
Characteristics of Chili (Capsicum annuum L.) That Are Resistant and Susceptible to Oriental Fruit Fly (Bactrocera dorsalis Hendel) Infestation



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Plant Materials
2.3. Resistance of Chili to Infestation of Fruit Flies
2.4. Morphological Characteristics of Chili Fruit
2.5. Analysis of Volatile Compounds
2.6. Oriental Fruit Fly Response to Selected Volatile Compounds
2.7. Analysis of Nutritional Content
2.7.1. Water Content
2.7.2. Carbohydrate Content
2.7.3. Fiber Content
2.8. Data Analysis
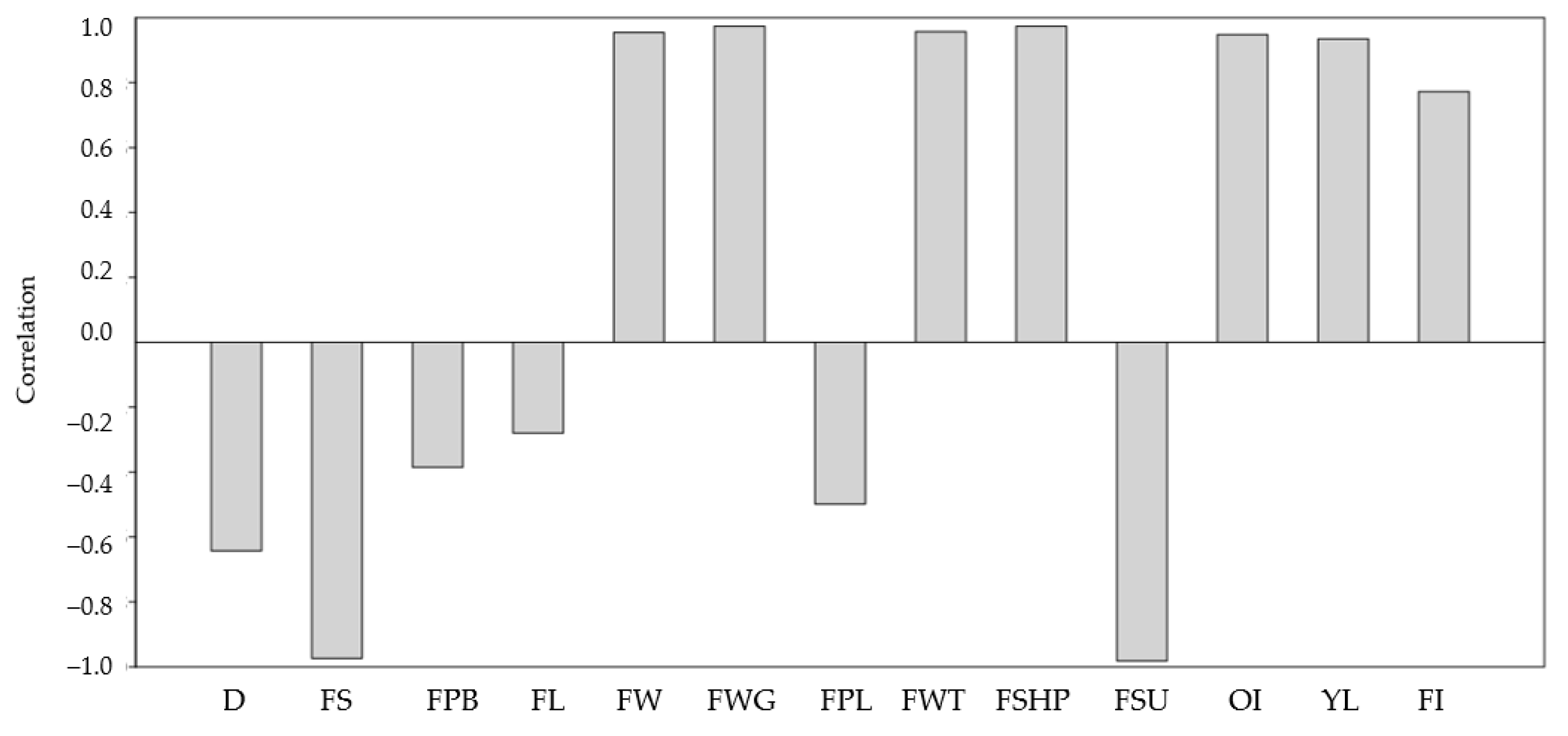
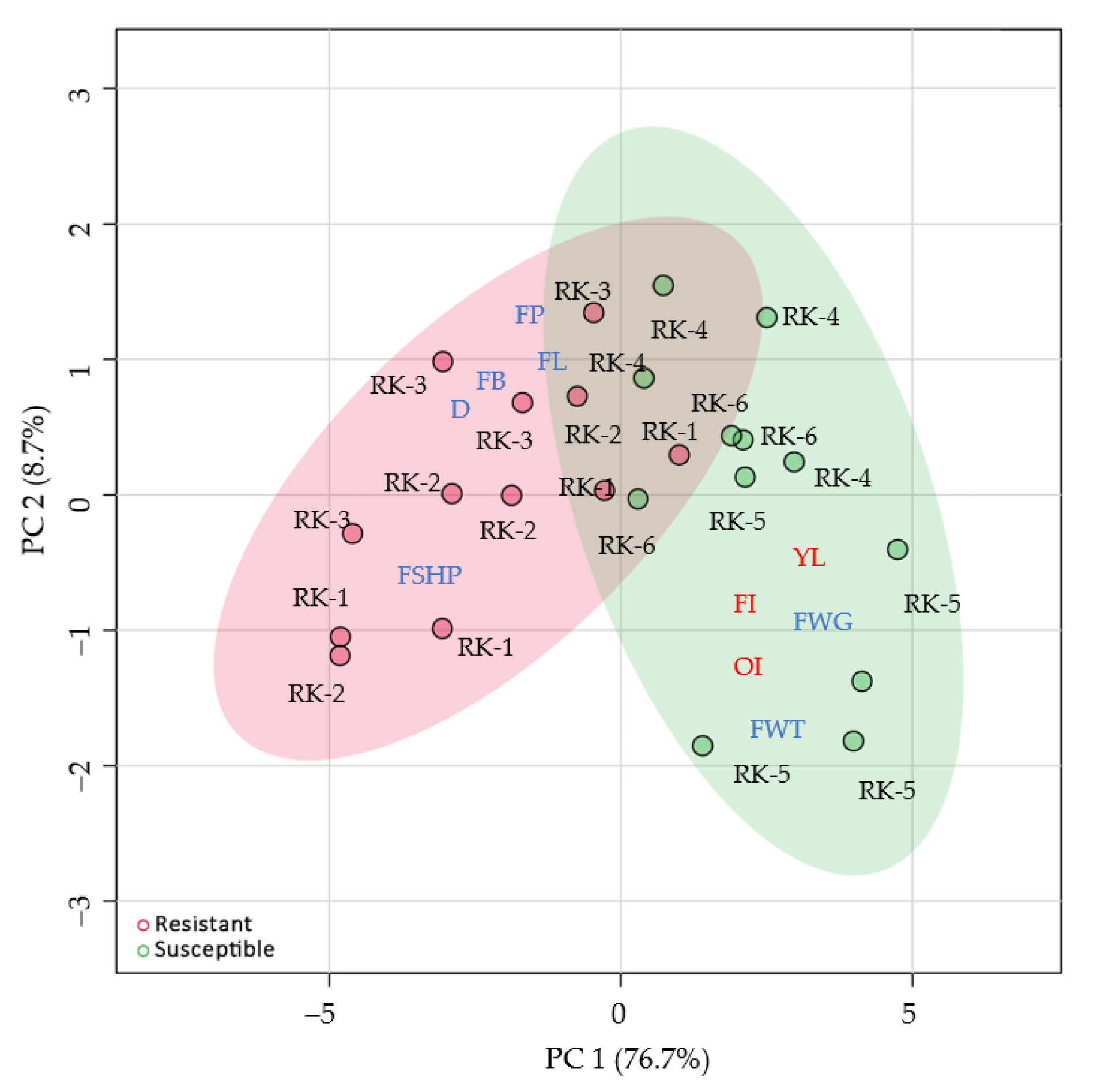
3. Results
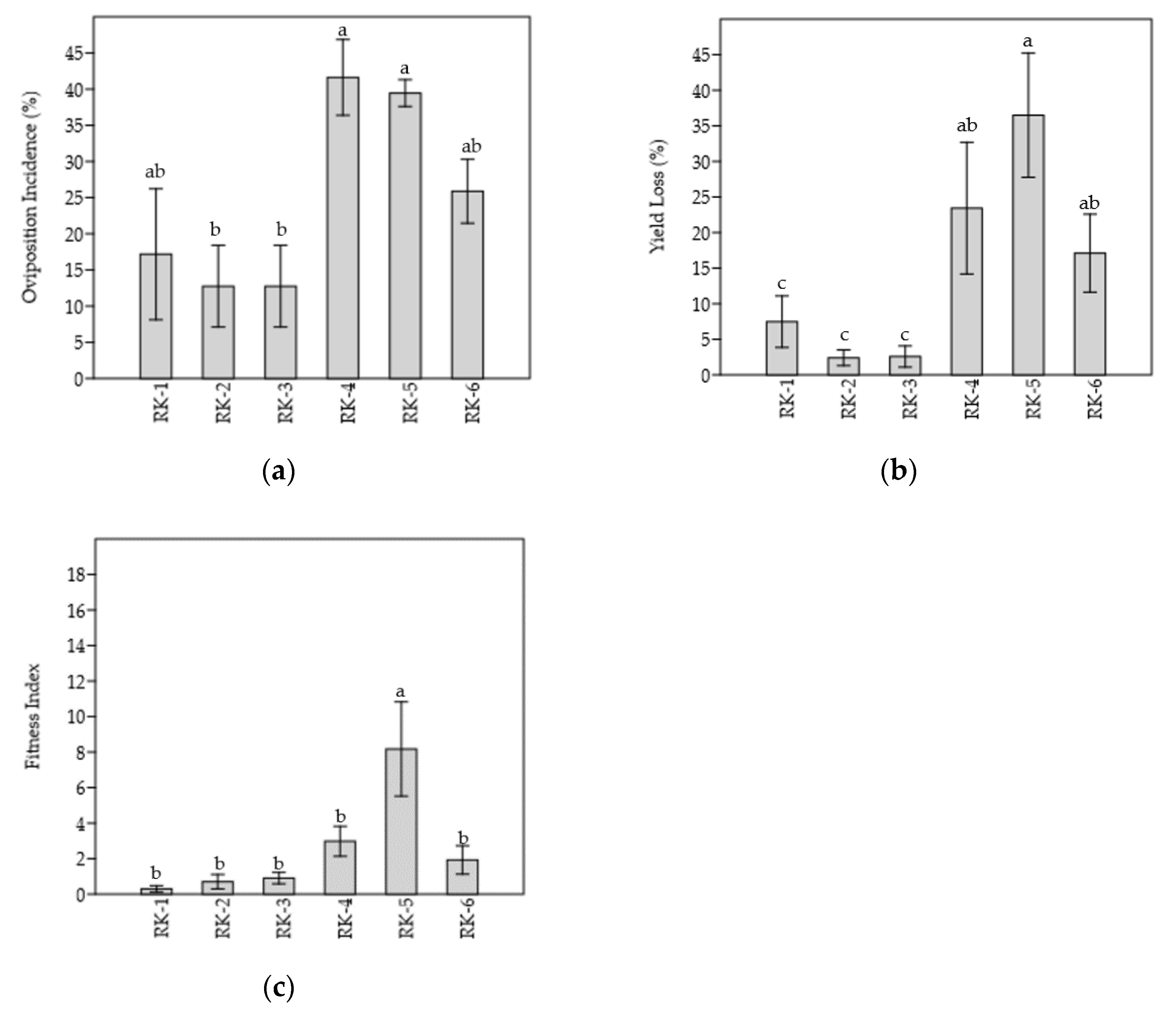
3.1. Resistance Characteristics
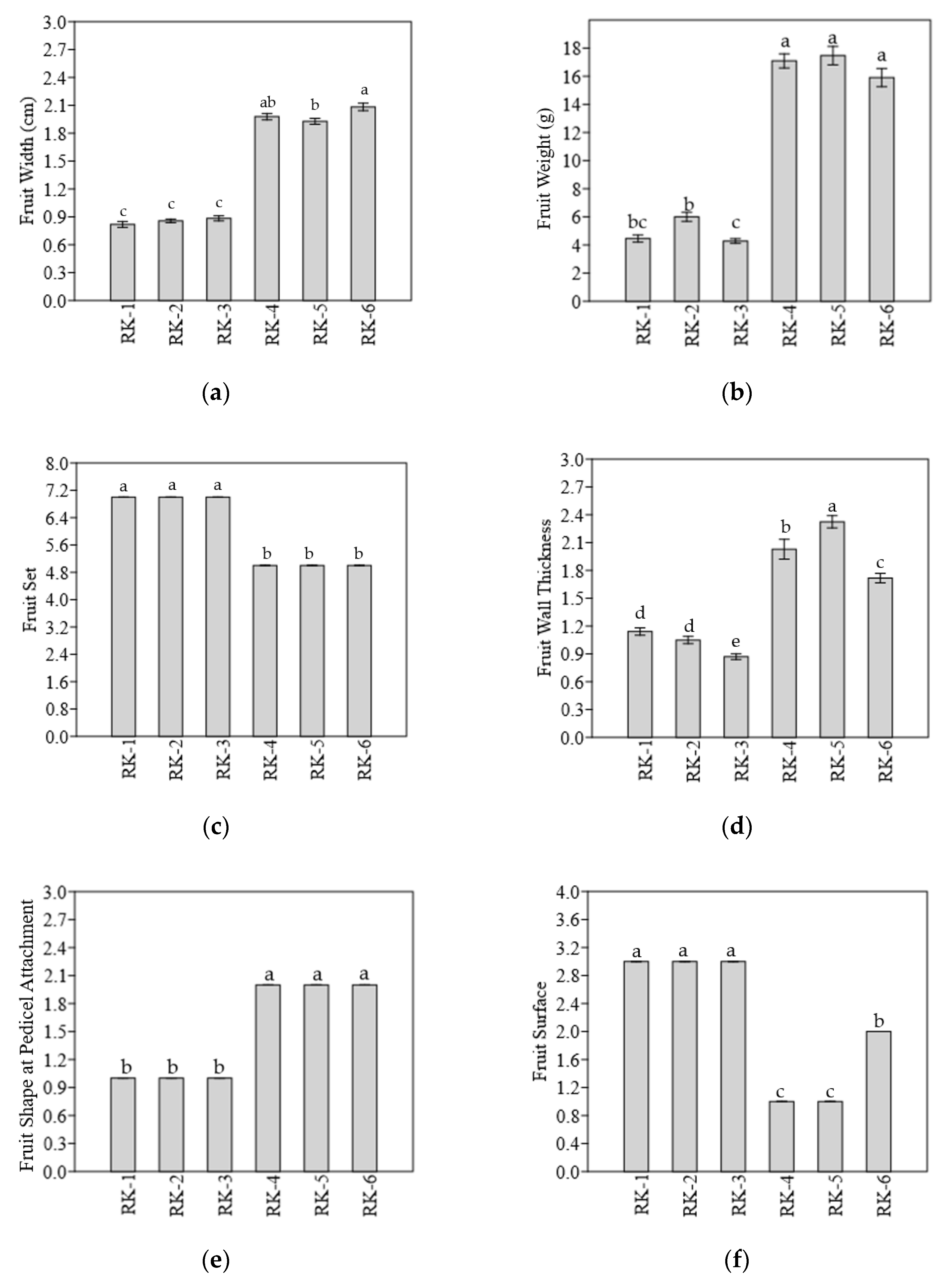
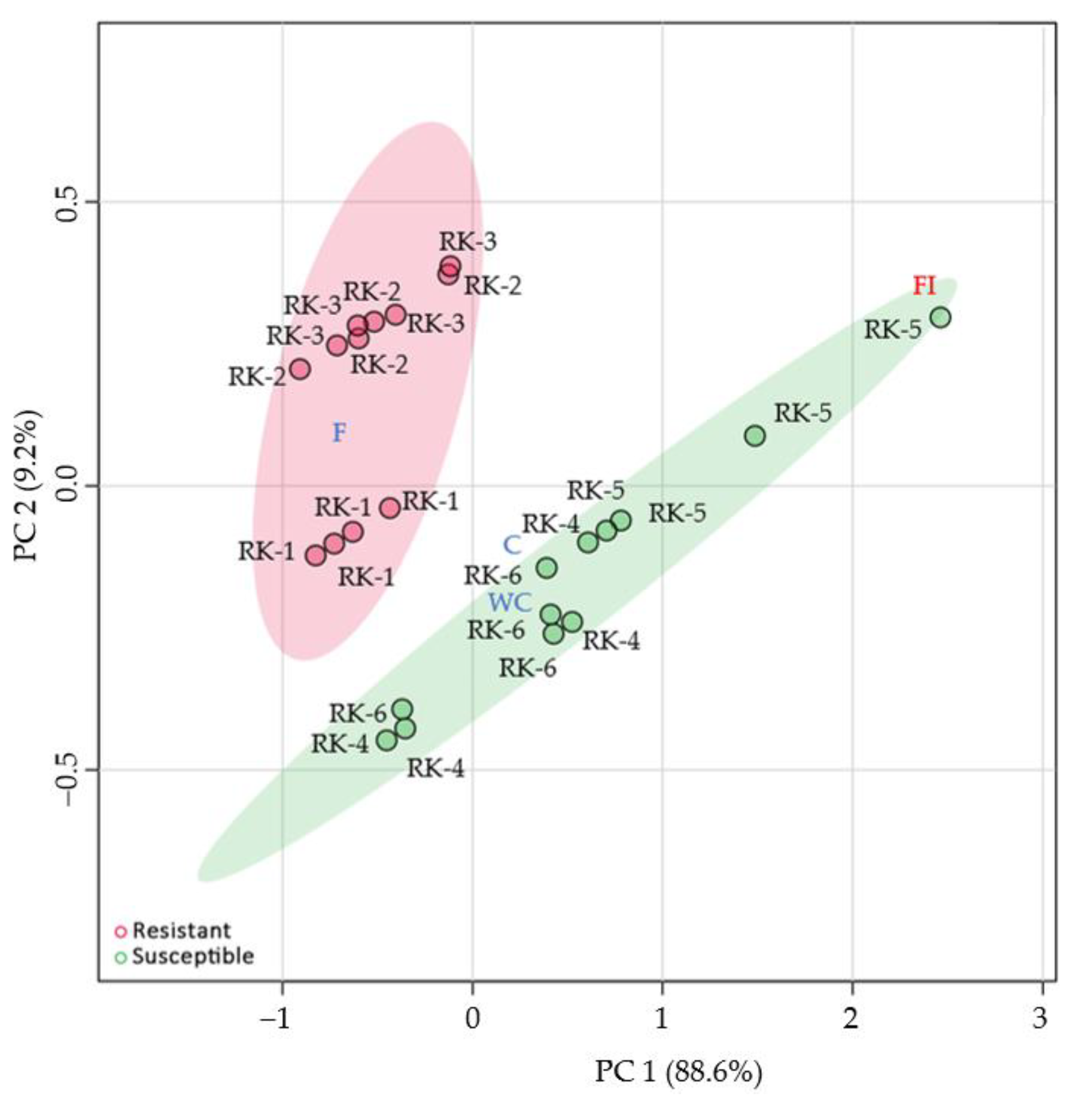
3.2. Morphological Characteristics of Chili Fruit
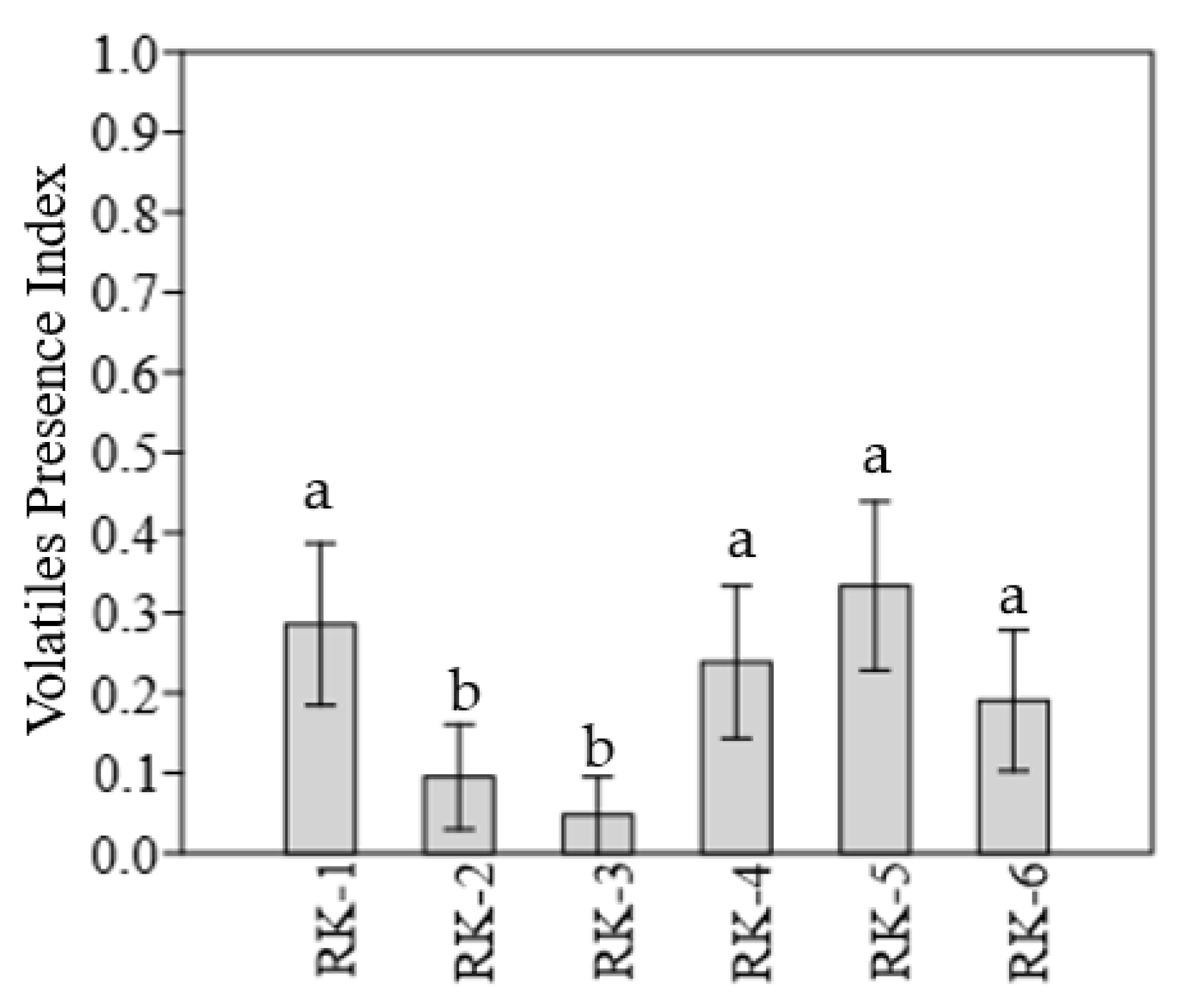
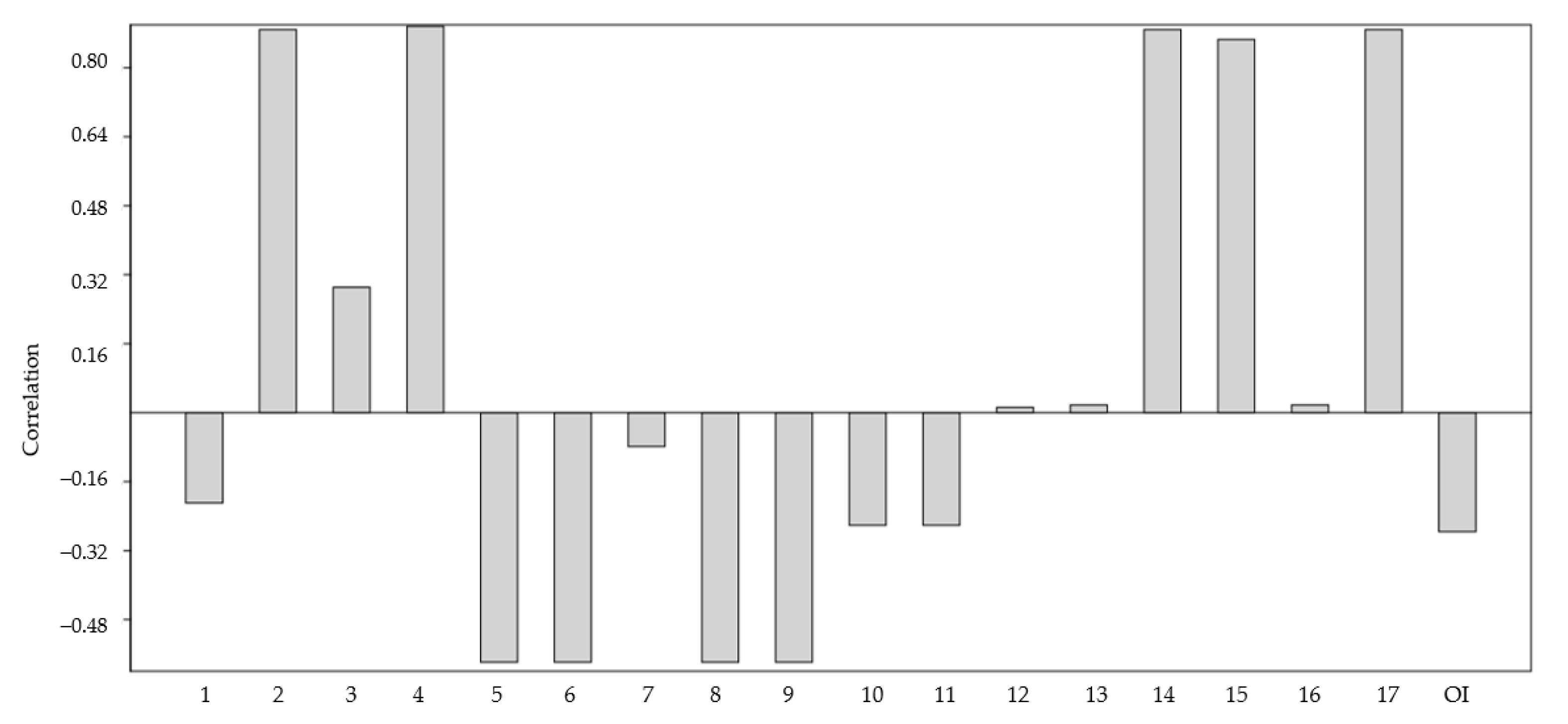
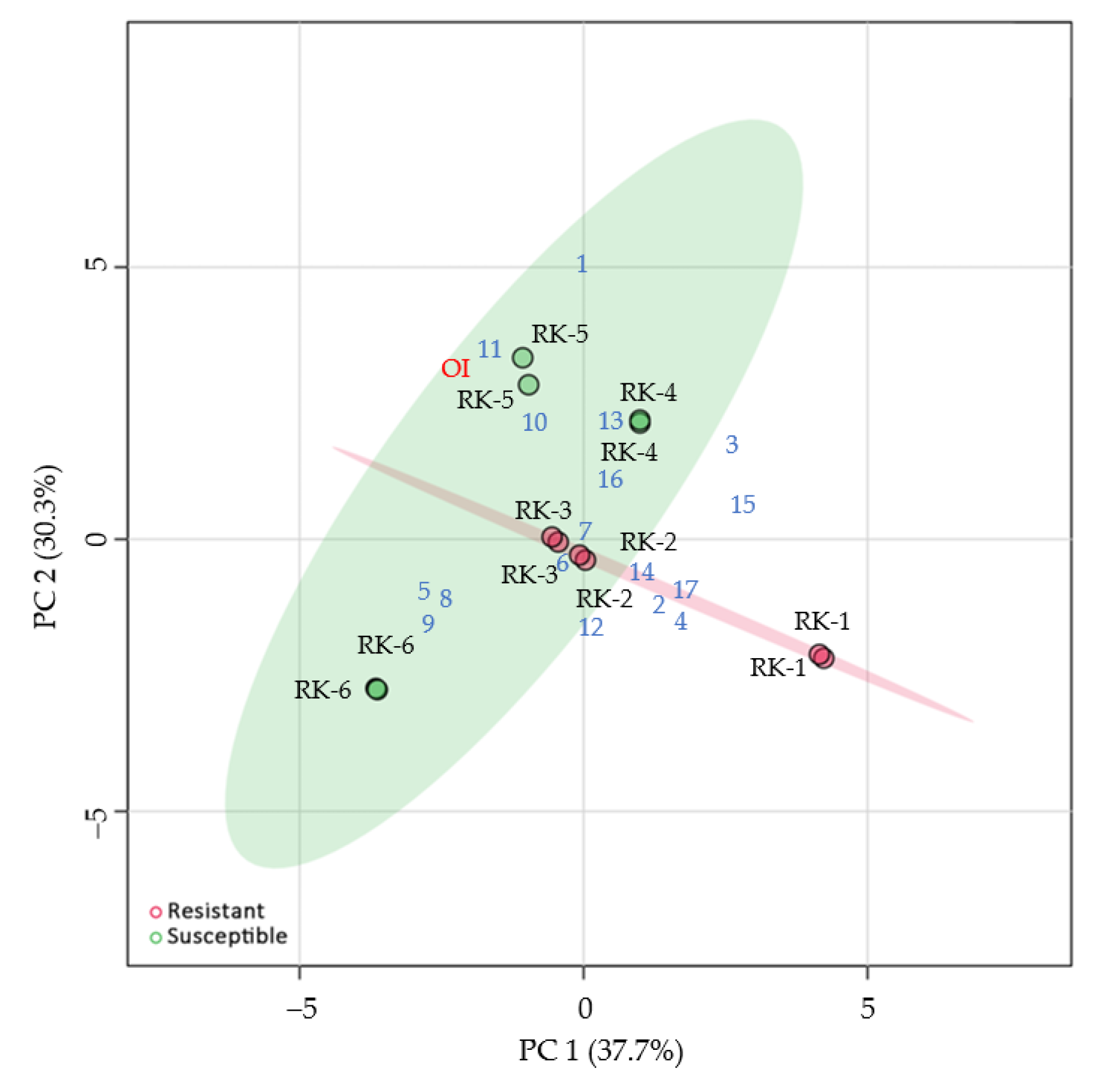
3.3. Characteristics of Volatile Compounds
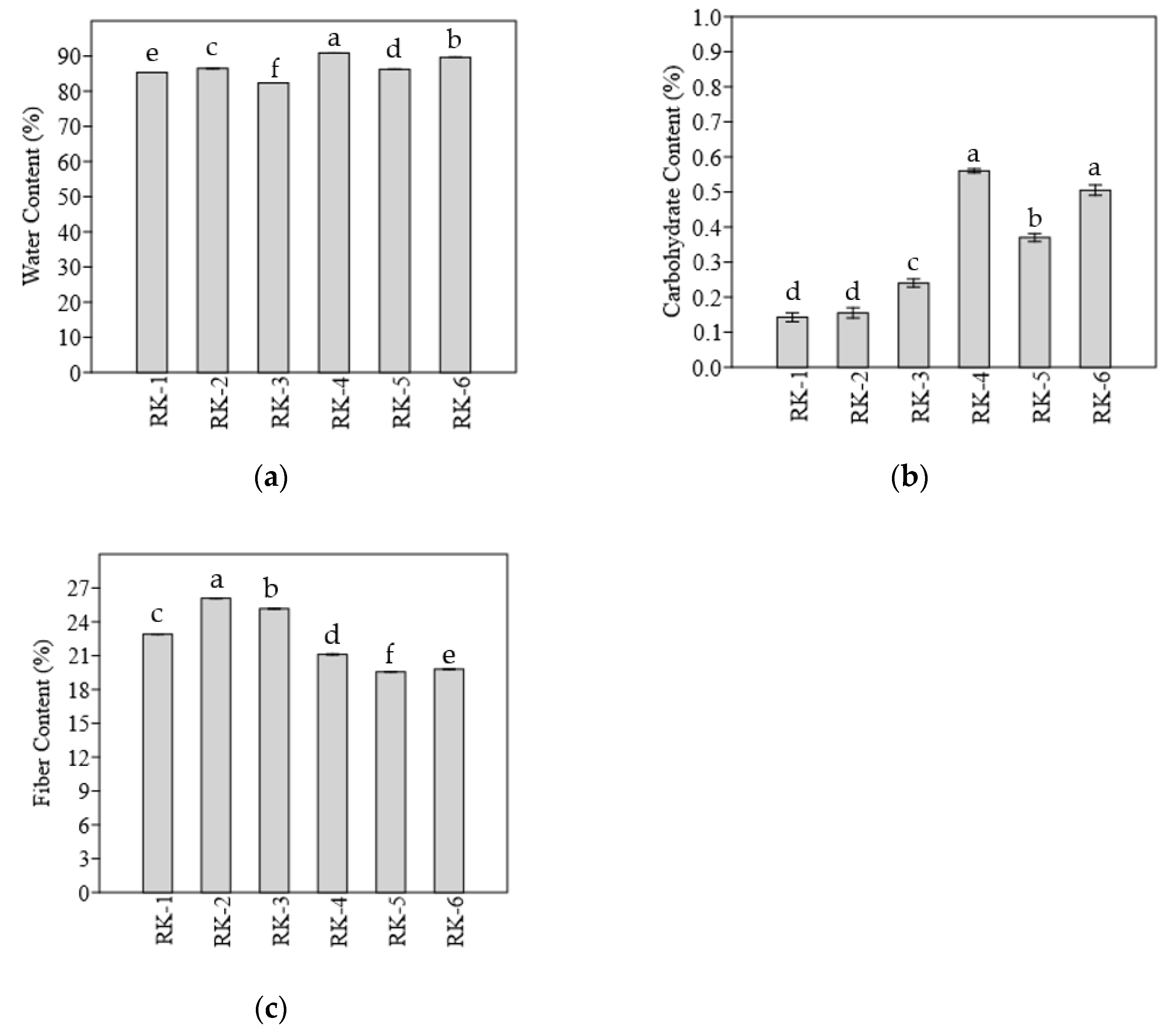
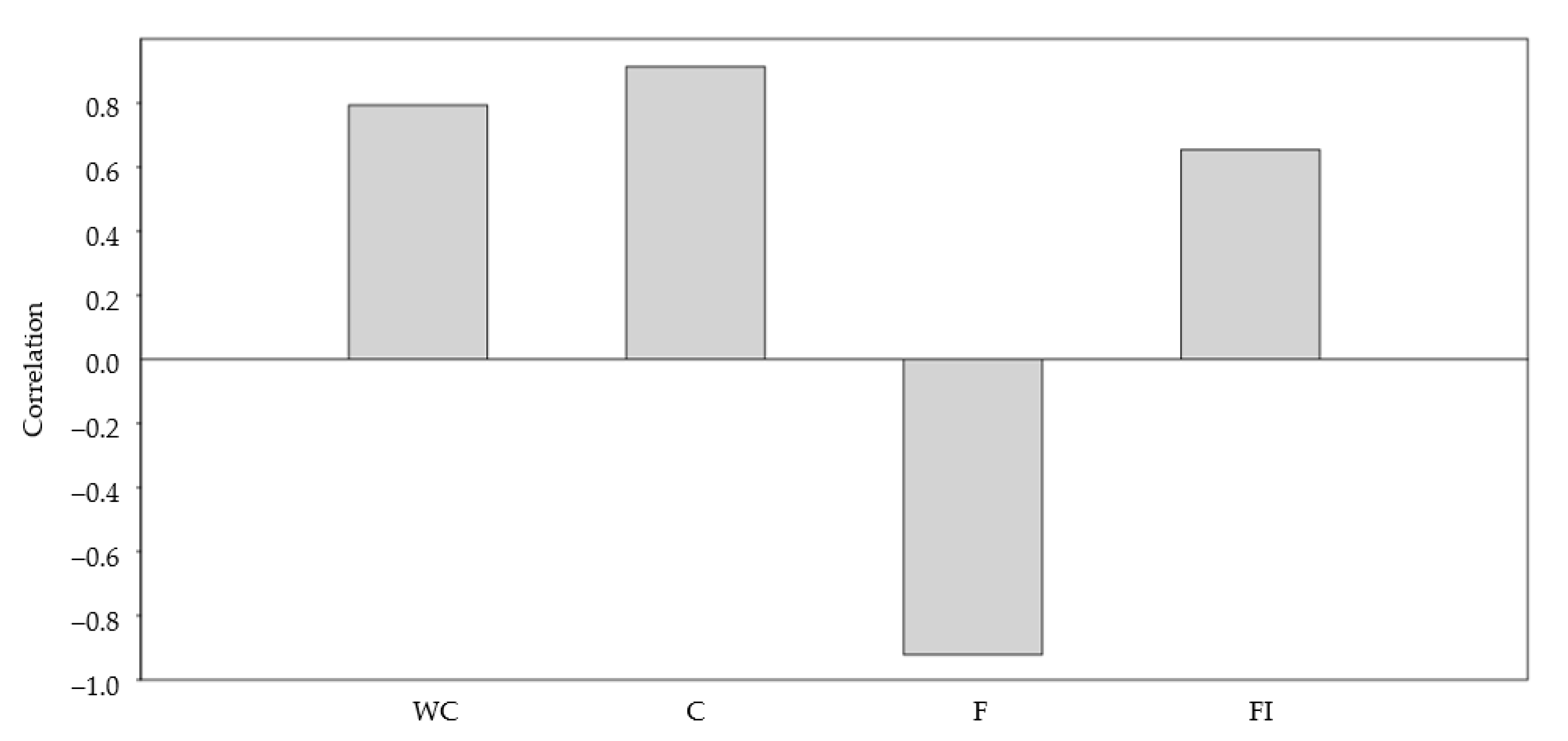
3.4. Nutritional Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Said, A.E.; Fatahuddin, A.; Nasruddin, A. Effect of sticky trap color and height on the capture of adult oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) on chili pepper. Am. J. Agric. Biol. Sci. 2017, 12, 13–17. [Google Scholar] [CrossRef]
- Hasyim, A.; Setiawati, W.; Sutarya, R. Screening for resistance to anthracnose caused by Colletotrichum acutatum in chili pepper (Capsicum annuum L.) in Kediri, East Java. Adv. Agric. Bot. 2014, 6, 104–118. [Google Scholar]
- Syamsudin, T.S.; Faizal, A.; Kirana, R. Dataset on antixenosis and antibiosis of chili fruit by fruit fly (Bactrocera dorsalis) infestation. Data Brief 2019, 23, 103758. [Google Scholar] [CrossRef] [PubMed]
- Bosland, P.W.; Votava, E.J. Peppers: Vegetable and Spice Capsicums; CABI: Boston, MA, USA, 2012. [Google Scholar]
- Wahyuni, Y.; Ballester, A.-R.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Secondary metabolites of Capsicum species and their importance in the human diet. J. Nat. Prod. 2013, 76, 783–793. [Google Scholar] [CrossRef]
- Aliu, S.A.; Rusinovci, I.; Fetahu, S.; Kaçiu, S.; Zeka, D. Assessment of morphological variability and chemical composition of some local pepper (Capsicum annuum L.) populations on the area of Kosovo. Acta Agric. Slov. 2017, 109, 205–213. [Google Scholar] [CrossRef][Green Version]
- Prabaningrum, L.; Moekasan, T.K. Pest and disease management on hot pepper cultivation in high land. J. Hortic. 2014, 24, 179–188. [Google Scholar]
- IPGRI; AVRDC; CATIE. Descriptors for Capsicum (Capsicum spp.); International Plant Genetic Resources Institute: Rome, Italy; The Asian Vegetable Research and Development Center: Taipei, Taiwan; The Centro Agronómico Tropical de Investigación y Enseñanza: Turrialba, Costa Rica, 1995; Volume 110. [Google Scholar]
- Junior, S.B.; Tavares, A.M.; Teixeira Filho, J.; Zini, C.A.; Godoy, H.T. Analysis of the volatile compounds of Brazilian chilli peppers (Capsicum spp.) at two stages of maturity by solid phase micro-extraction and gas chromatography-mass spectrometry. Food Res. Int. 2012, 48, 98–107. [Google Scholar] [CrossRef]
- Kirana, R.; Karyadi, A.K.; Faizal, A.; Syamsudin, T.S. Dataset on volatile compounds in susceptible and resistant chili variety to fruit fly infestation. Data Brief 2019, 22, 234–236. [Google Scholar] [CrossRef] [PubMed]
- Luning, P.A.; de Rijk, T.; Wichers, H.J.; Roozen, J.P. Gas chromatography, mass spectrometry, and sniffing port analyses of volatile compounds of fresh bell peppers (Capsicum annuum) at different ripening stages. J. Agric. Food Chem. 1994, 42, 977–983. [Google Scholar] [CrossRef]
- Faizal, A.; Esyanti, R.R.; Aulianisa, E.N.; Santoso, E.; Turjaman, M. Formation of agarwood from Aquilaria malaccensis in response to inoculation of local strains of Fusarium solani. Trees 2017, 31, 189–197. [Google Scholar] [CrossRef]
- Jayanthi, P.D.K.; Woodcock, C.M.; Caulfield, J.; Birkett, M.A.; Bruce, T.J. Isolation and identification of host cues from mango, Mangifera indica, that attract gravid female oriental fruit fly, Bactrocera dorsalis. J. Chem. Ecol. 2012, 38, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Aluja, M.; Arredondo, J.; Díaz-Fleischer, F.; Birke, A.; Rull, J.; Niogret, J.; Epsky, N. Susceptibility of 15 mango (Sapindales: Anacardiaceae) cultivars to the attack by Anastrepha ludens and Anastrepha obliqua (Diptera: Tephritidae) and the role of underdeveloped fruit as pest reservoirs: Management implications. J. Econ. Entomol. 2014, 107, 375–388. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Baedhowie, M.; Pranggonawati, S. Agricultural Product Quality Control Practice Guidelines; Department of Education and Culture: Jakarta, Indonesia, 1983; p. 129.
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Jallow, M.F.; Zalucki, M.P. Relationship between oviposition preference and offspring performance in Australian Helicoverpa armigera (Hübner)(Lepidoptera: Noctuidae). Aust. J. Entomol. 2003, 42, 343–348. [Google Scholar] [CrossRef]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef]
- Nufio, C.R.; Papaj, D.R. Superparasitism of larval hosts by the walnut fly, Rhagoletis juglandis, and its implications for female and offspring performance. Oecologia 2004, 141, 460–467. [Google Scholar] [CrossRef]
- Balagawi, S.; Vijaysegaran, S.; Drew, R.A.; Raghu, S. Influence of fruit traits on oviposition preference and offspring performance of Bactrocera tryoni (Froggatt)(Diptera: Tephritidae) on three tomato (Lycopersicon lycopersicum) cultivars. Aust. J. Entomol. 2005, 44, 97–103. [Google Scholar] [CrossRef]
- Rattanapun, W.; Amornsak, W.; Clarke, A.R. Bactrocera dorsalis preference for and performance on two mango varieties at three stages of ripeness. Entomol. Exp. Appl. 2009, 131, 243–253. [Google Scholar] [CrossRef]
- Muthuthantri, S.; Clarke, A.R. Five commercial citrus rate poorly as hosts of the polyphagous fruit fly Bactrocera tryoni (Froggatt) (Diptera: Tephritidae) in laboratory studies. Aust. J. Entomol. 2012, 51, 289–298. [Google Scholar] [CrossRef]
- Nishida, R. Chemical ecology of insect–plant interactions: Ecological significance of plant secondary metabolites. Biosci. Biotechnol. Biochem. 2014, 78, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Prokopy, R.J.; Vargas, R.I. Attraction of Ceratitis capitata (Diptera: Tephritidae) flies to odor of coffee fruit. J. Chem. Ecol. 1996, 22, 807–820. [Google Scholar] [CrossRef] [PubMed]
- Syamsudin, T.S. Dampak konsumsi metil egenol terhadap perilaku dan keberhasilan perkawinan lalat buah Bactrocera carambolae (Diptera: Tephritidae). Indones. J. Plant Prot. 1999, 5, 114–119. [Google Scholar]
- De Bruyne, M.; Baker, T.C. Odor detection in insects: Volatile codes. J. Chem. Ecol. 2008, 34, 882–897. [Google Scholar] [CrossRef] [PubMed]
- Brevault, T.; Quilici, S. Flower and fruit volatiles assist host-plant location in the tomato fruit fly Neoceratitis cyanescens. Physiol. Entomol. 2010, 35, 9–18. [Google Scholar] [CrossRef]
- Rodríguez-Burruezo, A.N.; Kollmannsberger, H.; González-Mas, M.C.; Nitz, S.; Fernando, N. HS-SPME comparative analysis of genotypic diversity in the volatile fraction and aroma-contributing compounds of Capsicum fruits from the annuum-chinense-frutescens complex. J. Agric. Food Chem. 2010, 58, 4388–4400. [Google Scholar] [CrossRef]
- Pino, J.; Fuentes, V.; Barrios, O. Volatile constituents of Cachucha peppers (Capsicum chinense Jacq.) grown in Cuba. Food Chem. 2011, 125, 860–864. [Google Scholar] [CrossRef]
- Cornelius, M.L.; Duan, J.J.; Messing, R.H. Volatile host fruit odors as attractants for the oriental fruit fly (Diptera: Tephritidae). J. Econ. Entomol. 2000, 93, 93–100. [Google Scholar] [CrossRef]
- Dudareva, N.; Martin, D.; Kish, C.M.; Kolosova, N.; Gorenstein, N.; Fäldt, J.; Miller, B.; Bohlmann, J.R. (E)-β-Ocimene and myrcene synthase genes of floral scent biosynthesis in snapdragon: Function and expression of three terpene synthase genes of a new terpene synthase subfamily. Plant Cell 2003, 15, 1227–1241. [Google Scholar] [CrossRef] [PubMed]
- Aluja, M.; Mangan, R.L. Fruit fly (Diptera: Tephritidae) host status determination: Critical conceptual, methodological, and regulatory considerations. Annu. Rev. Entomol. 2008, 53, 473–502. [Google Scholar] [CrossRef]
- Fletcher, B. The biology of dacine fruit flies. Annu. Rev. Entomol. 1987, 32, 115–144. [Google Scholar] [CrossRef]
- Farré-Armengol, G.; Filella, I.; Llusià, J.; Peñuelas, J. β-Ocimene, a key floral and foliar volatile involved in multiple interactions between plants and other organisms. Molecules 2017, 22, 1148. [Google Scholar] [CrossRef] [PubMed]
- Hafsi, A.; Facon, B.; Ravigné, V.; Chiroleu, F.; Quilici, S.; Chermiti, B.; Duyck, P.-F. Host plant range of a fruit fly community (Diptera: Tephritidae): Does fruit composition influence larval performance? BMC Ecol. 2016, 16, 40. [Google Scholar] [CrossRef]
- Allwood, A.; Chinajariyawong, A.; Kritsaneepaiboon, S.; Drew, R.; Hamacek, E.; Hancock, D.; Hengsawad, C.; Jipanin, J.; Jirasurat, M.; Krong, C.K. Host plant records for fruit flies (Diptera: Tephritidae) in Southeast Asia. Raffles Bull. Zool. 1999, 47, 1–92. [Google Scholar]
- Fontellas, T.M.d.L.; Zucoloto, F.S. Nutritive value of diets with different carbohydrates for adult Anastrepha obliqua (Macquart) (Diptera, Tephritidae). Rev. Bras. Zool. 1999, 16, 1135–1147. [Google Scholar] [CrossRef]
- Fernandes-da-Silva, P.G.; Zucoloto, F.S. The influence of host nutritive value on the performance and food selection in Ceratitis capitata (Diptera, Tephritidae). J. Insect Physiol. 1993, 39, 883–887. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | RT 1 (min.) | Volatile Compounds | Q 2 (%) | Relative Area (%) | |||||
|---|---|---|---|---|---|---|---|---|---|
| RK-1 | RK-2 | RK-3 | RK-4 | RK-5 | RK-6 | ||||
| 1 | 4.36 | Furan, 2-methyl- | 91 | 0 | 0 | 0 | 10.52 | 12.43 | 0 |
| 2 | 4.6 | 1-Hexyne, 5-methyl- | 38 | 11.7 | 0 | 0 | 0 | 0 | 0 |
| 3 | 5.87 | Silanediol, dimethyl | 2 | 5.21 | 0 | 0 | 17.26 | 0 | 0 |
| 4 | 6.03 | Acetamide, 2-fluoro | 3 | 10.26 | 3.91 | 0 | 0 | 0 | 0 |
| 5 | 6.19 | Formic acid, 2-methyl butyl ester | 35 | 0 | 0 | 0 | 0 | 0 | 28.4 |
| 6 | 9.52 | Furan, 3-butyltetrahydro-2-methyl-, trans- | 9 | 0 | 0 | 0 | 0 | 0 | 5.36 |
| 7 | 11.41 | 1,9-Decadiyne | 27 | 0 | 0 | 3.33 | 0 | 0 | 0 |
| 8 | 11.84 | trans-3-Decene | 37 | 0 | 0 | 0 | 0 | 0 | 3.2 |
| 9 | 12.19 | 4-Octene, 2,6-dimethyl-, [S-(E)]- | 64 | 0 | 0 | 0 | 0 | 0 | 2.75 |
| 10 | 13.14 | cis-2,6-Dimethyl-2,6-octadiene | 80 | 0 | 0 | 0 | 0 | 5.19 | 0 |
| 11 | 14.5 | β-cis-Ocimene | 91 | 0 | 0 | 0 | 0 | 17.81 | 0 |
| 12 | 21.74 | 1,4-Benzenedi-methanethiol, 2TBDMS derivates | 36 | 0 | 2.36 | 0 | 0 | 0 | 0 |
| 13 | 42.25 | 2-Tetradecanol | 35 | 0 | 0 | 0 | 2.78 | 0 | 0 |
| 14 | 43.1 | 1-Octadecene | 35 | 7.23 | 0 | 0 | 0 | 0 | 0 |
| 15 | 43.61 | Butyl dodecyl ether | 35 | 7.51 | 0 | 0 | 4.07 | 0 | 0 |
| 16 | 44.26 | Isobutyl nonyl carbonate | 50 | 0 | 0 | 0 | 2.25 | 0 | 0 |
| 17 | 46.38 | 1-Decanol, 2-hexyl | 22 | 12.67 | 0 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Syamsudin, T.S.; Kirana, R.; Karjadi, A.K.; Faizal, A. Characteristics of Chili (Capsicum annuum L.) That Are Resistant and Susceptible to Oriental Fruit Fly (Bactrocera dorsalis Hendel) Infestation. Horticulturae 2022, 8, 314. https://doi.org/10.3390/horticulturae8040314
Syamsudin TS, Kirana R, Karjadi AK, Faizal A. Characteristics of Chili (Capsicum annuum L.) That Are Resistant and Susceptible to Oriental Fruit Fly (Bactrocera dorsalis Hendel) Infestation. Horticulturae. 2022; 8(4):314. https://doi.org/10.3390/horticulturae8040314
Chicago/Turabian StyleSyamsudin, Tati Suryati, Rinda Kirana, Asih Kartasih Karjadi, and Ahmad Faizal. 2022. "Characteristics of Chili (Capsicum annuum L.) That Are Resistant and Susceptible to Oriental Fruit Fly (Bactrocera dorsalis Hendel) Infestation" Horticulturae 8, no. 4: 314. https://doi.org/10.3390/horticulturae8040314
APA StyleSyamsudin, T. S., Kirana, R., Karjadi, A. K., & Faizal, A. (2022). Characteristics of Chili (Capsicum annuum L.) That Are Resistant and Susceptible to Oriental Fruit Fly (Bactrocera dorsalis Hendel) Infestation. Horticulturae, 8(4), 314. https://doi.org/10.3390/horticulturae8040314