
The Effective Pollination Period of European Plum (Prunus domestica L.) Cultivars in Western Norway


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Air Temperature and Precipitation
2.3. Flowering, Pollen Collection and Pollen Germination In Vitro
2.4. Emasculation and Pollination
2.5. Fruit Set
2.6. Statistical Analysis
3. Results
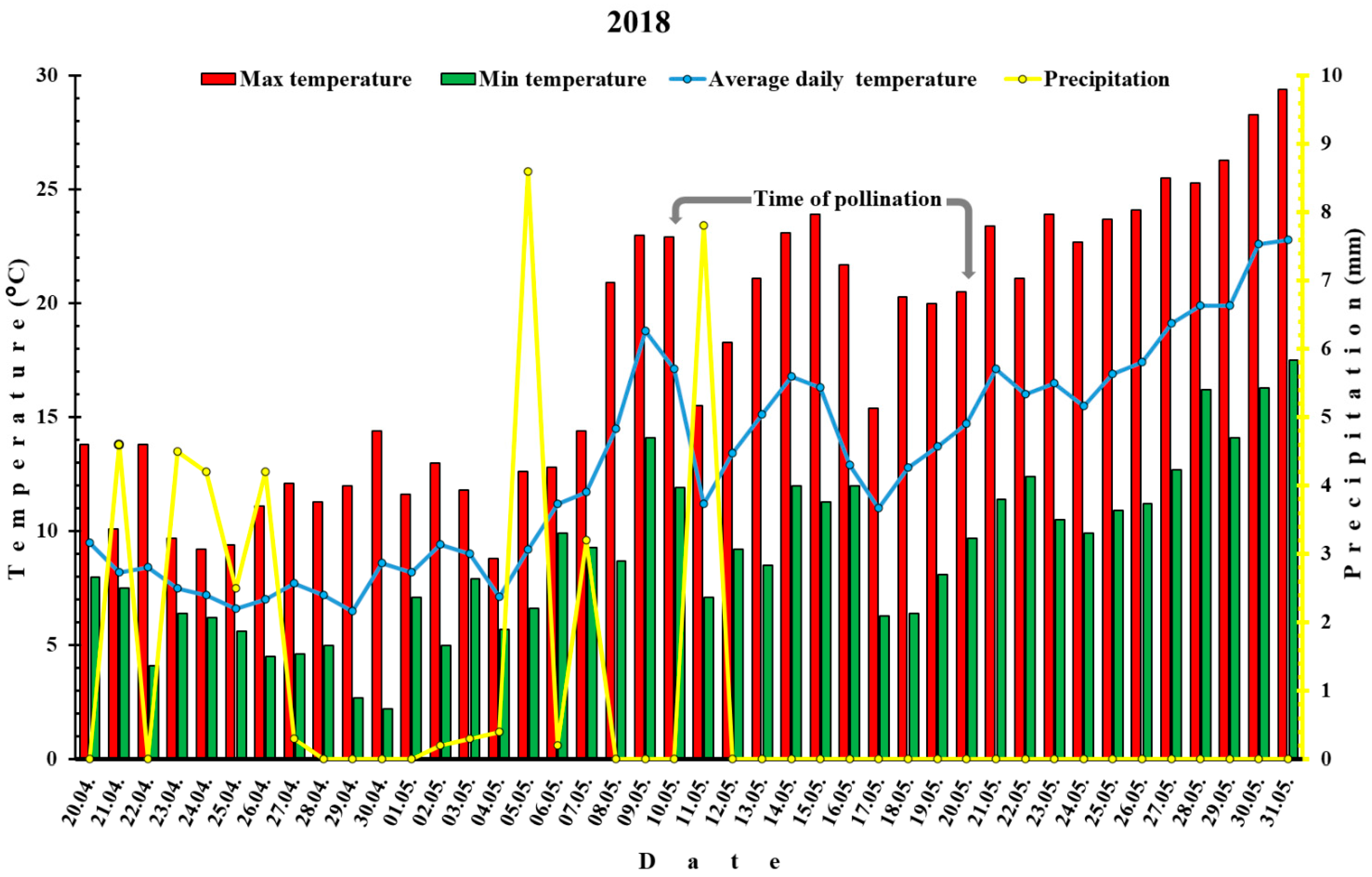
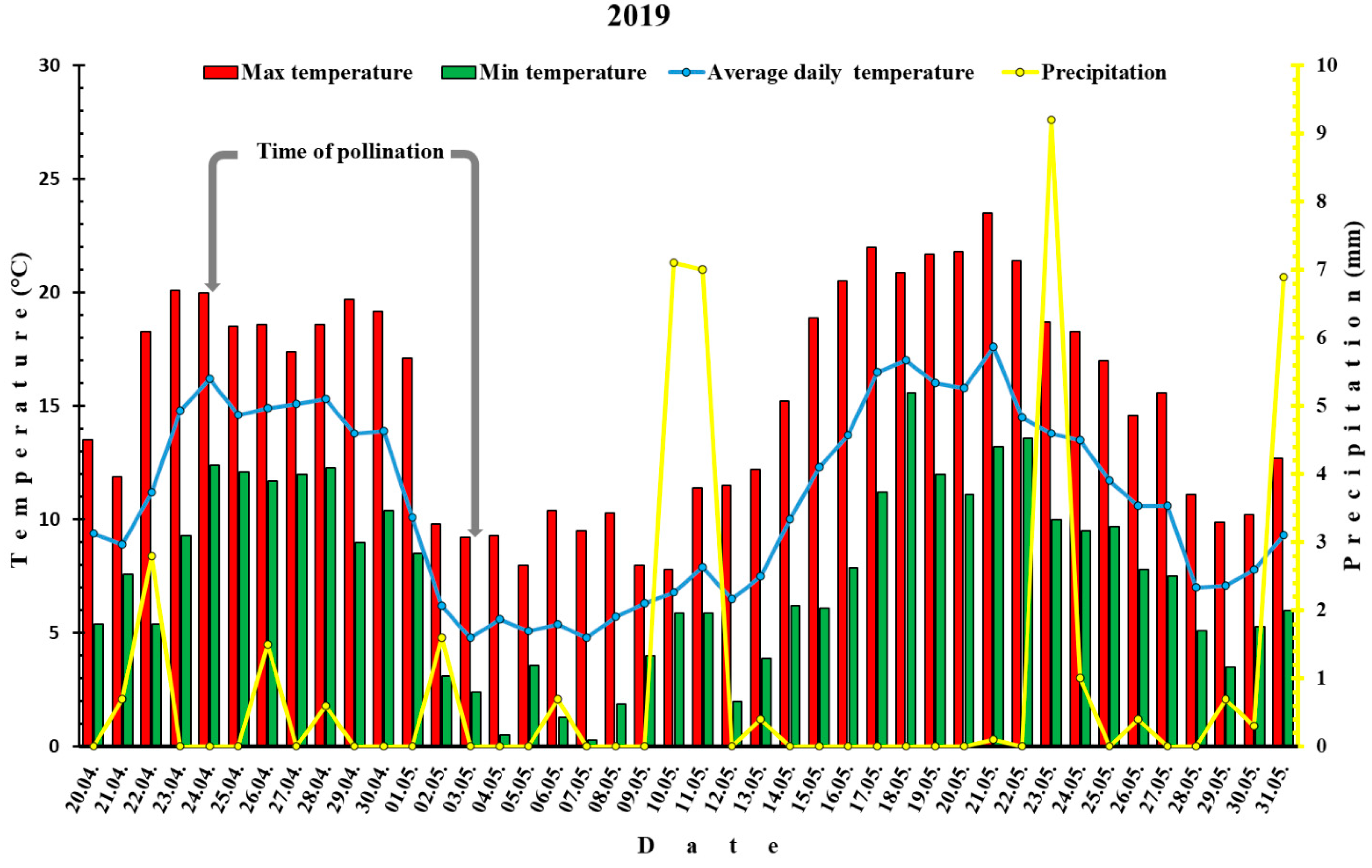
3.1. Meteorological Data
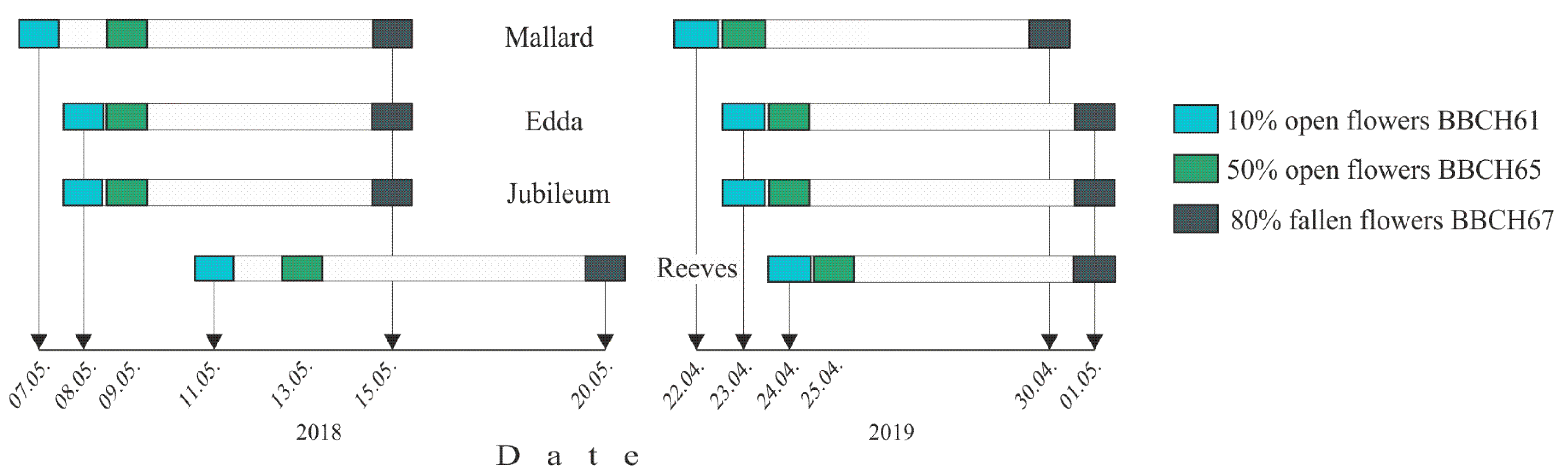
3.2. Flowering and In Vitro Pollen Germination
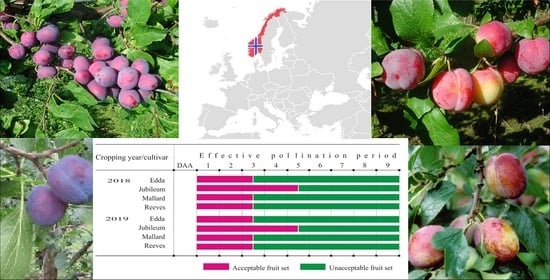
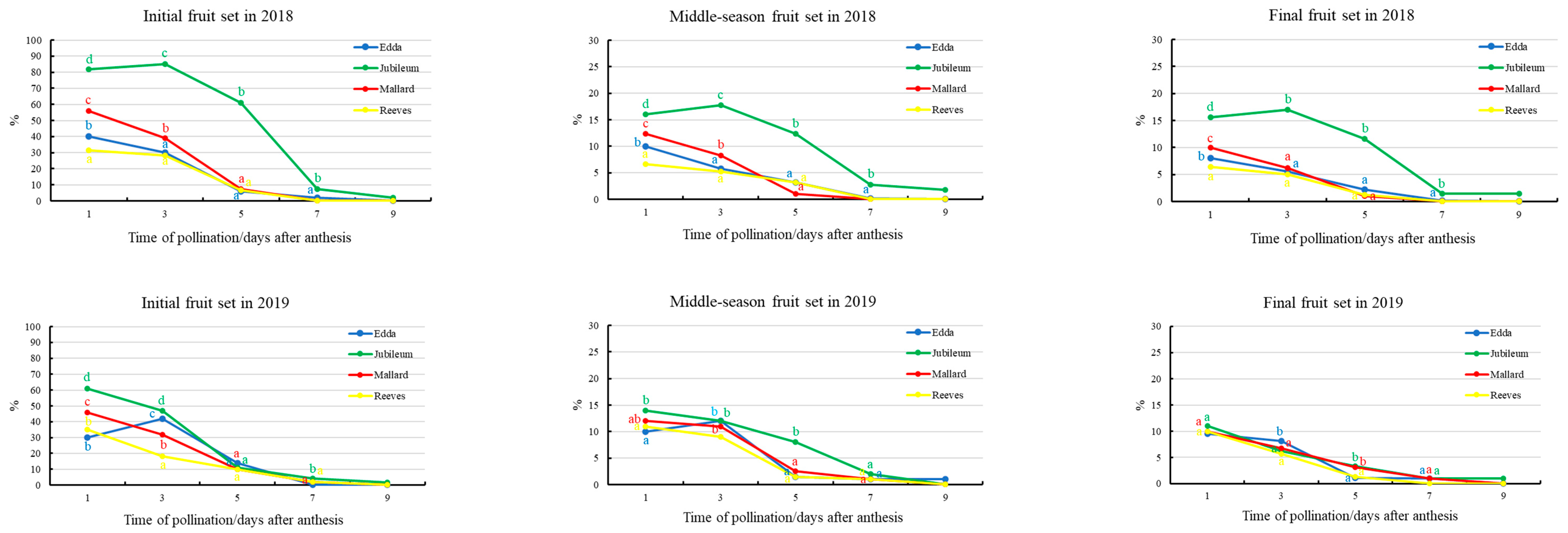
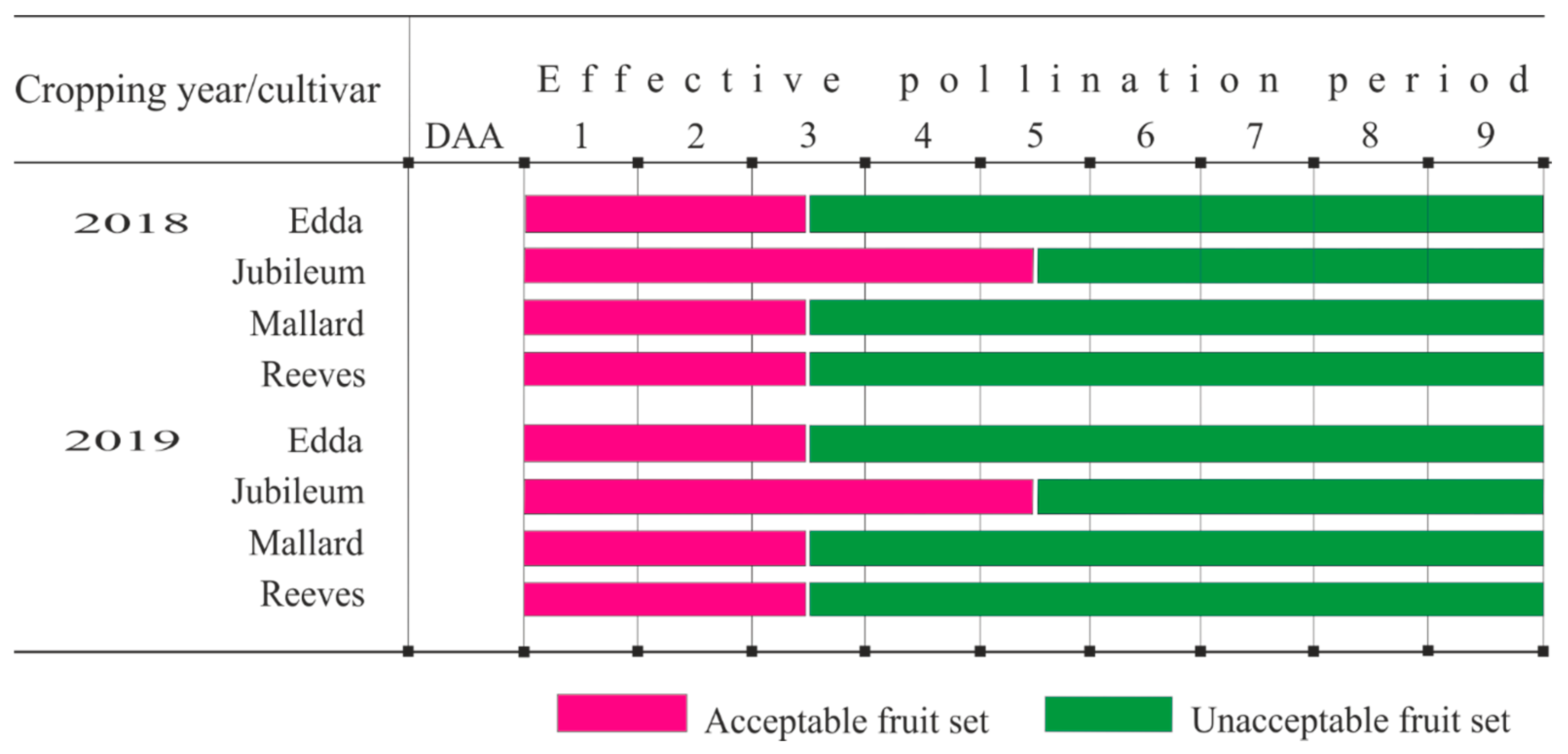
3.3. Fruit Set and Effective Pollination Period
4. Discussion
4.1. Flowering
4.2. Effective Pollination Period
4.3. Fruit Set
4.4. Correlation between Effective Pollination Period and Fruit Set
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOStat. 2019. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 24 October 2021).
- Sharma, D.D.; Kumar, M.; Singh, N.; Shylla, B. Plant growth and fruiting behavior of newly introduced plum (Prunus salicina Lindl.) cultivars under mid-hills conditions of Himachal Pradesh. Pharm. Innov. J. 2018, 7, 408–413. [Google Scholar]
- Redalen, G. Plum growing in Norway at 60° N. Acta Hortic. 2002, 577, 385–389. [Google Scholar]
- Hartmann, W.; Neümuller, M. Plum breeding. In Breeding Plantation Tree Crops: Temperate Species; Jain, S.M., Priyadarshan, P.M., Eds.; Springer Science: Stuttgart, Germany, 2009; pp. 161–231. [Google Scholar]
- Wertheim, S.J. Chemical thinning of deciduous fruit trees. Acta Hortic. 1998, 463, 445–462. [Google Scholar] [CrossRef]
- Szabó, Z.; Nyéki, J. Floral biology and fertility in peaches (Review article). Int. J. Hortic. Sci. 2000, 6, 11–27. [Google Scholar] [CrossRef]
- Meland, M.; Maas, F.M. Regulation of fruiting in plum production. In Proceedings of the 6th Conference „Innovation in Fruit Growing”, Belgrade, Serbia, 2 February 2017; Faculty of Agriculture: Belgrade, Serbia, 2017; pp. 51–67. [Google Scholar]
- Fotirić Akšić, M.; Rakonjac, V.; Nikolić, D.; Zec, G. Reproductive biology traits affecting productivity of sour cherry. Pesqui. Agropecu. Bras. 2013, 48, 33–41. [Google Scholar] [CrossRef][Green Version]
- Herrero, M. Male and female synchrony and the regulation of mating in flowering plants. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2003, 358, 1019–1024. [Google Scholar] [CrossRef]
- Ðorđević, M.; Cerović, R.; Radičević, S.; Nikolić, D.; Milošević, N.; Glišić, I.; Marić, S.; Lukić, M. Pollen Tube Growth and Embryo Sac Development in ‘Pozna Plava’ Plum Cultivar Related to Fruit Set. Erwerbs-Obstbau 2019, 61, 313–322. [Google Scholar] [CrossRef]
- Koskela, E.; Kemp, H.; van Dieren, M.C.A. Flowering and Pollination Studies with European Plum (Prunus domestica L.) Cultivars. Acta Hortic. 2010, 874, 193–201. [Google Scholar] [CrossRef]
- Fotirić Akšić, M.; Rakonjac, V.; Nikolić, D.; Čolić, S.; Milatović, D.; Ličina, V.; Rahović, D. Effective pollination period in ‘Oblačinska’ sour cherry clones. Genetika 2014, 46, 671–680. [Google Scholar] [CrossRef]
- Gheshlaghi, E.A. Effective pollination period and its influence on fruit characteristics of Hayward’ kiwifruit. Adv. Hortic. Sci. 2019, 33, 537–542. [Google Scholar]
- Abrol, D.P.; Sharma, D.; Monobrullah, M. Abundance and diversity of pollinating insects visiting peach and plum flowers and their impact on fruit production. J. Res. SKUAST J. 2005, 4, 38–45. [Google Scholar]
- Williams, R.R. Pollination studies in fruit trees. II. The effective pollination period for some apple and pear varieties. Rep. Long Ashton Res. Stn. 1965, 1966, 136–138. [Google Scholar]
- Sanzol, J.; Herrero, M. The effective pollination period in fruit trees. Scientia Hortic. 2001, 90, 1–17. [Google Scholar] [CrossRef]
- Cerović, R.; Ružić, Đ.; Mićić, N. Viability of plum ovules at different temperatures. Ann. Appl. Biol. 2000, 137, 53–59. [Google Scholar] [CrossRef]
- Zhebentyayeva, T.; Shankar, V.; Scorza, R.; Callahan, A.; Ravelonandro, M.; Castro, S.; DeJong, T.; Saski, C.A.; Dardick, C. Genetic characterization of worldwide Prunus domestica (plum) germplasm using sequence-based genotyping. Hortic. Res. 2019, 6, 12. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants: BBCH Monograph, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry: Berlin, Germany; Brunswick, Germany, 2001; pp. 1–158. [Google Scholar]
- Meland, M.; Frøynes, O.; Fotiric Akšić, M.; Pojskić, N.; Kalamujić Stroil, B.; Lašić, L.; Gaši, F. Identifying Pollen Donors and Success Rate of Individual Pollinizers in European Plum (Prunus domestica L.) Using Microsatellite Markers. Agronomy 2020, 10, 264. [Google Scholar] [CrossRef]
- Fotirić Akšić, M.; Cerović, R.; Rakonjac, V.; Bakić, I.; Čolić, S.; Meland, M. Vitality and in vitro pollen germination in ‘Oblačinska’ sour cherry clones. Genetika 2017, 49, 791–800. [Google Scholar] [CrossRef]
- Cerović, R.; Fotirić Akšić, M.; Đorđević, M.; Meland, M. The effects of pollinizers on pollen tube growth and fruit set of European plum (Prunus domestica L.) in a Nordic climate. Sci. Hortic. 2021, 288, 110390. [Google Scholar]
- Ohata, K.; Togano, Y.; Matsumoto, T.; Uchida, Y.; Kurahashi, T.; Itamura, H. Selection of Prune (Prunus domestica L.) Cultivars Suitable for the East Asian Temperate Monsoon Climate: Ripening Characteristics and Fruit Qualities of Certain Prunes in a Warm Southwest Region of Japan. Hortic. J. 2017, 86, 437–446. [Google Scholar] [CrossRef]
- Đorđević, M.; Radičević, S.; Cerović, R.; Milošević, N.; Mitrović, M. Initial and final fruit set in plum cultivar ‘Pozna Plava’ as affected by different types of pollination. Acta Hortic. 2012, 968, 121–124. [Google Scholar] [CrossRef]
- Rodrigo, J.; Herrero, M. The onset of fruiting in apricot (Prunus armeniaca L.). J. Appl. Bot. 2002, 76, 13–19. [Google Scholar]
- Hedhly, A.; Hormaza, J.I.; Herrero, H. Warm temperatures at bloom reduce fruit set in sweet cherry. J. Appl. Bot. Food Qual. 2007, 81, 158–164. [Google Scholar]
- Hedhly, A.; Hormaza, J.I.; Herrero, M. Flower emasculation accelerates ovule degeneration and reduces fruit set in sweet cherry. Sci. Hortic. 2009, 119, 455–457. [Google Scholar] [CrossRef]
- Yoder, K.; Yuan, R.C.; Combs, L.; Byers, R.; Mcferson, J.; Schmidt, T. Effects of temperature and the combination of liquid lime sulfur and fish oil on pollen germination, pollen tube growth, and fruit set in apples. Hortic. Sci. 2009, 44, 1277–1283. [Google Scholar] [CrossRef]
- Irenaeus, T.K.S.; Mitra, S.K. Understanding the pollen and ovule characters and fruit set of fruit crops in relation to temperature and genotype—A review. J. Appl. Bot. Food Qual 2014, 87, 157–167. [Google Scholar]
- Guerrero-Prieto, V.M.; Rascón-Chu, A.; Romo-Chacón, A.; Berlanga-Reyes, D.I.; Orozco-Avitia, J.A.; Gardea-Béjar, A.A.; Parra-Quezada, R.; Sánchez-Chávez, E. Effective pollination period in ‘Red Chief’ and ‘Golden Delicious’ apples (Malus domestica Borkh). Span. J. Agric. Res. 2009, 7, 928–932. [Google Scholar] [CrossRef]
- Guerra, M.E.; Wünsch, A.; López-Corrales, M.; Rodrigo, J. Flower emasculation as the cause for lack of fruit set in Japanese plum crosses. J. Am. Soc. Hortic. Sci. 2010, 135, 556–562. [Google Scholar] [CrossRef]
- Toyama, T.K. The pollen receptivity period and its relation to fruit setting in the stone fruits. Fruit Var. J. 1980, 34, 2–4. [Google Scholar]
- Cuevas, J.; Pinillos, V.; Polito, V.S. Effective pollination period for ‘Manzanillo’ and ‘Picual’ olive trees. J. Hortic. Sci. Biotechnol. 2009, 84, 370–374. [Google Scholar] [CrossRef]
- Roeder, S.; Serra, S.; Musacchi, S. Effective Pollination Period and Parentage Effect on Pollen Tube Growth in Apple. Plants 2021, 10, 1618. [Google Scholar] [CrossRef] [PubMed]
- Sanzol, J.; Rallo, P.; Herrero, M. Asynchronous development of stigmatic receptivity in the pear (Pyrus communis; Rosaceae) flower. Am. J. Bot. 2003, 90, 78–84. [Google Scholar] [CrossRef]
- Kaufmane, E.; Rumpunen, K. Pollination, pollen tube growth and fertilization in Chaenomeles japonica (Japanese quince). Sci. Hortic. 2002, 94, 257–271. [Google Scholar] [CrossRef]
- Rezaie, S.F.; Hajilou, J.; Nahandi, F.Z. Pollen germination and pistil performance in several Iranian peach cultivars. Int. J. Agric. Sci. 2011, 1, 71–77. [Google Scholar]
- Nejatian, M.; Arzani, K. Determination of self-incompatibility and effective pollination period in four local Iranian apricot (Prunus armeniaca L.) cultivars. Iran. Int. J. Hortic. Sci. Technol. 2004, 5, 147–156. [Google Scholar]
- Hedhly, A.; Hormaza, J.I.; Herrero, M. The effect of temperature on stigmatic receptivity in sweet cherry (Prunus avium L.). Plant Cell Environ. 2003, 26, 1673–1680. [Google Scholar] [CrossRef]
- Kodad, O.; Oukabli, A.; Mamouni, A.; Lahlou, M.; Socias i Company, R. Flowering and pollination time affect fruit set of foreign almond cultivars in Morocco. Acta Hortic. 2011, 912, 103–106. [Google Scholar] [CrossRef]
- Selak, G.V.; Cuevas, J.; Ban, S.G.; Pinillos, V.; Dumicic, G.; Perica, S. The effect of temperature on the duration of the effective pollination period in ‘Oblica’ olive (Olea europaea) cultivar. Ann. Appl. Biol. 2014, 164, 85–94. [Google Scholar] [CrossRef]
- Tromp, J.; Visser, K.A.; Dijkstra, J. Fruit set and the effective pollination period in red currant as affected by nitrogen fertilization and exposure to red light. J. Hortic. Sci. 1994, 69, 791–797. [Google Scholar] [CrossRef]
- Brevis, P.A.; Nesmith, D.S.; Wetzstein, H.Y. Flower age affects fruit set and stigmatic receptivity in rabbiteye blueberry. Hortic. Sci. 2006, 41, 1537–1540. [Google Scholar] [CrossRef]
- Ferradás, Y.; López, M.; Rey, M.; Victoria González, M. Programmed cell death in kiwifruit stigmatic arms and its relationship to the effective pollination period and the progamic phase. Ann. Bot. 2014, 114, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Dag, A.; Eisenstein, D.; Gazit, S. Effect of temperature regime on pollen and the effective pollination of ‘Kent’ mango in Israel. Sci. Hortic. 2000, 86, 1–11. [Google Scholar] [CrossRef]
- Alcaraz, M.L.; Hormaza, J.I. Avocado Pollination and Fruit Set—A Perspective from Spain. Calif. Avocado Soc. Yearb. 2009, 92, 113–135. [Google Scholar]
- Mesejo, C.; Martínez-Fuentes, A.; Reig, C.; Agustí, M. The effective pollination period in ‘Clemenules’ mandarin, ‘Owari’ Satsuma mandarin and ‘Valencia’ sweet orange. Plant Sci. 2007, 173, 223–230. [Google Scholar] [CrossRef]
- Guerra, M.E.; Wünsch, A.; López-Corrales, M.; Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. J. Amer. Soc. Hortic. Sci. 2011, 136, 375–381. [Google Scholar] [CrossRef]
- Keulemans, J.; Van Laer, H. Effective pollination period of plums: The influence of temperature on pollen germination and pollen tube growth. In Manipulation of Fruiting; Wright, C.J., Ed.; Butterworths: London, UK, 1989; pp. 159–171. [Google Scholar]
- Nikolić, D.T.; Rakonjac, V.S.; Fotirić-Akšić, M. The effect of pollenizer on the fruit set of plum cultivar Čačanska najbolja. J. Agric. Sci. 2012, 57, 9–18. [Google Scholar] [CrossRef]
- Goldway, M.; Stern, R.; Zisovich, A.; Raz, A.; Sapir, G.; Schnieder, D.; Nyska, R. The self-incompatibility fertilization system in Rosaceae: Agricultural and genetic aspects. Acta Hortic. 2012, 967, 77–82. [Google Scholar] [CrossRef]
- Sharafi, Y. In vitro pollen germination in stone fruit tree of Rosaceae family. Afr. J. Agric. Res. 2011, 6, 6021–6026. [Google Scholar]
- Đorđević, M.; Cerović, R.; Radičević, S.; Nikolić, D.; Marić, S.; Milosević, N.; Glišić, I.S. Influence of pollination mode on fruit set in plum (Prunus domestica). Acta Hortic. 2016, 1139, 347–352. [Google Scholar] [CrossRef]
- Glišić, I.; Cerović, R.; Milošević, N.; Đorđević, M.; Radičević, S. Initial and final fruit set in some plum (Prunus domestica L.) hybrids under different pollination types. Genetika 2012, 44, 583–593. [Google Scholar] [CrossRef]
- Kron, P.; Husband, B.C. The effects of pollen diversity on plant reproduction: Insights from apple. Sex. Plant Reprod. 2006, 19, 125–131. [Google Scholar] [CrossRef]
- Janssen, B.J.; Thodey, K.; Schaffer, R.J.; Alba, R.; Balakrishnan, L.; Bishop, R.; Bowen, J.H.; Crowhurst, R.N.; Gleave, A.P.; Ledger, S.; et al. Global gene expression analysis of apple fruit development from the floral bud to ripe fruit. BMC Plant Biol. 2008, 8, 16. [Google Scholar] [CrossRef]
- Bonghi, C.; Trainotti, L.; Botton, A.; Tadiello, A.; Rasori, A.; Ziliotto, F.; Zaffalon, V.; Casadoro, G.; Ramina, A. A microarray approach to identify genes involved in seed-pericarp cross-talk and development in peach. BMC Plant Biol. 2011, 11, 107. [Google Scholar] [CrossRef]
- Nyomora, A.M.S.; Brown, P.H.; Pinney, K.; Polito, V.S. Foliar application of boron to almond trees affects pollen quality. J. Am. Soc. Hortic. Sci. 2000, 125, 265270. [Google Scholar] [CrossRef]
- Meland, M.; Birken, E. Ethephon as a Blossom and Fruitlet Thinner Affects Crop Load, Fruit Weight and Fruit Quality of the European Plum Cultivar ‘Jubileum’. Acta Hortic. 2010, 884, 315–321. [Google Scholar] [CrossRef]
- Snider, J.L.; Oosterhuis, D.M.; Loka, D.A.; Kawakami, E.M. High temperature limits in vivo pollen tube growth rates by altering diurnal carbohydrate balance in field-grown Gossypium hirsutum pistils. J. Plant Physiol. 2011, 168, 1168–1175. [Google Scholar] [CrossRef]
- Weinbaum, S.A.; Parfitt, D.E.; Polito, V.S. Differential cold sensitivity of pollen grain germination in two Prunus species. Euphytica 1984, 33, 419–426. [Google Scholar] [CrossRef]
- Egea, J.; Burgos, L. Effective pollination period as related to stigma receptivity in apricot. Sci. Hortic. 1992, 52, 77–83. [Google Scholar]
- Kodad, O.; Socias i Company, R. Effect of pollination time on fruit set in an autogamous almond cultivar. J. Hortic. Sci. Biotechnol. 2009, 84, 350–354. [Google Scholar] [CrossRef]
- Guerrero-Prieto, V.M.; Vasilakakis, M.D.; Lombard, P.B. Factors controlling fruit-set of Napoleon sweet cherry in Western Oregon. HortScience 1985, 20, 913. [Google Scholar]
- Burgos, L.; Egea, J.; Dicenta, F. Effective pollination period in apricot (Prunus armeniaca L.) varieties. Ann. Appl. Biol. 1991, 119, 533–539. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | 1 DAA | 3 DAA | 5 DAA | 7 DAA | 9 DAA |
|---|---|---|---|---|---|
| Years | |||||
| 2018 | 24.5 ± 1.2 b * | 21.1 ± 1.1 b | 9.71 ± 0.4 b | 0.94 ± 0.07 a | 0.43 ± 0.3 b |
| 2019 | 21.63 ± 1.1 a | 17.48 ± 1.0 a | 5.61 ± 0.2 a | 1.33 ± 0.08 b | 0.23 ± 0.2 a |
| Cultivars | |||||
| ‘Edda’ | 17.92 ± 1.0 a | 17.23 ± 1.0 b | 4.63 ± 0.2 a | 0.7 ± 0.06 a | 0.17 ± 0.01 b |
| ‘Jubileum’ | 33.23 ± 1.8 c | 30.83 ± 1.7 c | 8.47 b ± 1.0 b | 3.08 ± 0.2 b | 1.15 ± 0.08 c |
| ‘Mallard’ | 24.38 b ± 1.2 b | 17.22 ± 1.0 b | 4.15 ± 0.2 a | 0.67 ± 0.06 a | 0 a |
| ‘Reeves’ | 16.72 ± 1.0 a | 11.88 ± 0.5 a | 4.0 ± 0.2 a | 0.5 ± 0.05 a | 0 a |
| Cultivars × Years | |||||
| ‘Edda’ 2018 | 19.33 ± 1.0 b | 13.77 ± 0.4 ab | 3.73 ± 0.2 a | 0.73 ± 0.05 b | 0.00 a |
| ‘Edda’ 2019 | 16.50 ± 0.5 a | 20.70 ± 1.1 c | 5.53 ± 0.2 a | 0.67 ± 0.05 b | 0.33 ± 0.3 b |
| ‘Jubileum’ 2018 | 37.80 ± 1.9 d | 39.93 ± 1.9 d | 11.6 ± 1.6 b | 3.83 ± 0.2 d | 1.73 ± 0.08 c |
| ‘Jubileum’ 2019 | 28.67 ± 1.6 c | 21.73 ± 1.5 c | 5.33 ± 0.3 a | 2.33 ± 0.1 c | 0.57 ± 0.04 ab |
| ‘Mallard’ 2018 | 26.10 ± 1.5 c | 17.83 ± 0.6 b | 3.10 ± 0.2 a | 0.00 a | 0.00 a |
| ‘Mallard’ 2019 | 22.67 ± 1.4 bc | 16.60 ± 0.6 b | 5.20 ± 0.2 a | 1.33 ± 0.08 c | 0.00 a |
| ‘Reeves’ 2018 | 14.77 ± 0.8 a | 12.87 ± 0.2 a | 3.73 ± 0.2 a | 0.00 a | 0.00 a |
| ‘Reeves’ 2019 | 18.67 ± 1.0 ab | 10.90 ± 0.2 a | 4.27 ± 0.2 a | 1.00 ± 0.07 bc | 0.00 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fotirić Akšić, M.; Cerović, R.; Hjeltnes, S.H.; Meland, M. The Effective Pollination Period of European Plum (Prunus domestica L.) Cultivars in Western Norway. Horticulturae 2022, 8, 55. https://doi.org/10.3390/horticulturae8010055
Fotirić Akšić M, Cerović R, Hjeltnes SH, Meland M. The Effective Pollination Period of European Plum (Prunus domestica L.) Cultivars in Western Norway. Horticulturae. 2022; 8(1):55. https://doi.org/10.3390/horticulturae8010055
Chicago/Turabian StyleFotirić Akšić, Milica, Radosav Cerović, Stein Harald Hjeltnes, and Mekjell Meland. 2022. "The Effective Pollination Period of European Plum (Prunus domestica L.) Cultivars in Western Norway" Horticulturae 8, no. 1: 55. https://doi.org/10.3390/horticulturae8010055
APA StyleFotirić Akšić, M., Cerović, R., Hjeltnes, S. H., & Meland, M. (2022). The Effective Pollination Period of European Plum (Prunus domestica L.) Cultivars in Western Norway. Horticulturae, 8(1), 55. https://doi.org/10.3390/horticulturae8010055