38 ± 24.83 c | 1.81 ± 0.31 a |
| Plant Height | Stem Diameter | New Shoot Length | New Shoot Number | SPAD Value | Pn | Ci | gs | E | N | P | K | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant height | 1 | |||||||||||
| Stem diameter | 0.843 ** | 1 | ||||||||||
| New shoot length | 0.756 ** | 0.915 ** | 1 | |||||||||
| New shoot number | 0.891 ** | 0.837 ** | 0.841 ** | 1 | ||||||||
| SPAD value | 0.875 ** | 0.793 ** | 0.831 ** | 0.903 ** | 1 | |||||||
| Pn | 0.064 | 0.158 | 0.074 | −0.024 | −0.134 | 1 | ||||||
| Ci | −0.315 | −0.438 * | −0.484* | −0.448* | −0.287 | −0.451 * | 1 | |||||
| gs | 0.075 | 0.019 | −0.021 | 0.026 | 0.027 | 0.701 ** | −0.128 | 1 | ||||
| E | 0.068 | 0.020 | −0.033 | 0.047 | 0.029 | 0.682 ** | −0.191 | 0.951 ** | 1 | |||
| N | 0.962 ** | 0.866 ** | 0.773 ** | 0.845 ** | 0.814 ** | 0.131 | −0.369 | 0.064 | 0.040 | 1 | ||
| P | 0.846 ** | 0.588 ** | 0.590 ** | 0.756 ** | 0.895 ** | −0.107 | −0.134 | 0.107 | 0.122 | 0.760 ** | 1 | |
| K | 0.894 ** | 0.804 ** | 0.756 ** | 0.768 ** | 0.866 ** | 0.015 | −0.283 | 0.013 | 0.051 | 0.893 ** | 0.847 ** | 1 |
| Treatments | Plant Height (cm) | Stem Diameter (mm) | New Shoot Length (cm) | New Shoot Number | Leaf Area (mm2) | Chlorophyll Content (SPAD Value) | Anthocyanin Content (mg/kg FW) |
|---|---|---|---|---|---|---|---|
| CK | 61.33 ± 20.66 a | 11.66 ± 1.05 a | 33.22 ± 4.38 b | 21.25 ± 5.32 a | 1349.39 ± 50.07 a | 53.53 ± 3.81 c | 28.80 ± 0.96 d |
| L1 | 81.25 ± 12.62 a | 12.30 ± 1.01 a | 51.43 ± 7.03 a | 17.50 ± 3.11 a | 1608.08 ± 261.03 a | 50.35 ± 2.44 c | 47.43 ± 1.35 c |
| L2 | 69.80 ± 11.00 a | 11.93 ± 1.65 a | 33.64 ± 8.94 b | 15.25 ± 1.50 a | 1427.97 ± 158.71 a | 64.68 ± 1.23 a | 132.45 ± 6.40 a |
| L3 | 68.63 ± 17.65 a | 11.69 ± 0.66 a | 37.68 ± 9.32 b | 22.67 ± 9.24 a | 1320.08 ± 228.47 a | 58.89 ± 2.68 b | 106.07 ± 2.64 b |
| Tre atments | Pn (µmol·m−2·s−1) | Gs (µmol·m−2·s−1) | Ci (µmol·m−2·s−1) | E (mmol·m−2·s−1) |
|---|---|---|---|---|
| CK | 1.33 ± 0.27 a | 38.87 ± 6.17 a | 311.38 ± 8.67 c | 0.98 ± 0.10 b |
| L1 | 0.68 ± 0.25 b | 39.25 ± 3.20 a | 339.38 ± 13.81 b | 1.23 ± 0.14 a |
| L2 | 0.42 ± 0.25 c | 42.63 ± 6.30 a | 354.13 ± 15.00 a | 1.17 ± 0.11 a |
| L3 | 1.35 ± 0.19 a | 44.50 ± 7.17 a | 316.75 ± 14.73 c | 1.22 ± 0.14 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, H.; Zhang, J.; Zhang, L.; Li, S.; Zhou, B.; Zhang, X. Effects of Nutrition and Light Quality on the Growth of Southern Highbush Blueberry (Vaccinium corymbosum L.) in an Advanced Plant Factory with Artificial Lighting (PFAL). Horticulturae 2023, 9, 287. https://doi.org/10.3390/horticulturae9020287
An H, Zhang J, Zhang L, Li S, Zhou B, Zhang X. Effects of Nutrition and Light Quality on the Growth of Southern Highbush Blueberry (Vaccinium corymbosum L.) in an Advanced Plant Factory with Artificial Lighting (PFAL). Horticulturae. 2023; 9(2):287. https://doi.org/10.3390/horticulturae9020287
Chicago/Turabian StyleAn, Haishan, Jiaying Zhang, Liqing Zhang, Shuigen Li, Boqiang Zhou, and Xueying Zhang. 2023. "Effects of Nutrition and Light Quality on the Growth of Southern Highbush Blueberry (Vaccinium corymbosum L.) in an Advanced Plant Factory with Artificial Lighting (PFAL)" Horticulturae 9, no. 2: 287. https://doi.org/10.3390/horticulturae9020287
APA StyleAn, H., Zhang, J., Zhang, L., Li, S., Zhou, B., & Zhang, X. (2023). Effects of Nutrition and Light Quality on the Growth of Southern Highbush Blueberry (Vaccinium corymbosum L.) in an Advanced Plant Factory with Artificial Lighting (PFAL). Horticulturae, 9(2), 287. https://doi.org/10.3390/horticulturae9020287