
Brassinolide Maximized the Fruit and Oil Yield, Induced the Secondary Metabolites, and Stimulated Linoleic Acid Synthesis of Opuntia ficus-indica Oil

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Location, Plant Source, and Layout
2.2. Preparation and Brassinolide Application
2.3. Cladodes Parameters and Harvesting Date
2.4. Fruit Characteristics
2.5. Fruit, Seed, and Oil Yields
2.6. GC–MS Analysis of the Seeds Oil
2.7. Chlorophyll Pigments Determination
2.8. Protein and Vitamin C Determination
2.9. Polyphenols Estimation
2.10. Antioxidant Activity (IC50) Determinations
2.11. Minerals Estimation
2.12. Statistical Analysis
3. Results
3.1. Cladodes Characteristics and Harvesting Date
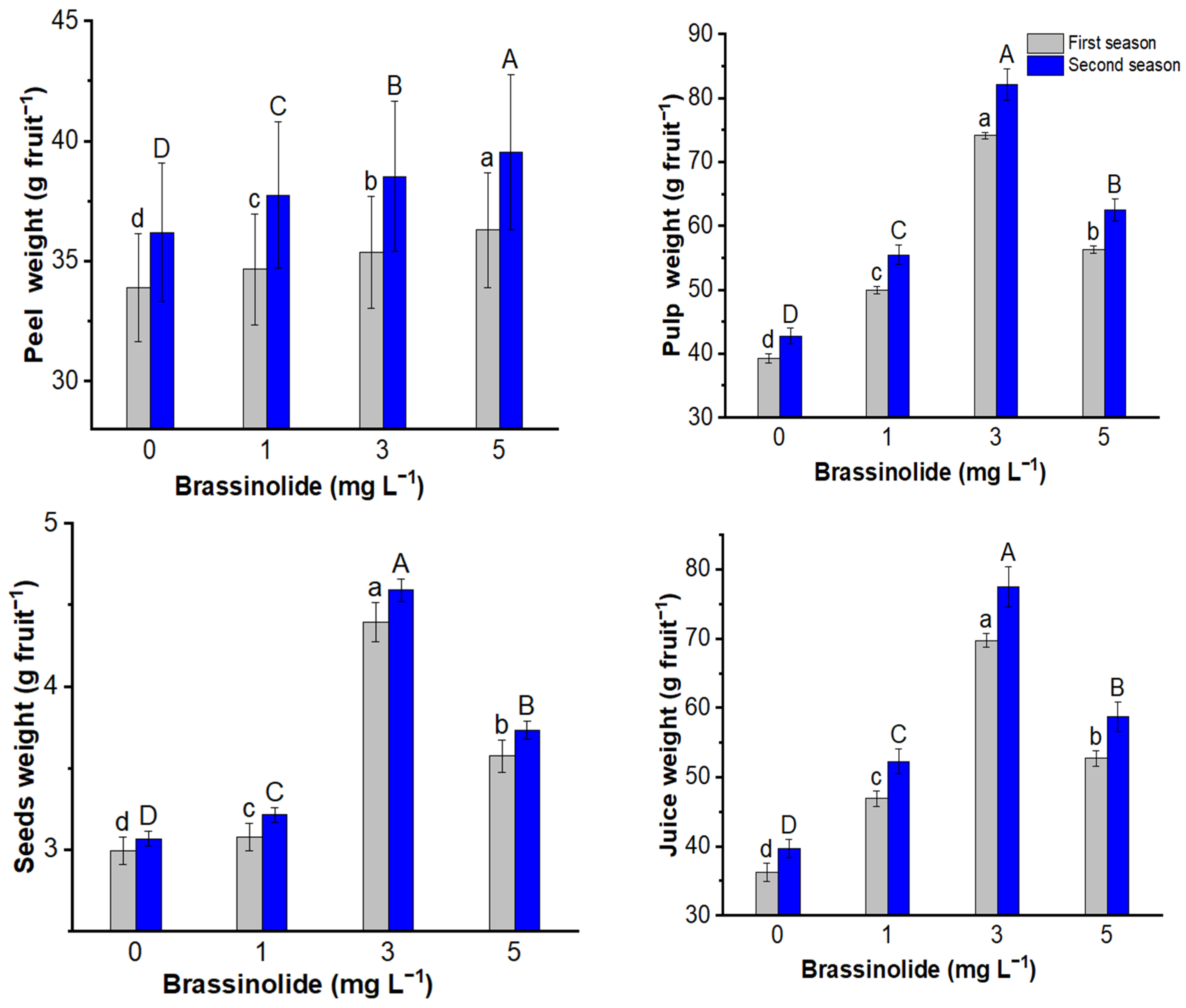
3.2. Fruit Characteristics
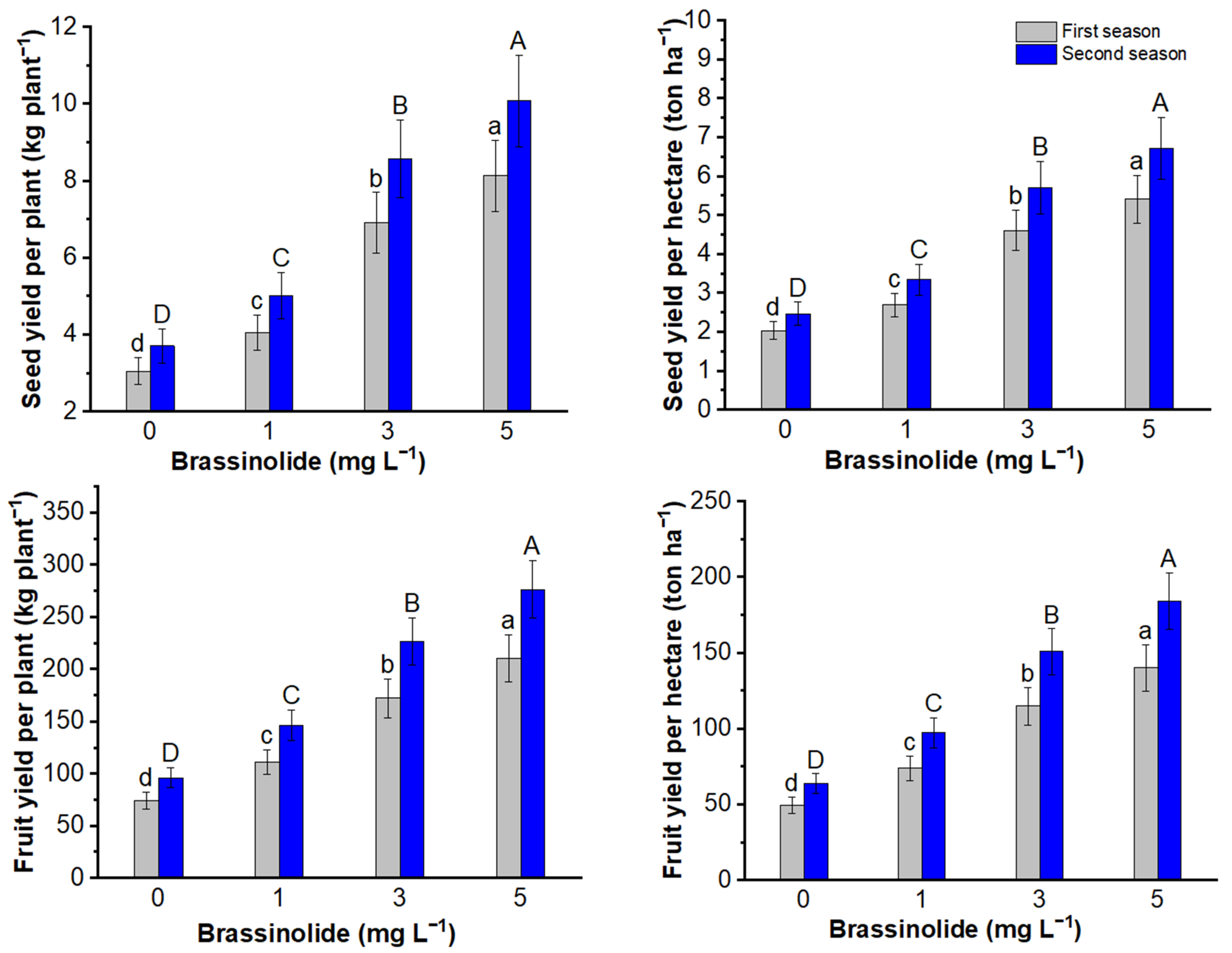
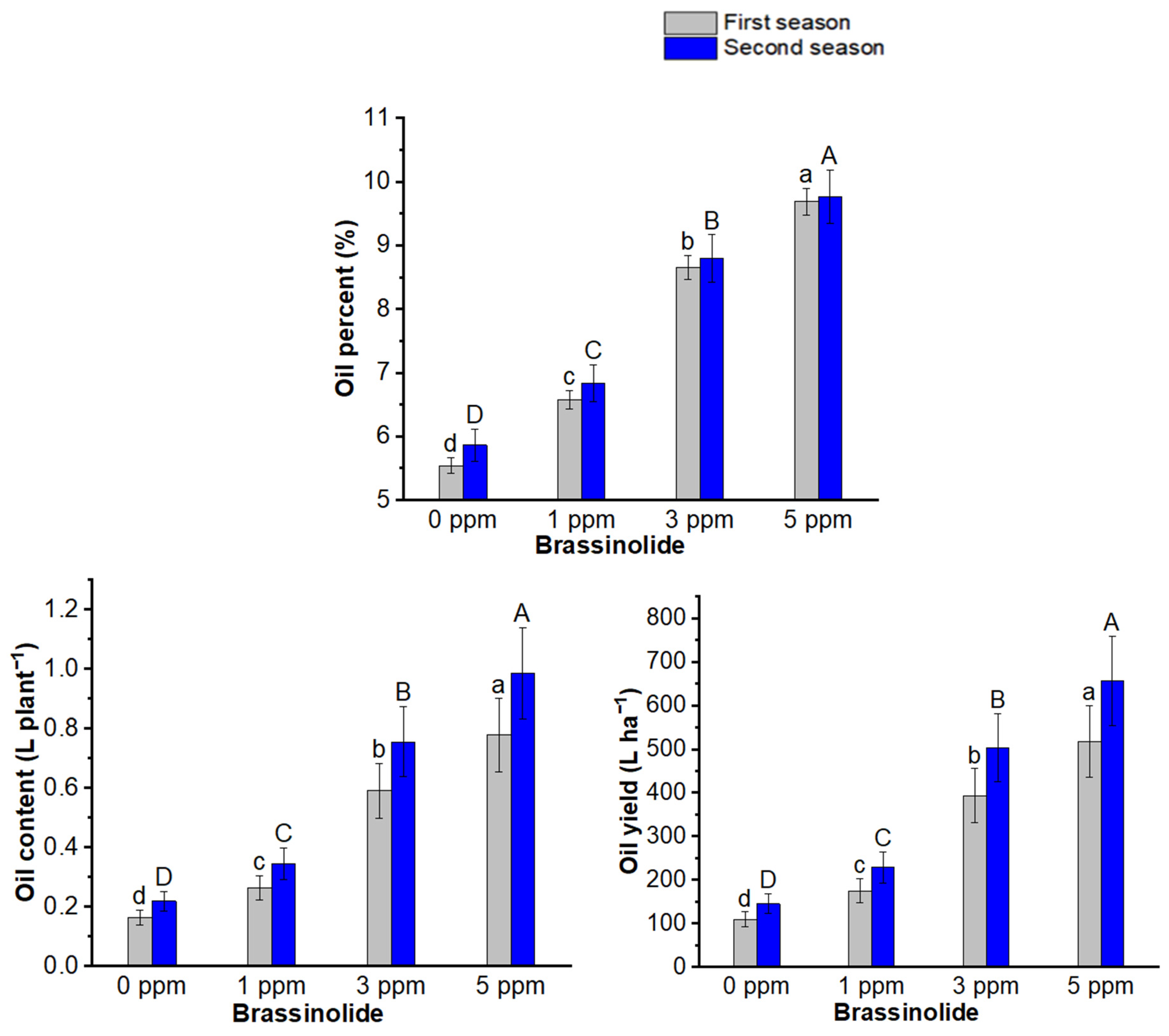
3.3. Seed, Fruit, and Oil Yields
3.4. GC–MS Analysis of the Seeds Oil

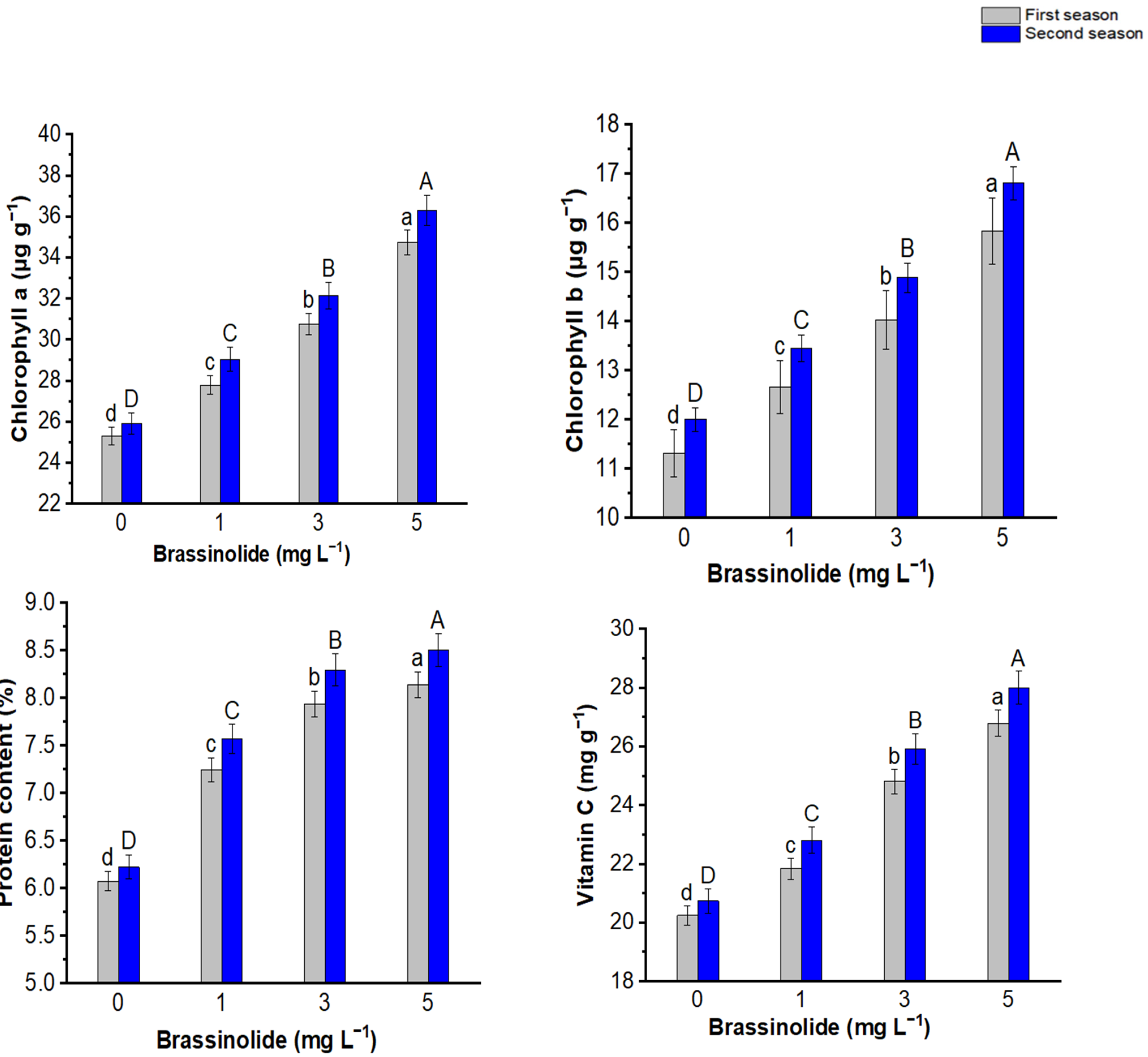
3.5. Chlorophyll Pigments
3.6. Protein and Vitamin C Content

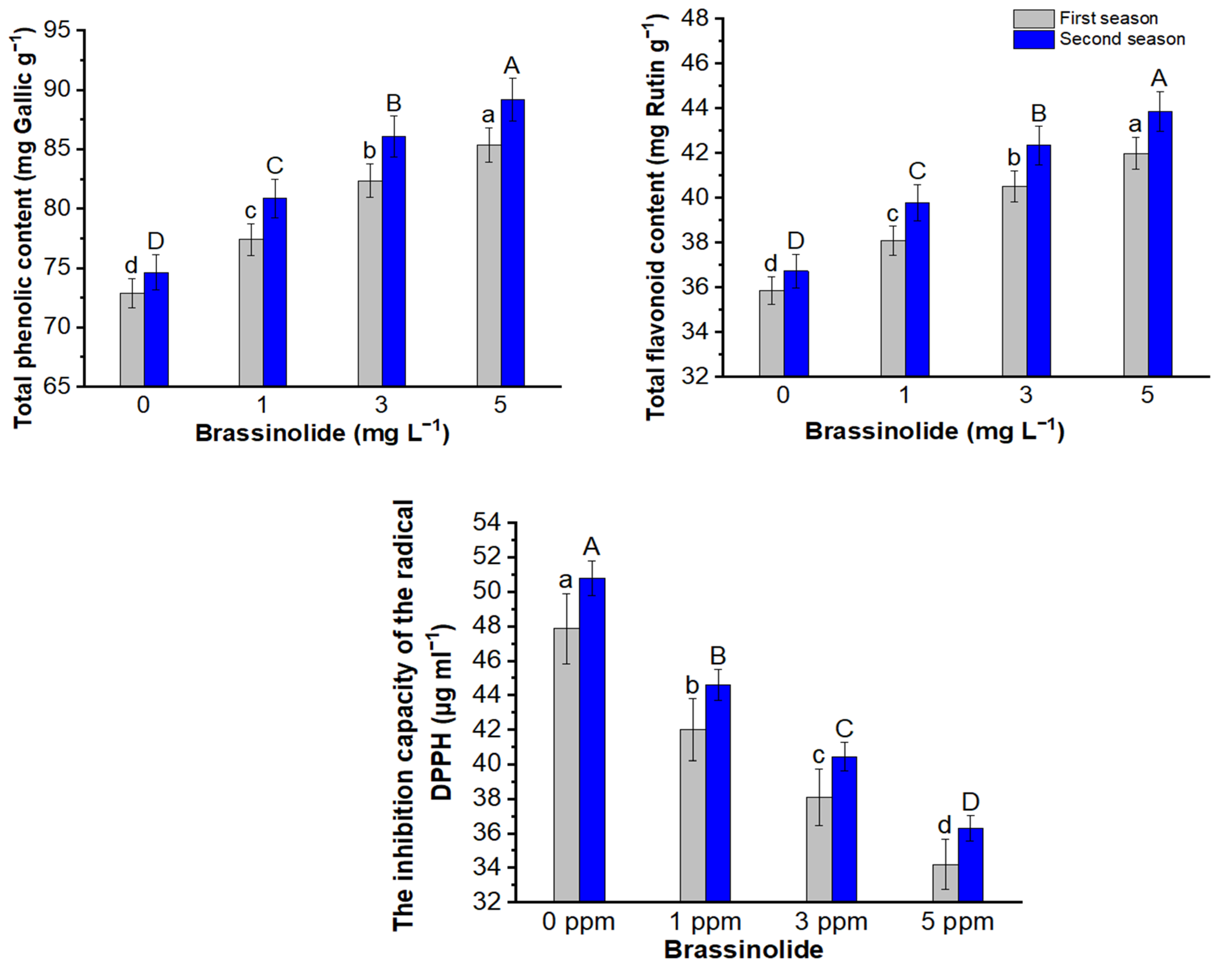
3.7. Phenolic, Flavonoid, and Antioxidant Activity (IC50)
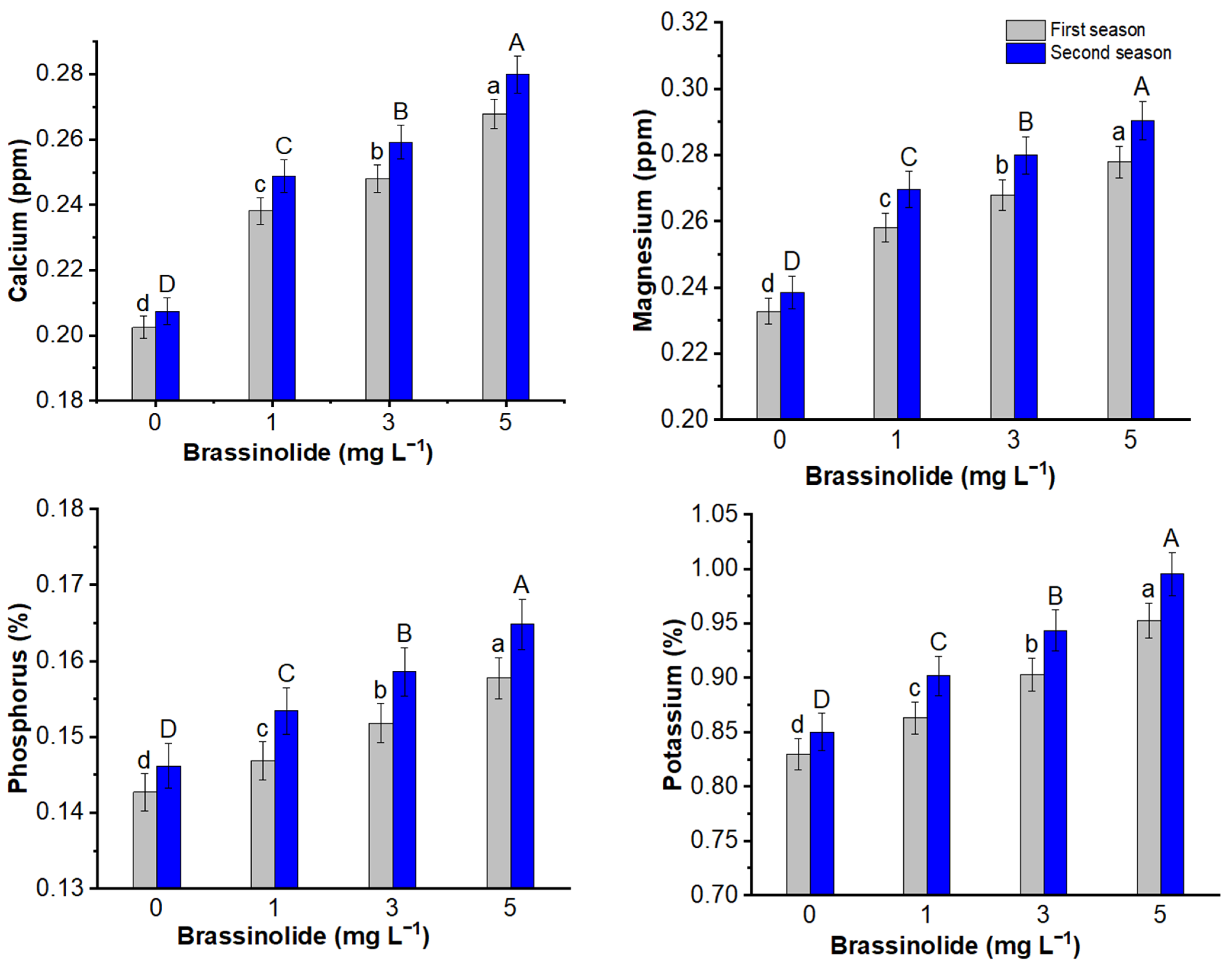
3.8. Minerals Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mobhammer, M.R.; Stintzing, F.C.; Carle, R. Cactus pear fruits (Opuntia spp.): A review of processing technologies and current uses. J. Prof. Assoc. Cactus Dev. 2006, 8, 1–24. [Google Scholar]
- Snyman, H.A.; Fouche, H.J.; Avenant, P.L.; Ratsele, C. Frost sensitivity of Opuntia ficus−indica and O. robusta in a semiarid climate of South Africa. J. Prof. Ass. Cactus Dev. 2007, 9, 1–21. [Google Scholar]
- Rabhi, A.; Falleh, H.; Limam, F.; Ksouri, R.; Abdelly, C.; Raies, A. Upshot of the ripening time on biological activities, phenol content and fatty acid composition of Tunisian Opuntia ficus-indica fruit. Afr. J. Biotechnol. 2013, 12, 5875–5885. [Google Scholar]
- Bhatt, M.R.; Nagar, P.S. Evaluation of physicochemical property and fatty acid composition of Opuntia elatior seed oil. J. Prof. Assoc. Cactus Dev. 2013, 15, 13–19. [Google Scholar]
- Chahdoura, H.; Barreira, J.C.; Barros, L.; Santos-Buelga, C.; Ferreira, I.C.; Isabel, C.F.; Achour, L. Seeds of opuntia spp. as a novel high potential by-product: Phytochemical characterization and antioxidant activity. Ind. Crops Prod. 2015, 65, 383–389. [Google Scholar] [CrossRef]
- Ramadan, M.F.; Mörsel, J.-T. Oil cactus pear (Opuntia ficus-indica L.). Food Chem. 2003, 82, 339–345. [Google Scholar] [CrossRef]
- Jun, H.I.; Cha, M.N.; Yang, E.I.; Choi, D.G.; Kim, Y.S. Physicochemical properties and antioxidant activity of Korean cactus (Opuntia humifusa) cladodes. Hortic. Environ. Biotechnol. 2013, 54, 288–295. [Google Scholar] [CrossRef]
- El-Mostafa, K.; Kharrassi, Y.; Badreddine, A.; Andreoletti, P.; Vamecq, J.; El Kebbaj, M.S.; Latruffe, N.; Lizard, G.; Nasser, B.; Cherkaoui-Malki, M. Nopal Cactus (Opuntia ficus-indica) as a source of bioactive compounds for nutrition, health and disease. Molecules 2014, 19, 14879–14901. [Google Scholar] [CrossRef]
- Farvid, M.S.; Ding, M.; Pan, A.; Sun, Q.; Chiuve, S.E.; Steffen, L.M.; Willett, W.C.; Hu, F.B. Dietary linoleic acid and risk of coronary heart disease: A systematic review and meta-analysis of prospective cohort studies. Circulatio 2014, 130, 1568–1578. [Google Scholar] [CrossRef]
- Chougui, N.; Tamendjari, A.; Hamidj, W.; Hallal, S.; Barras, A.; Richard, T.; Larbat, R. Oil composition and characterization of phenolic compounds of Opuntia ficus-indica seeds. Food Chem. 2013, 139, 796–803. [Google Scholar] [CrossRef]
- Morales, P.; Ramírez-Moreno, E.; Sánchez-Mata, M.C.; Carvalho, A.M.; Ferreira, I.C. Nutritional and antioxidant properties of pulp and seeds of two xoconostle cultivars (Opuntia joconostle F.A.C. Weber ex Diguet and Opuntia matudae Scheinvar) of high consumption in Mexico. Food Res. Int. 2012, 46, 279–285. [Google Scholar] [CrossRef]
- Ramírez-Moreno, E.; Cariño-Cortés, R.; Cruz-Cansino, N.D.S.; Delgado-Olivares, L.; Ariza-Ortega, J.A.; Vanessa YelinaMontañez-Izquierdo, V.Y.; Hernández-Herrero, M.M.; Tomás Filardo-Kerstupp, T. Antioxidant and antimicrobial properties of cactus pear (Opuntia) seed oils. J. Food Qual. 2017, 2017, 1–8. [Google Scholar] [CrossRef]
- Regalado-Rentería, E.; Aguirre-Rivera, J.R.; González-Chávez, M.M.; Sánchez-Sánchez, R.; Martínez-Gutiérrez, F.; Juárez-Flores, B.I. Assessment of extraction methods and biological value of seed oil from eight variants of prickly pear fruit (Opuntia spp.). Waste Biomass Valori. 2018, 11, 1181–1189. [Google Scholar] [CrossRef]
- Mobraten, K.; Haug, T.M.; Kleiveland, C.R.; Lea, T. Omega-3 and omega-6 PUFAs induce the same GPR120-mediated signalling events, but with different kinetics and intensity in Caco-2 cells. Lipids Health Dis. 2013, 12, 101. [Google Scholar] [CrossRef] [PubMed]
- Berraaouan, A.; Ziyyat, A.; Mekhfi, H.; Legssyer, A.; Sindic, M.; Aziz, M.; Bnouham, M. Evaluation of antidiabetic properties of cactus pear seed oil in rats. Pharm. Boil. 2014, 52, 1286–1290. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, B.; Esen, M.; Sangun, M.K.; Coleri, A.; Caliskan, M. Effective antibacterial and antioxidant properties of methanolic extrac of Laururs nobillis seed oil. J. Environ. Biol. 2010, 5, 637–641. [Google Scholar]
- Carovi’c-Stanko, K.; Orli’c, S.; Politeo, O.; Striki’c, F.; Kolak, I.; Milos, M.; Satovic, Z. Composition and antibacterial activities of essential oils of seven Ocimum taxa. Food Chem. 2010, 119, 196–201. [Google Scholar] [CrossRef]
- Tajkarimi, M.M.; Ibrahim, S.A.; Cliver, D.O. Antimicrobial herb and spice compounds in food. Food Cont. 2010, 21, 1199–1218. [Google Scholar] [CrossRef]
- Solórzano-Santos, F.; Miranda-Novales, M.G. Essential oils from aromatic herbs as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 136–141. [Google Scholar] [CrossRef]
- Atteya, A.K.G.; Albalawi, A.N.; El-Serafy, R.S.; Albalawi, K.N.; Bayomy, H.M.; Genaidy, E.A.E. Response of Moringa oleifera seeds and fixed oil production to vermicompost and npk fertilizers under calcareous soil con-ditions. Plants 2021, 10, 1998. [Google Scholar] [CrossRef]
- Genaidy, E.A.E.; Atteya, A.K.G.; Adss, I.A.A. Increase the economic value of the jojoba (Simmondsia chinensis) yield using evaluation of distinctive clones grown under the Egyptian environmental conditions. J. Agric. Sci. Technol. 2016, 12, 145–165. [Google Scholar]
- Atteya, A.K.G.; Genaidy, E.A.E.; Zahran, H.A. Chemical constituents and yield of Simmondsia chinensis plants as affected by foliar application of gibberellic acid and zinc sulphate. Biosci. Res. 2018, 15, 1528–1541. [Google Scholar]
- Ali, B.; Hayat, S.; Ahmad, A. 28-Homobrassinolide ameliorates the salt stress in chickpea (Cicer arietinum). Environ. Exp. Bot. 2007, 59, 33–41. [Google Scholar] [CrossRef]
- Caño-Delgado, A.; Yin, Y.; Yu, C.; Vafeados, D.; Mora-García, S.; Cheng, J.C.; Nam, K.H.; Li, J.; Chory, J. BRL1 and BRL3 are novel brassinosteroid receptors that function in vascular differentiation in Arabidopsis. Development 2004, 131, 5341–5351. [Google Scholar] [CrossRef]
- Saygideger, S.; Deniz, F. Efect of 24-epibrassinolide on biomass, growth and free proline concentration in Spirulina platensis (Cyanophyta) under NaCl stress. Plant Growth Regul. 2008, 56, 219–223. [Google Scholar] [CrossRef]
- Hayat, S.; Yadav, S.; Ali, B.; Ahmad, A. Interactive effect of nitric oxide and brassinosteroids on photosynthesis and the antioxidant system of Lycopersicon esculentum. Russ. J. Plant Physiol. 2010, 57, 212–221. [Google Scholar] [CrossRef]
- Fathima, M.S.A.; Johnson, M.; Lingakumar, K. Effect of crude brassinosteroid extract on growth and biochemical changes of Gossypium hirsutum L. and Vigna mungo L. J. Stress Physiol. Biochem. 2011, 7, 324–334. [Google Scholar]
- Sasse, J.M. Physiological Actions of Brassinosteroids: An Update. J. Plant. Growth Regul. 2003, 22, 276–288. [Google Scholar] [CrossRef]
- Asha, A.; Lingakumar, K. Effect of 24-epibrassinolide spray on vegetative growth, pigment composition and biochemical constituents of Vigna mungo (L.) Hepper (Blackgram). J. Global. Biosci. 2015, 4, 2007–2012. [Google Scholar]
- Helaly, M.N.; El-Hoseiny, H.M.; Elsheery, N.I.; Kalaji, H.M.; Santos-Villalobos, S.D.L.; Wróbel, J.; Hassan, I.F.; Gaballah, M.S.; Abdelrhman, L.A.; Mira, A.M.; et al. 5-aminolevulinic acid and 24-epibrassinolide improve the drought stress resilience and productivity of banana plants. Plants 2022, 11, 743. [Google Scholar] [CrossRef]
- Thakur, M.; Sohal, B.S. Role of elicitors in inducing resistance in plants against pathogen infection: A review. ISRN Biochem. 2013, 2013, 1–10. [Google Scholar] [CrossRef]
- Malikova, J.; Swaczynova, J.; Kolar, Z.; Strnad, M. Anticancer and antiproliferative activity of natural brassinosteroids. Phytochemistry 2008, 69, 418–426. [Google Scholar] [CrossRef]
- Xia, X.J.; Huang, Y.Y.; Wang, L.; Huang, L.F.; Yu, Y.L.; Zhou, Y.H.; Yu, J.Q. Pesticides induced depression of photosynthesis was alleviated by 24-epibrassinolide pretreatment in Cucumis sativus L. Pestic. Biochem. Physiol. 2006, 86, 42–48. [Google Scholar] [CrossRef]
- Jackson, M.L. Methods of Chemical Analysis; Prentice Hall of India: New Delhi, India, 1973. [Google Scholar]
- Cottenie, A.; Verloo, M.; Kikens, L. Chemical Analysis of Plants and Soils; RUG Laboratory of Analytical and Agrochemistry: Gent, Belgium, 1982. [Google Scholar]
- Association of Official Analytical Chemists. Official Methods of Analysis, 12th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1980. [Google Scholar]
- Wintermans, J.F.G.M.; Mats, D.E. Spectrophtometeric characteristic of chlorophyll and their pheophytins in ethanol. Biochem. Biophys. Acta 1965, 109, 448–453. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdicphosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Kim, D.; Chun, O.; Kim, Y.; Moon, H.; Lee, C. Quantification of phenolics and their antioxidant capacity in fresh plums. J. Agric. Food Chem. 2003, 51, 6509–6515. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Lebensm. Wiss. Technol. 1995, 26, 25–30. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT User’s Guide; Release 6.03 Edition; SAS Institute: Cary, NC, USA, 1988. [Google Scholar]
- Potgieter, J.P.; Mashope, B.K. Cactus pear (Opuntia spp.) germplasm conservation in South Africa. Acta. Hort. 2009, 811, 47–54. [Google Scholar] [CrossRef]
- El-Kossori, R.L.; Villaume, C.; El Boustani, E.; Sauvaire, Y.; Méjean, L. Composition of pulp, skin and seeds of prickly pears fruit (Opuntia ficus indica sp.). Plant Foods Hum. Nutr. 1998, 52, 263–270. [Google Scholar] [CrossRef]
- Tanveer, M. Role of 24-Epibrassinolide in ınducing thermo-tolerance in plants. J. Plant Growth Regul. 2019, 38, 945–955. [Google Scholar] [CrossRef]
- Li, J.; Jin, H. Regulation of brassinosteroid signaling. Trends Plant Sci. 2007, 12, 37–41. [Google Scholar] [CrossRef]
- Divi, U.K.; Krishna, P. Brassinosteroid: A biotechnological target for enhancing crop yield and stress tolerance. New Biotechol. 2009, 26, 131–136. [Google Scholar] [CrossRef]
- Clouse, S.D.; Zurek, D. Molecular Analysis of Brassinolide Action in Plant Growth and Development; Cutler, H.G., Yokota, T., Adam, G., Eds.; In Brassinosteroids; American Chemical Society: Washington, DC, USA, 1991; pp. 122–140. [Google Scholar]
- Oh, M.H.; Clause, S.D. Brassinolide affects the rate of cell division in isolated leaf protoplasts of Petunia hybrida. Plant Cell Rep. 1998, 17, 921–924. [Google Scholar] [CrossRef]
- Eveland, A.L.; Jackson, D.P. Sugars, signalling, and plant development. J. Exp. Bot. 2011, 63, 3367–3377. [Google Scholar] [CrossRef]
- Vert, G.; Chory, J. Crosstalk in cellular signaling: Background noise or the real thing? Dev. Cell 2011, 21, 985–991. [Google Scholar] [CrossRef]
- El-Serafy, R.S.; El-Sheshtawy, A.-N.A.; Atteya, A.K.; Al-Hashimi, A.; Abbasi, A.M.; Al-Ashkar, I. Seed priming with silicon as a potential to increase salt stress tolerance in Lathyrus odoratus. Plants 2021, 10, 2140. [Google Scholar] [CrossRef]
- Atteya, A.K.G.; Albalawi, A.N.; Bayomy, H.M.; Alamri, E.S.; Genaidy, E.A.E. Maximizing Leaves, Inflores-cences, and Chemical Composition Production of Moringa oleifera Trees under Calcareous Soil Conditions. Plants 2022, 11, 234. [Google Scholar] [CrossRef]
- Atteya, A.K.; Albalawi, A.N.; Bayomy, H.M.; El Moataz Bellah, A.; Ghozlan, M.H.; Genaidy, E.A. Impact of planting density and soaking seeds in melatonin solution on yield, secondary products content and antimicrobial activity of lovage plant. Saudi J. Biol. Sci. 2022, 29, 2656–2673. [Google Scholar] [CrossRef]
- Tong, H.; Xiao, Y.; Liu, D.; Gao, S.; Liu, L.; Yin, Y.; Jin, Y.; Qian, Q.; Chu, C. Brassinosteroid regulates cell elongation by modulating gibberellin metabolism in rice. Plant Cell 2014, 26, 4376–4393. [Google Scholar] [CrossRef]
- Felner, M. Recent progress in brassinosteroid research: Hormone perception and signal transduction. In Brassinosteroids: Bioactivity and Crop Productivity; Hayat, S., Ahmad, A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 69–86. [Google Scholar]
- Cosgrove, D. Relaxation in a high stress environment: The molecular basis of extensible cell walls and enlargement. Plant Cell 1997, 9, 1031–1041. [Google Scholar] [CrossRef]
- Yu, J.Q.; Huang, L.F.; Hu, W.H.; Zhou, Y.H.; Mao, W.H.; Ye, S.F.; Nogues, S. A role for brassinosteroids in the regulation of photosynthesis in Cucumis sativus. J. Exp. Bot. 2004, 55, 1135–1143. [Google Scholar] [CrossRef]
- Anuradha, S.; Rao, S.S.R. Effect of brassinosteroids on salinity stress induced inhibition of seed germination and seedling growth of rice (Oryza sativa L.). Plant Growth Regul. 2001, 33, 151–153. [Google Scholar] [CrossRef]
- Bajguz, A.; Czerpak, R. Physiological and biochemical role of brassinosteroids and their structure activity relationship in the green alga Chlorella vulgaris Beijerinck (Chlorophyceae). Plant Growth Regul. 1998, 17, 131–139. [Google Scholar] [CrossRef]
- Swamy, K.N.; Rao, S.S.R. Effect of 24-epibrassinolide on growth, photosynthesis, and essential oil content of Pelargonium graveolens (L.) Herit. Russ. J. Plant Physiol. 2009, 56, 616–620. [Google Scholar]
- Houimli, S.M.; Denden, M.; Mouhandes, B.D. Effects of 24-epi-brassinolide on growth, chlorophyll, electrolyte leakage and proline by pepper plants under NaCl-stress. Eurasia. J. BioSci. 2010, 4, 96–104. [Google Scholar]
- Shahid, M.A.; Pervez, M.A.; Balal, R.M.; Mattson, N.S.; Rashid, A.; Ahmad, R.; Ayyub, C.M.; Abbas, T. Brassinosteroid (24-epibrassinolide) enhances growth and alleviates the deleterious effects induced by salt stress in pea (Pisum sativum L.). Aust. J. Crop. Sci. 2011, 5, 500–510. [Google Scholar]
- Vogler, F.; Schmalzl, C.; Englhart, M.; Bircheneder, M.; Sprunck, S. Brassinosteroids promote Arabidopsis pollen germination and growth. Plant Reprod. 2014, 27, 153–167. [Google Scholar] [CrossRef]
- Maita, S.; Sotomayor, C. The effect of three plant bioregulators on pollen germination, pollen tube growth and fruit set in almond [Prunus dulcis (Mill.) DA Webb] cvs. Non Pareil Carmel. Electron. J. Biotechnol. 2015, 18, 381–386. [Google Scholar] [CrossRef]
- Khamsuk, O.; Sonjaroon, W.; Suwanwong, S.; Jutamanee, K.; Suksamrarn, A. Effects of 24-epibrassinolide and the synthetic brassinosteroid mimic on chili pepper under drought. Acta Physiol. Plant 2018, 40, 106. [Google Scholar] [CrossRef]
- Kang, Y.Y.; Guo, S.S.R. Role of brassinosteroids on horticultural crops. In Brassinosteroids: A Class of Plant Hormone; Springer: New York, NY, USA, 2011; pp. 143–192. [Google Scholar]
- Ozga, J.; Dennis, M. Hormonal interactions in fruit development. J. Plant Growth Regul. 2003, 22, 73–81. [Google Scholar] [CrossRef]
- Brenner, M.L.; Cheikh, N. The role of hormones in photosynthate partitioning and seed filling. In Plant Hormones: Physiology, Biochemistry, and Molecular Biology, 2nd ed.; Davis, P.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; pp. 649–670. [Google Scholar]
- Naeema, M.; Mohd, I.; Masidur, A.M.; Aftaba, T.; Masroor, M.; Khana, A.; Moinuddin. Brassinosteroid mediated enrichment in yield attributes, active constituents and essential oil production in Mentha arvensis L. Russ. Agric. Sci. 2012, 38, 106–113. [Google Scholar] [CrossRef]
- Maia, N.B.; Bovi, O.A.; Zullo, M.A.T.; Perecin, M.B.; Granja, N.P.; Carmello, Q.A.C.; Robaina, C.; Coll, F. Hydroponic cultivation of mint and vetiver with spirostane analogues of brassinosteroids. Acta Hortic. (ISHS) 2004, 644, 55–59. [Google Scholar] [CrossRef]
- Swamy, K.N.; Rao, S.S.R. Influence of 28-homobrassinolide on growth, photosynthesis metabolite and essential oil content of geranium [Pelargonium graveolens (L.) herit]. Am. J. Plant Physiol. 2008, 3, 173–179. [Google Scholar] [CrossRef][Green Version]
- Farooqi, H.A.; Sharma, S. Effect of growth retardants on growth and essential oil content in Japanese mint. Plant Growth Regul. 1988, 7, 39–45. [Google Scholar] [CrossRef]
- Santoro, M.V.; Nievas, F.; Zygadlo, J.; Giordano, W.; Banchio, E. Effects of growth regulators on biomass and the production of secondary metabolites in Peppermint (Mentha piperita) micropropagated in vitro. Am. J. Plant Sci. 2013, 4, 49–55. [Google Scholar] [CrossRef]
- Ennouri, M.; Evelyne, B.; Laurence, B.; Hamadi, A. Fatty acid composition and rheological behavior of prickly pear seed oils. Food Chem. 2005, 93, 431–437. [Google Scholar] [CrossRef]
- Taoufik, F.; Zine, S.; El Hadek, M.; Hassani, L.M.I.; Gharby, S.; Harhar, H.; Matthaus, B. Oil content and main constituents of cactus seed oils Opuntia Ficus Indica of different origin in Morocco. Mediterr. J. Nutr. Metab. 2015, 8, 85–92. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | Unit | Value |
|---|---|---|
| Clay | % | 19 |
| Sand | % | 38 |
| Silt | % | 43 |
| Texture class | Clay loam | |
| OM | % | 1.3 |
| pH | 7.9 | |
| Ece | ds m−1 | 2.3 |
| CaCO3 | % | 1.8 |
| Soluble ions (meq L−1) | ||
| HCO−3 | 1.4 | |
| Cl− | 13.2 | |
| SO−2 | 5.21 | |
| Ca+2 | 8.82 | |
| Mg+2 | 4.03 | |
| Na+ | 4.55 | |
| K+ | 0.32 | |
| Fatty Acids | Brassinolide Concentration | |||
|---|---|---|---|---|
| Untreated | 1 mg L−1 | 3 mg L−1 | 5 mg L−1 | |
| Stearic acid (C18:0) | 3.09 ± 0.14 | 3.13 ± 0.13 | 3.22 ± 0.14 | 3.28 ± 0.16 |
| Linoleic acid (C18:2) | 59.09 ± 2.52 | 60.37 ± 2.63 | 61.86 ± 2.62 | 62.05 ± 2.62 |
| α-Linolenic acid (C18:3) | 0.20 ± 0.01 | 0.18 ± 0.01 | 0.14 ± 0.01 | 0.12 ± 0.01 |
| Oleic acid (C18:1) | 20.49 ± 0.92 | 20.88 ± 0.93 | 21.1 ± 0.9 | 21.37 ± 0.85 |
| Palmitic acid (C16:0) | 10.26 ± 0.44 | 10.55 ± 0.45 | 10.94 ± 0.47 | 11.12 ± 0.50 |
| Palmitoleic acid (C16:1) | 0.34 ± 0.01 | 0.37 ± 0.02 | 0.39 ± 0.02 | 0.32 ± 0.03 |
| Arachidic acid (C20:0) | 0.10 ± 0.004 | 0.20 ± 0.008 | 0.29 ± 0.012 | 0.31 ± 0.01 |
| Paullinic acid (C20:1) | 0.26 ± 0.011 | 0.25 ± 0.011 | 0.23± 0.010 | 0.17 ± 0.01 |
| Behenic acid (C22:0) | 0.08 ± 0.003 | 0.09 ± 0.004 | 0.11 ± 0.005 | 0.12 ± 0.005 |
| Lignoceric acid (C24:0) | 0.07 ± 0.003 | 0.08 ± 0.003 | 0.09 ± 0.004 | 0.09 ± 0.004 |
| Total identified fatty acids | 98.98 | 96.10 | 98.37 | 98.95 |
| Total unsaturated fatty acids | 80.83 | 82.05 | 83.72 | 84.03 |
| Total saturated fatty acids | 13.60 | 14.05 | 14.65 | 14.92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atteya, A.K.G.; El-Serafy, R.S.; El-Zabalawy, K.M.; Elhakem, A.; Genaidy, E.A.E. Brassinolide Maximized the Fruit and Oil Yield, Induced the Secondary Metabolites, and Stimulated Linoleic Acid Synthesis of Opuntia ficus-indica Oil. Horticulturae 2022, 8, 452. https://doi.org/10.3390/horticulturae8050452
Atteya AKG, El-Serafy RS, El-Zabalawy KM, Elhakem A, Genaidy EAE. Brassinolide Maximized the Fruit and Oil Yield, Induced the Secondary Metabolites, and Stimulated Linoleic Acid Synthesis of Opuntia ficus-indica Oil. Horticulturae. 2022; 8(5):452. https://doi.org/10.3390/horticulturae8050452
Chicago/Turabian StyleAtteya, Amira K. G., Rasha S. El-Serafy, Khaled M. El-Zabalawy, Abeer Elhakem, and Esmail A. E. Genaidy. 2022. "Brassinolide Maximized the Fruit and Oil Yield, Induced the Secondary Metabolites, and Stimulated Linoleic Acid Synthesis of Opuntia ficus-indica Oil" Horticulturae 8, no. 5: 452. https://doi.org/10.3390/horticulturae8050452
APA StyleAtteya, A. K. G., El-Serafy, R. S., El-Zabalawy, K. M., Elhakem, A., & Genaidy, E. A. E. (2022). Brassinolide Maximized the Fruit and Oil Yield, Induced the Secondary Metabolites, and Stimulated Linoleic Acid Synthesis of Opuntia ficus-indica Oil. Horticulturae, 8(5), 452. https://doi.org/10.3390/horticulturae8050452