
NPK Fertilization of Serianthes Plants Influences Growth and Stoichiometry of Leaf Nutrients


Abstract
:
1. Introduction
2. Materials and Methods
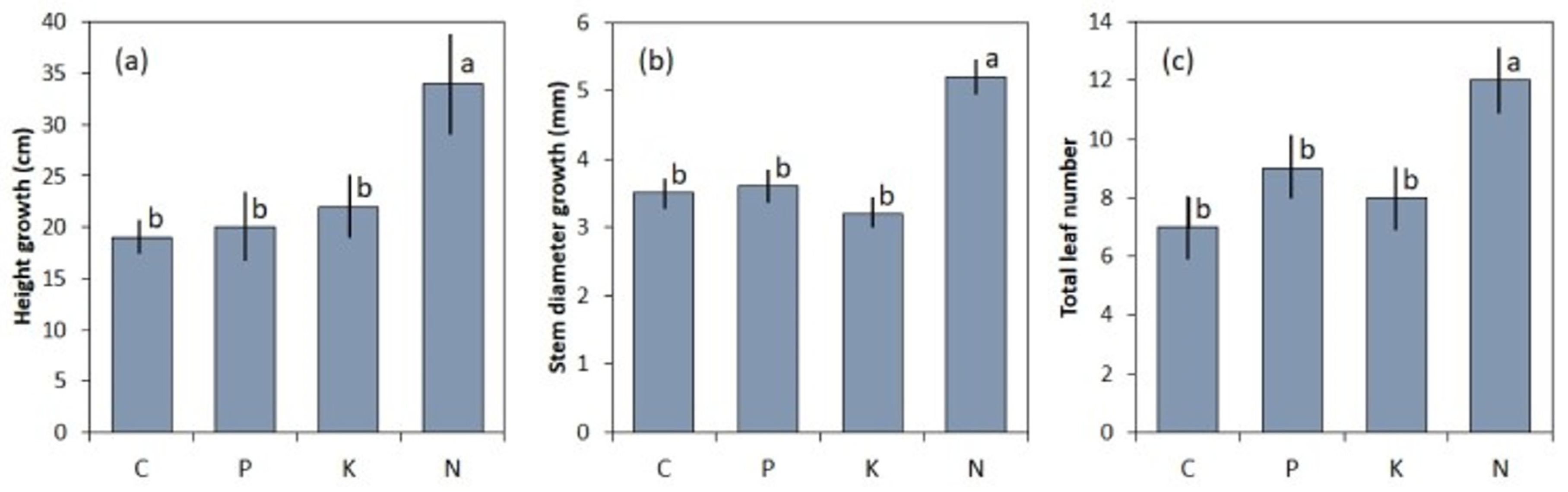
2.1. Serianthes kanehirae
2.2. Serianthes grandiflora and Serianthes nelsonii
2.3. Statistical Analysis
2.4. Soil Nutrients
3. Results
3.1. Serianthes kanehirae
3.2. Serianthes grandiflora and Serianthes nelsonii
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marler, T.E. Asexual reproduction to propel recovery efforts of the critically endangered Håyun Lågu tree (Serianthes nelsonii Merr.). Trop. Conserv. Sci. 2017, 10, 1940082917697707. [Google Scholar] [CrossRef] [Green Version]
- Marler, T.E. Diel root extension patterns of three Serianthes species are modulated by plant size. Plant Signal. Behav. 2017, 12, e1327496. [Google Scholar] [CrossRef] [Green Version]
- Marler, T.E. Temperature and imbibition influence Serianthes seed germination. Plants 2019, 8, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marler, T.E.; Musser, C.; Cascasan, A.N.J.; Cruz, G.N.; Deloso, B.E. Adaptive management lessons for Serianthes nelsonii conservation. Horticulturae 2021, 7, 43. [Google Scholar] [CrossRef]
- Wiles, G.; Williams, E. Serianthes nelsonii. IUCN Red List Threat. Species 2017. [Google Scholar] [CrossRef]
- United States Fish and Wildlife Service. Determination of endangered status for Serianthes nelsonii Merr. (Hayun lagu or Tronkon Guafi). Fed. Regist. 1987, 52, 4907–4910. [Google Scholar]
- Marler, T.E. Repetitive pruning of Serianthes nursery plants improves transplant quality and post-transplant survival. Plant Signal. Behav. 2019, 14, e1621246. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E.; Callaway, R.M. Talking with strangers: Improving Serianthes transplant quality with interspecific companions. Forests 2021, 12, 1192. [Google Scholar] [CrossRef]
- Jiang, J.; Wang, Y.P.; Yang, Y.; Yu, M.; Wang, C.; Yan, J. Interactive effects of nitrogen and phosphorus additions on plant growth vary with ecosystem type. Plant Soil 2019, 440, 523–537. [Google Scholar] [CrossRef]
- Jaquetti, R.K.; Gonçalves, J.F.d.C.; Ferraz, J.B.S.; Ferreira, M.J.; dos Santos Junior, U.M. Ecofunctional Traits and Biomass Production in Leguminous Tree Species under Fertilization Treatments during Forest Restoration in Amazonia. Forests 2016, 7, 76. [Google Scholar] [CrossRef] [Green Version]
- Rose, K.M.E.; Baribault, T.W.; Jacobs, D.F. Alternative field fertilization techniques to promote restoration of leguminous Acacia koa on contrasting tropical sites. For. Ecol. Manag. 2016, 376, 126–134. [Google Scholar] [CrossRef]
- Gerber, K.; Ross-Davis, A.; Perakis, S.S.; Davis, A.S. Early growth and ecophysiological responses of Koa (Acacia koa A. Gray) seedlings to reduced water and phosphorus. New For. 2022, 53, 643–660. [Google Scholar] [CrossRef]
- Jarrell, W.M.; Beverly, R.B. The dilution effect in plant nutrition studies. Adv. Agron. 1981, 34, 197–224. [Google Scholar]
- Chen, R.; Zhu, Y.; Cao, W.; Tang, L. A bibliometric analysis of research on plant critical dilution curve conducted between 1985 and 2019. Eur. J. Agron. 2021, 123, 126199. [Google Scholar] [CrossRef]
- Marler, T.E. Leaf elemental concentrations, stoichiometry, and resorption in Guam’s coastal karst forests. Diversity 2021, 13, 545. [Google Scholar] [CrossRef]
- Marler, T.E. Foliar nutrition of Serianthes nelsonii seedlings as a conservation tool. HortScience 2022, 57, 389–390. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Farrington, H. Nutrient limitation and soil development: Experimental test of a biogeochemical theory. Biogeochemistry. 1997, 37, 63–75. [Google Scholar] [CrossRef]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; U. S. Dept. of Agric.: Washington, DC, USA, 1954; p. 19. [Google Scholar]
- Santiago, L.S.; Wright, S.J.; Harms, K.E.; Yavitt, J.B.; Korine, C.; Garcia, M.N.; Turner, B.L. Tropical tree seedling growth responses to nitrogen, phosphorus and potassium addition. J. Ecol. 2012, 100, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.J. Plant responses to nutrient addition experiments conducted in tropical forests. Ecol. Monogr. 2019, 89, e01382. [Google Scholar] [CrossRef]
- Fataftah, N.; Edlund, E.; Lihavainen, J.; Bag, P.; Björkén, L.; Näsholm, T.; Jansson, S. Nitrate fertilization may delay autumn leaf senescence, while amino acid treatments do not. Physiol. Plant. 2022, 174, e13690. [Google Scholar] [CrossRef] [PubMed]
- National Fish and Wildlife Foundation. Guam Habitat Conservation Initiative. Available online: https://www.nfwf.org/programs/guam-habitat-conservation-initiative (accessed on 31 July 2022).
- Marler, T.E. Perennial trees associating with nitrogen-fixing symbionts differ in leaf after-life nitrogen and carbon release. Nitrogen 2020, 1, 111–124. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | C | P | K | N | p |
|---|---|---|---|---|---|
| Calcium 2 | 12.6 ± 0.6a 1 | 11.1 ± 0.9ab | 9.5 ± 0.6b | 8.1 ± 0.6c | <0.001 |
| Carbon 2 | 415 ± 4 | 414 ± 5 | 413 ± 4 | 421 ± 4 | 0.091 |
| Magnesium 2 | 2.7 ± 0.1b | 3.1 ± 0.2a | 2.4 ± 0.2bc | 2.2 ± 0.2c | 0.011 |
| Nitrogen 2 | 17.2 ± 0.5c | 21.2 ± 0.6b | 19.4 ± 0.6bc | 26.6 ± 0.7a | <0.001 |
| Phosphorus 2 | 1.9 ± 0.1b | 2.4 ± 0.2a | 1.6 ± 0.1b | 1.0 ± 0.1c | <0.001 |
| Potassium 2 | 10.4 ± 0.4b | 10.2 ± 0.3b | 15.3 ± 0.4a | 8.7 ± 0.2c | <0.001 |
| Boron 3 | 36.7 ± 1.1a | 38.0 ± 2.1a | 29.7 ± 1.1b | 24.9 ± 1.1c | <0.001 |
| Copper 3 | 1.7 ± 0.1b | 2.3 ± 0.2a | 1.4 ± 0.1c | 1.6 ± 0.2bc | 0.008 |
| Iron 3 | 61.3 ± 3.9a | 59.3 ± 3.2a | 45.3 ± 2.9b | 36.8 ± 1.9c | <0.001 |
| Manganese 3 | 51.9 ± 2.5a | 37.8 ± 2.4b | 39.8 ± 2.1b | 30.8 ± 1.4c | <0.001 |
| Zinc 3 | 33.2 ± 1.5a | 26.4 ± 1.1b | 22.1 ± 1.1c | 36.2 ± 1.9a | 0.008 |
| Variable | C | P | K | N | p |
|---|---|---|---|---|---|
| N:P | 9.3 ± 0.4c 1 | 9.0 ± 0.4d | 12.6 ± 1.4b | 27.0 ± 1.9a | <0.001 |
| N:K | 1.7 ± 0.1c | 2.1 ± 0.1b | 1.3 ± 0.1d | 3.1 ± 0.2a | <0.001 |
| K:P | 5.6 ± 0.3b | 4.3 ± 0.2c | 9.9 ± 0.8a | 8.8 ± 0.6a | <0.001 |
| Nutrient | C | P | K | N | p |
|---|---|---|---|---|---|
| S. nelsonii | |||||
| Nitrogen | 13.2 ± 0.3d 1 | 17.7 ± 0.5b | 15.1 ± 0.5c | 23.1 ± 0.6a | <0.001 |
| Phosphorus | 2.0 ± 0.1b | 2.6 ± 0.1a | 1.8 ± 0.1b | 1.2 ± 0.1c | <0.001 |
| S. grandiflora | |||||
| Nitrogen | 16.8 ± 0.4c | 19.9 ± 0.6b | 18.8 ± 0.5b | 27.5 ± 0.6a | <0.001 |
| Phosphorus | 1.5 ± 0.1b | 2.4 ± 0.1a | 1.5 ± 0.1b | 1.1 ± 0.1c | <0.001 |
| Nutrient | C | P | K | N | p |
|---|---|---|---|---|---|
| Calcium 2 | 13.0 ± 6a 1 | 12.5 ± 0.6ab | 11.2 ± 0.6b | 8.7 ± 0.4c | <0.001 |
| Carbon 2 | 419 ± 2b | 418 ± 3b | 417 ± 2b | 425 ± 3a | <0.001 |
| Magnesium 2 | 2.8 ± 0.1ab | 3.1 ± 0.2a | 2.6 ± 0.2b | 2.1 ± 0.2c | <0.001 |
| Potassium 2 | 11.0 ± 0.4b | 11.2 ± 0.4b | 15.5 ± 0.4a | 8.8 ± 0.2c | <0.001 |
| Boron 3 | 36.8 ± 1.9a | 38.2 ± 2.2a | 32.1 ± 1.7b | 29.5 ± 1.7c | <0.001 |
| Copper 3 | 1.9 ± 0.2ab | 2.3 ± 0.2a | 1.8 ± 0.2b | 1.9 ± 0.2ab | <0.001 |
| Iron 3 | 63.5 ± 3.5a | 60.9 ± 2.3a | 47.6 ± 2.6b | 39.8 ± 1.6c | <0.001 |
| Manganese 3 | 51.9 ± 2.2a | 38.0 ± 1.9b | 42.2 ± 1.3b | 30.5 ± 1.2c | <0.001 |
| Zinc 3 | 34.1 ± 2.1a | 30.0 ± 1.8b | 27.3 ± 2.1b | 37.8 ± 2.8a | <0.001 |
| Variable | C | P | K | N | p |
|---|---|---|---|---|---|
| N:P | 9.0 ± 0.8b 1 | 7.6 ± 0.6c | 10.8 ± 1.1b | 23.6 ± 1.4a | <0.001 |
| N:K | 1.4 ± 0.1c | 1.7 ± 0.1b | 1.1 ± 0.1d | 2.9 ± 0.2a | <0.001 |
| K:P | 6.3 ± 0.3c | 4.5 ± 0.2d | 9.8 ± 0.5a | 8.1 ± 0.2b | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marler, T.E. NPK Fertilization of Serianthes Plants Influences Growth and Stoichiometry of Leaf Nutrients. Horticulturae 2022, 8, 717. https://doi.org/10.3390/horticulturae8080717
Marler TE. NPK Fertilization of Serianthes Plants Influences Growth and Stoichiometry of Leaf Nutrients. Horticulturae. 2022; 8(8):717. https://doi.org/10.3390/horticulturae8080717
Chicago/Turabian StyleMarler, Thomas E. 2022. "NPK Fertilization of Serianthes Plants Influences Growth and Stoichiometry of Leaf Nutrients" Horticulturae 8, no. 8: 717. https://doi.org/10.3390/horticulturae8080717
APA StyleMarler, T. E. (2022). NPK Fertilization of Serianthes Plants Influences Growth and Stoichiometry of Leaf Nutrients. Horticulturae, 8(8), 717. https://doi.org/10.3390/horticulturae8080717