Abstract
Chinese yam, with its rich taste and high nutritive value, has remained an essential part of Chinese cuisine. As China is rich in yam germplasm resources, understanding the genetic diversity of local varieties, which is important for the proper management, conservation, variety improvement, and sustainable utilization of yam germplasms, is crucial. In this study, an Illumina Hi-seq 2500 sequencing (Illumina, San Diego, CA, USA) system was used to sequence cDNA libraries of root, stem, and leaf tissues of Chinese yam; 14,450 unigenes containing SSR sites were identified from the transcriptome dataset. About half of the SSRs were mononucleotides and approximately 23% were dinucleotides and trinucleotides. In addition, 100 SSR primers were randomly designed for PCR and polymorphism verification. A total of 32 yam materials used in the study were identified by 24 primer pairs. Based on the Qinling-Huaihe River line, the northern and southern germplasms were divided into different taxa, but no clear geographical pattern was observed in the southern germplasms. The 32 germplasms were also divided into three categories using a clustering analysis, with samples in the same group possessing similar morphological characters. These 24 newly identified SSR markers can provide the basis for the genetic identification, diversity analysis, and marker-assisted selection breeding of Dioscoreaceae.
1. Introduction
Yam (Dioscorea spp.), belonging to the Dioscoreaceae family of the order Dioscoreales, consists of about 600 domesticated and wild species [1]. It is one of the world’s most important root crops, with its production rate only behind that of potato, sweet potato, and cassava [2]. Yams are a major staple food and an important source of carbohydrates for people in Africa, Central and South America, parts of Asia, the Caribbean, and the Pacific Islands [3,4,5]. However, only about 10 edible species are widely grown for food. China, which has been domesticating yams since the Song Dynasty and cultivating them for over 1000 years, is an important producer of yams. In China, yam is extremely abundant, with 65 different species present across the country [6]. The different species of yam are mainly divided into four categories: Dioscorea, Dioscorea alata, Dioscorea fordii, Dioscorea persimilis [7]. Yam is cultivated in all provinces except Qinghai and Tibet and includes a large number of landraces. Yam tubers are rich in nutritional elements, such as polysaccharides, proteins, vitamins, and mineral elements, and medicinal health components, like mucilage, saponins, allantoin, and choline [8,9,10,11]. They improve human digestive function and immunity, reduce levels of blood sugar and blood lipids, and promote kidney repair and regeneration. The tuber is juicy and soft and is highly edible with a high medicinal value. In 2002, it became one of the first Chinese herbal medicines to be labeled as a medicine food homology plant by the National Health Commission [12]. Yam is an important component of a Chinese herbal detoxifying soup where it has been shown to help in lung-clearing and moistening of the spleen. This soup has an overall efficiency of more than 90% in the treatment of COVID-19 [13].
Chinese yam is a high-yield crop that shows strong adaptability; the tubers are resistant to microbial rot during storage. The cultivation of yam is widespread and the continuous expansion of Chinese yam planting areas has resulted in an increasing demand for improved varieties [14]. However, most research on Chinese yam is focused on medicinal ingredients or food processing; fewer studies have covered the molecular analysis of Chinese yam. Due to historical changes and long-term mutual introgression, as well as the vulnerability of yams to their surroundings, some germplasm resources have similar external morphological characteristics. This makes them difficult to identify quickly as the same species or the same taxa [15], which seriously hinders the selection and promotion of yam varieties. Given that differences between breeding parents and germplasm resources can affect target traits, genetic diversity evaluation and research are of great importance in yam breeding [16].
The genetic diversity of a species is reflected in its morphological, biochemical, cellular, and molecular differences, which are essentially linked to variations in genetic material. DNA molecular markers directly reflect heredity and variation in a species. Because of their high stability, low cost, and simple operation, molecular markers are important tools in the construction of genetic linkage maps, molecular marker-assisted breeding, quantitative trait mining, phylogeny, and genetic diversity. Microsatellite sequences, such as simple sequence repeats (SSR), are widely distributed in the plant genome. SSRs offer unique advantages in genetic studies as they are attributed to co-dominant inheritance, high polymorphism, good repeatability. SSRs are widely used in genetic research related to the breeding of crops such as Chinese cabbage [17], pepper [18], and tea tree [19].
In this study, we used high-throughput sequencing to analyze and screen the distribution characteristics of SSR markers in transcriptome sequences of Chinese yam. And next included the design of SSR primers, and further validation of SSRs by evaluating the genetic diversity and identifying the 32 germplasms used in the study. Our study would form a basis for the identification, genetic diversity analysis, gene mapping, core germplasm building, and molecular marker-assisted breeding of Chinese yam.
2. Materials and Methods
2.1. Plant Materials and DNA Extraction
For transcriptome sequencing, GN_ZY_huaishan was selected from the yam germplasm resource nursery in Yichun City, Jiangxi Province, China. This yam variety has good traits, and the tubers are rich in anthocyanins. The roots, stems, and leaves of the plant were used for the experimental material. The tissue materials grew normally without any diseases, insect pests, or mechanical damage. Three biological replicates were maintained for each tissue. These samples were flash-frozen in liquid nitrogen and stored in a −80 °C freezer at the Institute of Bast Fiber Crops, Chinese Academy of Agricultural Sciences in Changsha for subsequent RNA extraction and transcriptome sequencing.
For SSR screening and germplasm genetic diversity evaluation, 32 yam germplasm tubers were selected as experimental materials. The yams were collected from 10 provinces across China and had different morphological characteristics (Table 1). DNA extraction of the 32 yam tubers was performed using a Plant Genome DNA Extraction Kit (Tiangen Biochemical Technology, Beijing, China) following the manufacturer’s instructions. The concentration and purity of the total genomic DNA were measured using a NanoDrop 2000 Ultra-Micro Spectrophotometer (Thermo, Dalaware, MA, USA). The qualified genomic DNA stock solutions were diluted to 20 ng/μL and stored in a −20 °C freezer until further analysis.

Table 1.
List of experimental material.
2.2. RNA Extraction and cDNA Library Preparation
Roots, stems, and leaves of GN_ZY_huaishan were used as materials for RNA extraction, which was conducted using a mirVana™ miRNA ISOlation Kit (Mito-Bio, Shanghai, China) digested with DNase. We prepared a cDNA library as described by Christopher [20], which we first enriched with eukaryotic mRNA using magnetic beads with Oligo (dT); next, mRNA was broken into short fragments by adding an interruption reagent, and the interrupted mRNA was used as a template to synthesize one-stranded cDNA with six-base random primers. The purified double-stranded cDNA was then end-repaired, A-tailed, and sequenced, followed by fragment size selection, and finally PCR amplification, resulting in the construction of a cDNA library.
2.3. De Novo Transcriptome Assembly
Transcriptome sequencing without reference was commissioned by Shanghai Ouyi Biomedical Technology (Shanghai, China) and was conducted using the Illumina HiSeq 2500 sequencing platform. Before assembly, the raw sequencing data were filtered by removing the adapter, N bases accounting for more than 10%, and the number of bases with a mass value of ≤15 to obtain clean reads [19]. The transcript sequences were obtained using the paired-end splicing method of the short sequence assembly software, Trinity [21]. The longest reads were selected as unigenes based on the similarity and length of the sequences. Clustering and removal of redundancy was performed using CD-HIT [22] to obtain the final unigene sequence. Unigenes were blasted with NR, KOG, GO, Swiss-Prot, eggNOG, and KEGG databases by using DIAMOND, and Pfam databases were blasted using HMMER [23] to analyze the functionality of unigenes.
2.4. Development and Primer Design of SSR Markers
SSRs were identified using the microsatellite identification tool, MicroSatellite (MISA; accessed on 7 February 2022, http://pgrc.ipk-gatersleben.de/misa/misa.html) [24], and through a manual search of the unigene sequences. The following selection criteria were used for SSRs: the number of nucleotide and dinucleotide motifs were not less than 15 and 8, respectively; the number of trinucleotide, tetranucleotide, pentanucleotide, and hexanucleotide motifs exceeded five. Sequences with SSR site flanking lengths ≥50 bp were selected and primers were designed using Primer Premier 5.0. To obtain better amplification, the Musa Kavas’s parameters [25] were used: the PCR product length was 100–300 bp, the primer length was 18–23 bp, the GC content was 40–60%, the annealing temperature was 52–60 °C, and the difference in annealing temperatures between the forward and reverse primers was <2 °C. The designed primers were then subjected to BLAST analysis in the NCBI database for target specificity. A total of 100 primer pairs were designed, including five pairs of single nucleotides, thirty pairs of dinucleotides, thirty pairs of trinucleotides, twenty pairs of tetranucleotides, ten pairs of pentanucleotides, and five pairs of hexanucleotides, and synthesized by Tsingke Biotechnology Corporation (Beijing, China).
2.5. PCR Amplification and Polymorphism Analysis
To analyze the genetic diversity of Chinese yam, PCR was performed (Bio-Rad T100™ Thermal Cycler, Bio-Rad Laboratories, Hercules, CA, USA) with SSR primers and DNA from 32 yam germplasms as the template. PCR reactions and electrophoresis were conducted as outlined by Niu [26], and the PCR system was designed as follows: 1 μL of 1 × Taq buffer, 0.5 μL of 0.2 mmol/L dNTP Mix, 0.15 μL of 1U Taq enzyme, 0.5 μL of 0.25 μmol/L of each primer, 1.5 μL of 20 ng DNA, 6 μL of nuclease-free water. The PCR cycle conditions used were as follows: initial denaturation at 94 °C for 5 min, 34 cycles of 94 °C for 30 s, 55 °C for 30 s, 72 °C for 1 min, and final extension at 72 °C for 10 min. PCR products were detected using an 8.0% non-denaturing polyacrylamide gel (PAGE) electrophoresis, which was run at 105 V for 3 h. DL100 DNA marker (Beijing Tiangen Biotechnology, Beijing, China) was used as the standard size for the PCR products. The PAGE gel was then stained using 0.1% AgNO3 solution; a combination of 1.5% NaOH and formaldehyde was used as the developer solution.
2.6. Data Analysis
The SSR results were evaluated as described by Sabreena [27], where the polymorphic bands produced by the different primers that were present in a sample were recorded as “ones” under that polymorphic locus and those not present were recorded as “zeros”, thereby constructing a matrix of zeros and ones. The results were tabulated using Microsoft Excel 2013. In addition, SSR marker characteristics, such as major allele frequency, genotype number, and allele number, were analyzed using PowerMarker v3.25 [28]; clustering was performed using ITOL (accessed on 11 June 2022, https://itol.embl.de/); Structure2.3.4 and Structure Harvester (accessed on 14 June 2022, http://taylor0.biology.ucla.edu/structureHarvester/) were used to analyze population genetic structures [29].
3. Results
3.1. De Novo Transcriptome Assembly
Transcriptome sequencing of root, stem, and leaf tissues of GN_ZY_huaishan resulted in a total of 19.32 G of clean data; the amount of clean data for the roots, stems, and leaves were 6.35, 6.68, and 6.29 G, respectively, with a Q30 base distribution of 94.67–95.44% and a GC content of 45.76–46.27%. All indices met the requirements of transcriptome sequencing. The clean reads were spliced to obtain a total of 56,195 unigenes with a total length of 56,209,464 bp and an average length of 1000.26 bp (Table 2). Most of the unigenes were concentrated in the 300–500 bp range, which accounted for 51.37% of the total number of unigenes; unigenes in the 500–2000 bp range accounted for 35.78%, and those >2000 bp accounted for 12.85% (Figure 1).
Table 2.
RNA-sequencing data of Dioscorea spp.
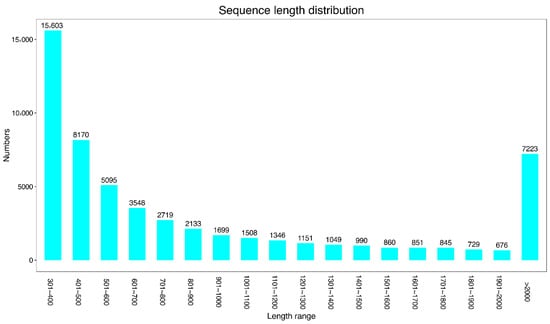
Figure 1.
Unigenes from the de novo transcriptome assembly of Chinese yam. The bars represent different sizes and number of unigenes obtained using transcriptome analysis of leaf, stem, and root tissues of Chinese yam.
3.2. UniGene Function
The unigene sequence was blasted with Pfam, NR, KOG, eggNOG, Swiss prot, GO, and KEGG databases using DIAMOND [30]; e < 1e−5 annotations were used to screen for proteins with the highest sequence similarity; 22,045 (39.23%) unigenes were annotated in the Pfam database, 34,938 (62.17%) were annotated in the NR database, 33,336 (59.32%) were annotated in the eggNOG database, 24,861 (44.24%) were annotated in the Swiss-Prot database, 22,577 (40.18%) were annotated in the GO database, 21,841 (38.87%) were annotated in the KOG database, and 7802 (13.88%) were annotated in the KEGG database. There were 4678 unigenes that were annotated in all seven databases (Figure 2).
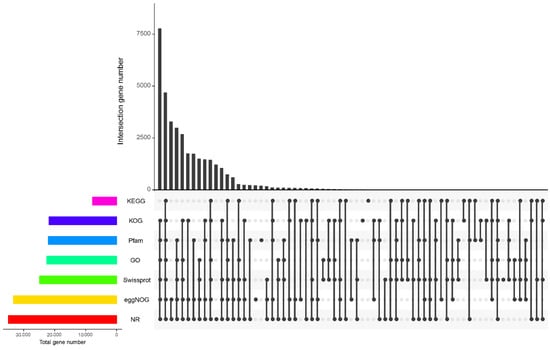
Figure 2.
Each database annotates a Venn diagram.
3.3. Microsatellite Feature Analysis
A SSR search was carried out on the 56,195 unigenes of the Chinese yam transcriptome using MISA. SSR sites were detected in 14,450 unigenes, but the number of different repeat sequences varied greatly, and the distribution was not particularly balanced. Single nucleotide repeats constituted the most common form, with 10,389 SSRs accounting for 50.73% of the total SSRs. This was followed by 4751 dinucleotide and 4791 trinucleotide SSRs, accounting for 23.2% and 23.39% of the total SSRs, respectively. Tetranucleotides (399), pentanucleotides (68), and hexanucleotides (83) constituted a small proportion, accounting for only 1.95%, 0.33%, and 0.41% of the total number of SSRs, respectively, (Figure 3).
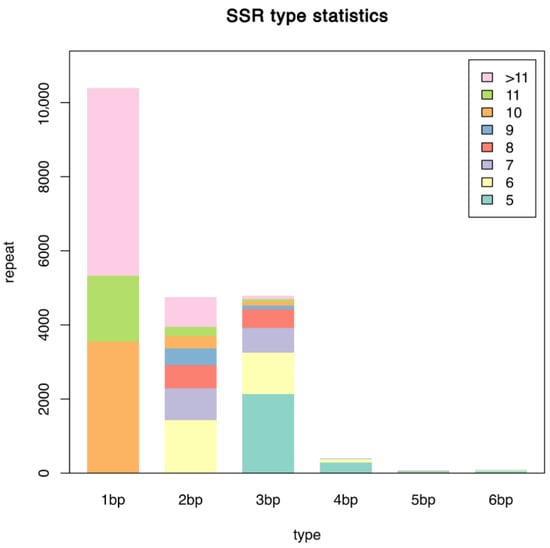
Figure 3.
SSR type statistics chart.
3.4. Genetic Diversity of Chinese Yam Germplasm
To determine the genetic diversity of the Chinese yam germplasm, we randomly designed 100 pairs of SSR primers from the SSR sites detected using transcriptome screening. The results showed that 24 primer pairs amplified SSR products in all 33 samples; 76 primer pairs amplified products that were not clear or polymorphic. A total of 58 polymorphic sites were detected, and the number of primer elements was between two and four, with an average number of primer elements of 2.42. The characteristics of the SSR primers that detected polymorphism are indicated in Table 3. Major.Allele.Frq ranges from 0.47 to 0.96, with a mean of 0.69; GenotypeNo ranges from 2.00 to 8.00, with an average of 3.29; AlleleNo ranges from 2.00 to 5.00, with an average of 2.54; Availability ranges from 0.31 to 0.97, with an average of 0.76; Heterozygosity ranges from 0.00 to 0.56, with an average of 0.21; and PIC amplitude ranges from 0.07 to 0.58, with an average of 0.36 (Table 3).
Table 3.
Characteristics of 24 SSR primers that amplified successfully and exhibited polymorphism in Chinese yam.
The clustering tree constructed using PowerMarker showed that the 24 SSR markers divided the 32 germplasms into three groups (Figure 4). The first group mainly included Dioscorea and Dioscorea alata and consisted of the Ming Huai II, GN_ZY_huaishan, YF_huaishan, HN_yam, YN_XJ_yam germplasms which originate from Fujian Province, Hainan Province, Yunnan Province, and other southern provinces and cities in China, and mainly have a long columnar tuber shape. The second group, which is characterized by germplasms with a long columnar shape, mainly included Dioscorea persimilis, with two-thirds of the germplasms distributed north of the Qinling-Huaihe line. The third group contained many Dioscorea alata germplasms, most of which, with the exception of SDHZ_TG_yam, XS_yam, and Bikeqichang yam, are distributed south of the Qinling Huaihe line and are cylindrical and flat block-shaped. Genetic structure analysis was performed on the 32 germplasms using STRUCTURE 2.3.4 software. These germplasms were completely distinguished at K = 3, and were divided into different groups by the Qinling-Huaihe line (Figure 5). The results of the cluster analysis and genetic structure analysis showed that the genetic background of the germplasms was correlated with their source, altitude, type, and tuber shape.
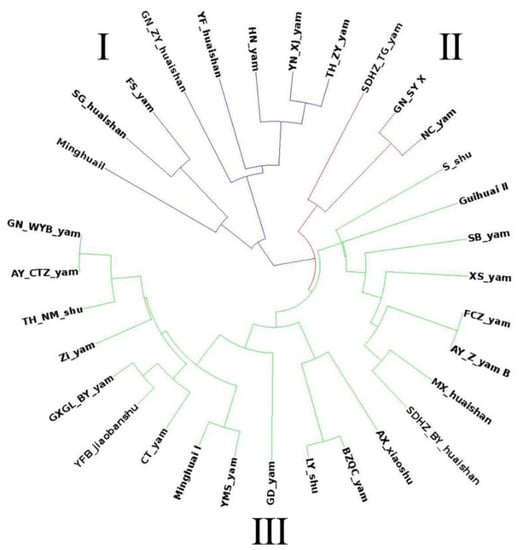
Figure 4.
Phylogenetic tree of 32 Chinese yam germplasms. purple lines: Clade I; red lines: Clade II; green lines: Clade III.
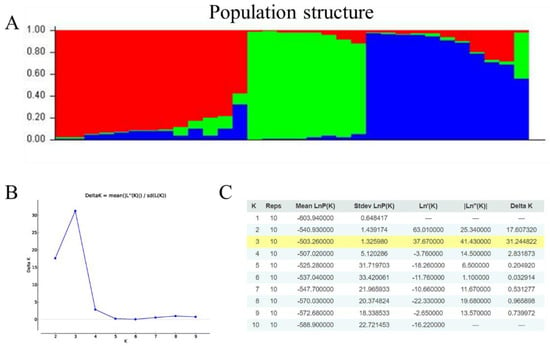
Figure 5.
Population structure analysis of 32 Chinese yam accessions. (A) Population structure based on K = 3. When K = 3, red: Group 1; green: Group 2; blue: Group 3. (B) Delta K based on the rate of change of L (K) between successive K values. (C) Evanno table output.
4. Discussion
Yam is a highly productive and efficient cash crop for food and medicine. China has a wide variety of yam species and a large cultivation area, but there are problems such as confusing names, degraded varieties, poor quality, and low yields [31]. Meanwhile, research on the genetic background, germplasm creation, hybrid embryo culture, and tissue propagation techniques of yam in China is not comprehensive enough. For this reason, this study collected 32 germplasm resources of Dioscorea, Dioscorea alata, Dioscorea persimilis with different shapes and colors from different cities and counties in 10 provinces across China. SSR molecular markers were developed by transcriptome sequencing of different tissues of GN_ZY_huaishan, and genetic distance trees were constructed to understand their genetic background at the molecular level and to identify and classify their germplasm more accurately. This information can serve as practical guidance for germplasm innovation, variety improvement, resource conservation, and yam utilization.
4.1. Transcriptome Sequencing, Assembly, and Functional Annotation
With the development of high-throughput sequencing technology, Illumina RNA-Seq is recognized as a highly efficient, relatively fast, and inexpensive high-throughput method for obtaining the full transcript sequence of different species and has been successfully applied to the research of gene mining and gene expression [32,33,34]. GN_ZY_huaishan is a cultivar from Guanan County, Lianyungang City, Jiangsu Province, and belongs to Dioscorea. Unlike the common yellow-skinned and white-fleshed yam on the market, it has purple skin and purple flesh, is rich in anthocyanins, has health benefits, and has great potential for development. In this study, the transcriptome of GN_ZY_huaishan was sequenced by simultaneously growing root, stem, and leaf tissues in the Yichun Institute of Science Germplasm Resource Nursery, resulting in the production of 6.35 G, 6.68 G, 6.29 G of clean data, respectively, with marginal differences among them.
Using DIAMOND and HAMMER, 4678 unigenes were blasted, revealing functional annotations in all seven public databases. These results indicate that the sequencing data comprised a wide diversity of transcripts, where unigenes that were not blasted may locate 3′ or 5′ UTR of the gene, non-coding RNA, or short sequences without known protein domains. Thus, many new unigenes may represent potential yam-specific genes. Detecting these genes would increase our understanding of the processes that regulate yam growth, development, and secondary metabolite production and would help build high-density microarrays to further characterize relevant gene expression profiles.
4.2. SSR Markers
Knowledge of the genetic background of germplasm is important for the conservation, management, and improvement of crops [35]. The large amount of genetic variation distributed in yam populations, which controls traits such as flowering (FL), leaf shape (LS), leaf color (LC), stem color (SC), twining direction (TD), tuber shape (TS), root hair density (RHD), tuber skin color (TSC), flesh color (FC), and tuber flesh weight (TFW) [36], means that analysis of genetic structure and diversity is essential for understanding the development of yam. Some researchers have dissected the genetic diversity of yam using phenotypic, molecular, or combinatorial analysis [36,37,38]. DNA markers in genetic characterization studies can help researchers understand the structure and extent of variations within plant species, as the genome covers a wide range and is more consistent. As one of the most widely used molecular markers in crops and fruit trees, RNA-Seq has become the preferred method for building gene maps and quantifying transcriptomes, especially in non-model organisms such as yam.
A total of 14,450 SSRs were identified in the yam transcriptome dataset; unlike other plants, we obtained an extremely high rate of single nucleotide SSRs, which accounted for 50.73% of the total SSRs. Excluding single nucleotide repeat types, approximately 12.67% of unigenes contained polynucleotide SSRs, which was higher than the proportion obtained for pepper (4.9%) [18], sweet potato (8.2%) [39], flax (3.5%) [40], peanut (6.8%) [41], and sesame (8.9%) [42], but lower than that obtained for radish (23.8%) [43], coffee (18.5%) [44], and castor bean (28.4%) [45]. Expressed sequence tags-SSR (EST-SSR) detection depends on many factors, such as genomic structure, tools, and the method used for EST-SSR exploration [46]. In the sequencing results of yam, dinucleotides and trinucleotides constituted the most common motifs, accounting for 47.09% and 47.47% of the total polynucleotides. This was different from the sequencing results of sweet potato [47] and sesame [42], where dinucleotides accounted for the greatest number of motifs.
In this study, we collected 32 germplasm resources of Dioscorea, Dioscorea alata, Dioscorea persimilis from different cities and counties in 10 provinces. Among the resources, tuber shapes included flat block shape, long column shape and cylinder shape, the epidermal color of the tuber included purple and yellow, and the flesh color of the tuber included white and purple. Genetic diversity and genetic structure analysis of the 32 resources was determined using SSRs found by transcriptome sequencing of different tissues of yam using 100 pairs of randomly selected and designed primers. The analysis showed that the germplasm resources were clustered into three categories based on geographic origin, with the Qinling-Huaihe line as the dividing line, and that resources from the north and south were clustered into one category, similar to the results of Zhang et al. [17]. In addition, germplasm from the same province did not always cluster in the same taxon; YF_huaishan, TH_ZY_yam, TH_NM_shu, and AY_CTZ_yam from Jiangxi were not classified in the same taxon, indicating that factors other than geographical location, such as variety, altitude, climate, biological characteristics, and genetic drift, may also influence germplasm genetic diversity [48]. Regarding yam characteristics, the first group mainly included Dioscorea alata and Dioscorea, and included varieties with a long columnar tuber shape, the second group mainly consisted of Dioscorea persimilis, with all varieties having a long columnar tuber shape, and the third group mainly included Dioscorea alata and consisted of varieties with flat and cylindrical tuber shapes.
Our results indicate that Chinese yam is highly diverse, showing polymorphism at the molecular level. However, because the germplasms used in this study were few, this may have led to deviations in genetic diversity analysis due to the homonymy phenomenon. Huang Yuxian [49] used the SPARP molecular marker technology to divide 94 yam germplasm resources into four categories: Dioscorea, Dioscorea alata, Dioscorea fordii, and Dioscorea persimilis; Hua Shumei [50] used RAPD marker cluster analysis to divide 34 yam resources into four groups: ordinary yam, field yam, flat yam, and Fujian big yam. In comparison to the above studies, the taxa used in this study were relatively simple, which may be related to the number of tested materials and the limitations discussed in the introduction.
5. Conclusions
Yam germplasm, which is widely used in cultivation, exhibits high intraspecific and interspecific diversity in China. Phenotypic and molecular markers are very effective tools for detecting yam diversity. A comprehensive analysis of yam transcriptome data using the Illumina sequencing platform identified 14,450 SSR markers. To further analyze the application potential and the relationship between genetic diversity and phylogeny of yam germplasm resources, 32 yam germplasm resources were identified with 24 polymorphic primers. To identify related varieties, these germplasms were classified according to phenotypic characteristics and geography, which is important for the conservation of yam germplasm resources and promotion of yam industrialization. The results of this study not only lay a good foundation for the study of yam genetic diversity, but also provide a theoretical foundation for the application of fingerprinting, genetic diversity analysis, marker trait association studies, and QTL localization markers in future research.
Author Contributions
H.W. and Y.W. contributed equally to this work. M.L. and X.C. jointly supervised this study. H.W., Y.W., L.X. and J.S. performed transcriptome and gene functional analyses. H.W., Y.W., Y.C., C.O., B.L. and H.Z. prepared the DNA and RNA samples and performed the PCR analysis. Y.W. conceived the study and drafted the manuscript. H.W. and Y.W. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The materials of this study were provided by the Yichun Institute of Science. Correspondence and requests for materials should be addressed to Xiaorong Chen (chenxiaorong1977@126.com).
Acknowledgments
This work was financially supported by the Yichun Innovation Drive 5511 Engineering Science and Technology Project (AZA-15-03), the Agricultural Science and Technology Innovation Program (ASTIP) of CAAS (Grant No. 2017IBFC) and Jiangxi Province Vegetable Industry Major Technology Collaborative Promotion Program (Ganong Technology [2022] No. 3).
Conflicts of Interest
The authors declare no competing financial interest.
References
- Azeteh, I.N.; Hanna, R.; Njukeng, A.P.; Oresanya, A.O.; Sakwe, P.N.; Kumar, P.L. Distribution and Diversity of Viruses Infecting Yams (Dioscorea spp.) in Cameroon. VirusDisease 2019, 30, 526–537. [Google Scholar] [CrossRef]
- Luo, G.F.; Podolyan, A.; Kidanemariam, D.B.; Pilotti, C.; Houliston, G.; Sukal, A.C. A Review of Viruses Infecting Yam (Dioscorea spp.). Viruses 2022, 14, 662. [Google Scholar] [CrossRef] [PubMed]
- Adenji, M.O.; Shoyinka, S.A.; Ikotun, T.; Asiedu, R.; Hughes, J.d.A.; Odu, B.O. Yield Loss in Guinea Yam (Dioscorea rotundata Poir.) Due to Infection by Yam Mosaic(Ymv) Genus Potyvirus. Ife J. Sci. 2012, 14, 237–244. [Google Scholar]
- Amusa, N.; Adegbite, A.A.; Muhammed, S.; Baiyewu, R.A. Yam Diseases and Its Management in Nigeria. Afr. J. Biotechnol. 2003, 2, 497–502. [Google Scholar] [CrossRef]
- Tamiru, M.; Natsume, S.; Takagi, H.; White, B.; Yaegashi, H.; Shimizu, M.; Yoshida, K.; Uemura, A.; Oikawa, K.; Abe, A.; et al. Genome Sequencing of the Staple Food Crop White Guinea Yam Enables the Development of a Molecular Marker for Sex Determination. BMC Biol. 2017, 15, 86. [Google Scholar] [CrossRef]
- Guo, S.L.; Liu, P. A Study on the Floristic Geographical Elements of the Chinese Dioscorea. J. Zhejiang Norm. Univ. 1994, 17, 81–85. [Google Scholar]
- Lei, F.G.; Hua, S.M.; Tu, Q.C.; He, P.Z.; Li, Q.X.; Zao, Y.Y. Genetic Relationship of Dioscorea polystachya Turcz. Resources Based on ISSR Markers. Fujian J. Agric. Sci. 2013, 28, 27–32. [Google Scholar] [CrossRef]
- Zhang, M.; Lin, D.; Huang, G.D.; Zhong, X.F. Study on Extraction of the Total Saponins of Chinese Yam. J. Chin. Med. Mater. 2007, 7, 854–857. [Google Scholar]
- Chen, M.Y.; Liu, W.; Chou, G.X.; Wang, Y.L. Research Progress on Chemical Constituents and Pharmacological Activities of Dioscorea opposita Thunb. Acta Chin. Med. Pharm. 2020, 2, 5. [Google Scholar] [CrossRef]
- Fang, X.Y.; Hou, Y.J.; Jia, S.Y.; Si, R.H.; Zheng, B.W.; Liu, G.Z. Research Progress on Chemical Constituents of Dioscoreae rhizoma and Pharmacological Effects of Saponins. Inf. Tradit. Chin. Med. 2021, 38, 79–84. [Google Scholar] [CrossRef]
- Jie, N. Study on Nutrition Content and Quality Evaluation of Different Chinese Yam (Dioscorea Opposite Thunb.) Cultivars. Ph.D. Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2010. [Google Scholar]
- Dong, M.J.; Li, J.C.; Meng, Y.J.; Yang, T.X.; Ge, S.J. Research Progress on Germplasm Resources Identification and Breeding of Chinese Yam. J. Henan Agric. Sci. 2021, 11, 6–14. [Google Scholar] [CrossRef]
- Yao, J.; Shi, X.Y.; Chen, Q.; Fang, S.M.; Yang, R.X.; Peng, B. Theoretical Study on Corona Virus Disease 2019 Treated by Qingfei Paidu Decoction. Liaoning J. Tradit. Chin. Med. 2020, 5, 94–98. [Google Scholar]
- Jing, X.; Jiang, H.; Du, H.H.; Meng, Y.P. Research Progress on Dioscorea opposita in China. J. Anhui Agric. Sci. 2016, 15, 4. [Google Scholar] [CrossRef]
- Liu, X.Y.; Huo, W.X.; Yang, M.; Wang, D.; Zhou, Y.H.; Zhao, Z.H.; Tai, L.H. Genetic Diversity Analysis and Primary Core Collection Construction in Yam (Dioscorea opposita Thunb.) by ISSR Marker. Acta Bot. Boreali-Occident. Sin. 2015, 5, 915–921. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Wang, L.J.; Lei, J.; Chai, S.S.; Yang, X.S.; Zhang, W.Y. Genetic Diversity Analysis and Construction of DNA Fingerprint of Yam (Dioscorea opposita Thunb.) Germplasm by cpSSR Marker. Acta Agric. Zhejiangensis 2021, 33, 1222–1233. [Google Scholar] [CrossRef]
- Ding, Q.; Li, J.J.; Wang, F.D.; Zhang, Y.H.; Li, H.Y.; Zhang, J.N.; Gao, J.W. Characterization and Development of EST-SSRs by Deep Transcriptome Sequencing in Chinese Cabbage (Brassica rapa L. ssp. pekinensis). Int. J. Genom. 2015, 2015, 473028. [Google Scholar] [CrossRef]
- Kenta, S.; Kohei, I.; Cholgwang, K.; Tomohiro, B.; Munenori, S.; Takashi, I.; Toshiya, M.; Megumi, K.; Noriko, N.; Sachiko, I.; et al. Development of Capsicum EST-SSR Markers for Species Identification and in Silico Mapping onto the Tomato Genome Sequence. Mol. Breed. 2013, 31, 101–110. [Google Scholar] [CrossRef]
- Tan, L.Q.; Wang, L.Y.; Zhang, C.C.; Wei, K.; Zhang, C.C.; Wu, L.Y.; Qi, G.N.; Cheng, H.; Zhang, Q.; Cui, Q.M.; et al. Floral Transcriptome Sequencing for SSR Marker Development and Linkage Map Construction in the Tea Plant (Camellia sinensis). PLoS ONE 2013, 8, e81611. [Google Scholar] [CrossRef]
- Saski, C.A.; Bhattacharjee, R.; Scheffler, B.E.; Asiedu, R. Genomic Resources for Water Yam (Dioscorea alata L.): Analyses of EST-Sequences, De Novo Sequencing and GBS Libraries. PLoS ONE 2015, 10, e134031. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.L.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Trinity: Reconstructing a Full-Length Transcriptome without a Genome from RNA-Seq Data. Nat. Biotechnol. 2013, 29, 644–652. [Google Scholar] [CrossRef]
- Li, W.; Jaroszewski, L.; Godzik, A. Clustering of Highly Homologous Sequences to Reduce the Size of Large Protein Databases. Bioinformatics 2001, 17, 282–283. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Finn, R.D.; Eddy, S.R.; Bateman, A.; Punta, M. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucleic Acids Res. 2013, 41, e121. [Google Scholar] [CrossRef] [PubMed]
- Thiel, T.; Michalek, W.; Varshney, R.; Graner, A. Exploiting EST Databases for The Development and Characterization of Gene-Derived SSR-Markers in Barley (Hordeum vulgare L.). Theor. Appl. Genet. 2003, 106, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Kavas, M.; Yıldırım, K.; Seçgin, Z.; Gökdemir, G. Discovery of Simple Sequence Repeat (SSR) Markers in Hazelnut (Corylus avellana L.) by Transcriptome Sequencing and SSR-Based Characterization of Hazelnut Cultivars. Scand. J. For. Res. 2020, 35, 227–237. [Google Scholar] [CrossRef]
- Niu, J.; Wang, Y.J.; Shi, Y.L.; Wang, X.F.; Sun, Z.M.; Huang, K.Y.; Gong, C.; Luan, M.B.; Chen, J.H. Development of SSR Markers via De Novo Transcriptome Assembly in Akebia trifoliata (Thunb.) Koidz. Genome 2019, 62, 817–831. [Google Scholar] [CrossRef]
- Sabreena; Nazir, M.; Mahajan, R.; Hashim, M.J.; Iqbale, J.; Alyemeni, M.N.; Ganai, B.A.; Zargar, S.M. Deciphering Allelic Variability and Population Structure in Buckwheat: An Analogy between the Efficiency of ISSR and SSR markers. Saudi J. Biol. Sci. 2021, 28, 6050–6056. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. PowerMarker: An Integrated Analysis Environment for Genetic Marker Analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Yang, S.H.; Rehman, H.M.; Baloch, F.S.; Lee, J.D.; Park, J.H.; Chung, G. Genetic Diversity and Population Structure of Korean Wild Soybean (Glycine soja Sieb. and Zucc.) Inferred from Microsatellite Markers. Biochem. Syst. Ecol. 2017, 71, 87–96. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, H.D. Fast and Sensitive Protein Alignment Using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Huang, Y.X.; Wang, F.Q.; Du, J.F.; Hua, S.M.; Lei, F.G.; Xu, X.M.; Liang, K.J.; Zhang, Z.Y. Numerical Analysis of Morphological Variation of Germplasm Resources of Dioscorea. China J. Chin. Mater. Med. 2013, 38, 318–324. [Google Scholar]
- Kamunya, S.M.; Wachira, F.N.; Pathak, R.S.; Korir, R.; Sharma, V.; Kumar, R.; Bhardwaj, P.; Chalo, R.; Ahuja, P.S.; Sharma, R.K. Genomic Mapping and Testing for Quantitative Trait Loci in Tea (Camellia sinensis (L.) O. Kuntze). Tree Genet. Genomes 2010, 6, 915–929. [Google Scholar] [CrossRef]
- Hu, C.Y.; Lee, T.C.; Tsai, H.T.; Tsai, Y.Z.; Lin, S.F. Construction of an Integrated Genetic Map Based on Maternal and Paternal Lineages of Tea (Camellia sinensis). Euphytica 2013, 191, 141–152. [Google Scholar] [CrossRef]
- Wu, H.; Chen, D.; Li, J.X.; Yu, B.; Qiao, X.Y.; Huang, H.L.; He, Y.M. De Novo Characterization of Leaf Transcriptome Using 454 Sequencing and Development of EST-SSR Markers in Tea (Camellia sinensis). Plant Mol. Biol. Rep. 2013, 31, 524–538. [Google Scholar] [CrossRef]
- Darkwa, D.; Agre, A.P.; Olasanmi, B.; Iseki, K.; Matsumoto, R.; Powell, A.; Bauchet, G.; Koeyer, D.D.; Muranaka, S.; Adebola, P.; et al. Comparative Assessment of Genetic Diversity Matrices and Clustering Methods in White Guinea Yam (Dioscorea rotundata) Based on Morphological and Molecular Markers. Sci. Rep. 2020, 10, 13191. [Google Scholar] [CrossRef]
- Cao, T.; Sun, J.Y.; Shan, N.; Chen, X.; Wang, P.; Zhu, Q.; Xiao, Y.; Zhang, H.Y.; Zhou, Q.H.; Huang, Y.J. Uncovering the Genetic Diversity of Yams (Dioscorea spp.) in China by Combining Phenotypic Trait and Molecular Marker Analyses. Ecol. Evol. 2021, 11, 9970–9986. [Google Scholar] [CrossRef]
- Agre, P.A.; Dassou, A.G.; Loko, L.E.Y.; Idossou, R.; Dadonougbo, E.; Gbaguidi, A.; Mondo, J.M.; Muyideen, Y.; Adebola, P.O.; Asiedu, R. Diversity of White Guinea Yam (Dioscorea rotundata Poir.) Cultivars from Benin as Revealed by Agro-Morphological Traits and SNP Markers. Plant Genet. Resour. 2021, 19, 437–446. [Google Scholar] [CrossRef]
- Cao, J.C.; Jiang, D.; Zhao, Z.Y. Development of Chloroplast Genomic Resources in Chinese Yam. BioMed Res. Int. 2018, 2018, 6293847. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Li, J.; Luo, Z.X.; Yuan, S.B.; Zhang, Y.J.; Zhang, T.; Zhong, W.H.; Yuan, Q.J.; Huang, L.Q. Characterization and Development of EST-Derived SSR Markers in Cultivated Sweetpotato (Ipomoea batatas). BMC Plant Biol. 2011, 11, 139. [Google Scholar] [CrossRef]
- Cloutier, S.; Niu, Z.; Datla, R.; Duguid, S. Development and Analysis of EST-SSRs for Flax (Linum usitatissimum L.). Theor. Appl. Genet. 2009, 119, 53–63. [Google Scholar] [CrossRef]
- Liang, X.; Chen, X.; Hong, Y.; Liu, H.; Zhou, G.Y.; Li, X.S.; Guo, B.Z. Utility of EST-Derived SSR in Cultivated Peanut (Arachis hypogaea L.) and Arachis Wild Species. BMC Plant Biol. 2009, 9, 35. [Google Scholar] [CrossRef]
- Wei, W.L.; Qi, X.Q.; Wang, L.H.; Zhang, Y.X.; Hua, W.; Li, D.H.; Lv, H.X.; Zhang, X.R. Characterization of the Sesame (Sesamum indicum L.) Global Transcriptome Using Illumina Paired-End Sequencing and Development of EST-SSR Markers. BMC Genom. 2011, 12, 451. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, X.; He, Q.; Liu, X.X.; Xu, W.L.; Li, L.B.; Gao, J.W.; Wang, F.D. Transcriptome Analysis of the Roots at Early and Late Seedling Stages Using Illumina Paired-End Sequencing and Development of EST-SSR Markers in Radish. Plant Cell Rep. 2012, 31, 1437–1447. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, R.K.; Hendre, P.S.; Varshney, R.K.; Bhat, P.R.; Krishnakumar, V.; Singh, L. Identification, Characterization and Utilization of Est-Derived Genic Microsatellite Markers for Genome Analyses of Coffee and Related Species. Theor. Appl. Genet. 2007, 114, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.J.; Yang, C.; Tian, B.; Yang, J.B.; Liu, A.Z. Exploiting EST Databases for the Development and Characterization of EST-SSR Markers in Castor Bean (Ricinus communis L.). BMC Plant Biol. 2010, 10, 278. [Google Scholar] [CrossRef]
- Kumpatla, S.; Mukhopadhyay, S. Mining and Survey of Simple Sequence Repeats in Expressed Sequence Tags of Dicotyledonous Species. Genome 2005, 48, 985–998. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Fang, B.P.; Chen, J.Y.; Zhang, X.J.; Luo, Z.X.; Huang, L.F.; Chen, X.L.; Li, Y.J. De Novo Assembly and Characterization of Root Transcriptome Using Illumina Paired-End Sequencing and Development of cSSR Markers in Sweet Potato (Ipomoea batatas). BMC Genom. 2010, 11, 726. [Google Scholar] [CrossRef]
- Wróblewska, A. Genetic Diversity and Spatial Genetic Structure of Chamaedaphne Calyculata (Ericaceae) at the Western Periphery in Relation to Its Main Continuous Range in Eurasia. Folia Geobot. 2013, 49, 193–208. [Google Scholar] [CrossRef]
- Huang, Y.X.; Huang, S.; Liang, K.J.; Ma, H.B.; Lei, F.G.; Hua, S.M.; Xu, X.M. Genetic Diversity of Germplasm Resources on Dioscorea by SRAP Markers. Chin. Wild Plant Resour. 2011, 30, 48–54. [Google Scholar] [CrossRef]
- Hua, S.M.; Tu, Q.C.; Lei, F.G. Genetic Diversity of Dioscorea polystachya Turcz. Revealed by RAPD Markers. J. Plant Genet. Resour. 2009, 10, 195–200. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).