
Exogenous Antioxidants Enhance Seedling Growth and Yield of Artificially Aged Cabbage and Lettuce Seeds



Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterisation of Seed Ageing Rates and Patterns via Controlled Deterioration
2.1.1. Seed Material
2.1.2. Seed Vigour Assessment
2.1.3. Controlled Deterioration
2.1.4. Germination Test
2.1.5. Seed Ageing Rates and Patterns
2.2. Assessment of the Influence of Exogenous Antioxidant Invigoration on Subsequent Seedling Performance of Aged Seeds of Cabbage and Lettuce
2.2.1. Application of Exogenous Antioxidant Solutions
2.2.2. Greenhouse Pot Trial
2.2.3. Seedling Vigour and Biomass Accumulation
2.2.4. Leaf Chlorophyll Content, Gas Exchange, and Chlorophyll Fluorescence
2.3. Statistical Analysis
3. Results
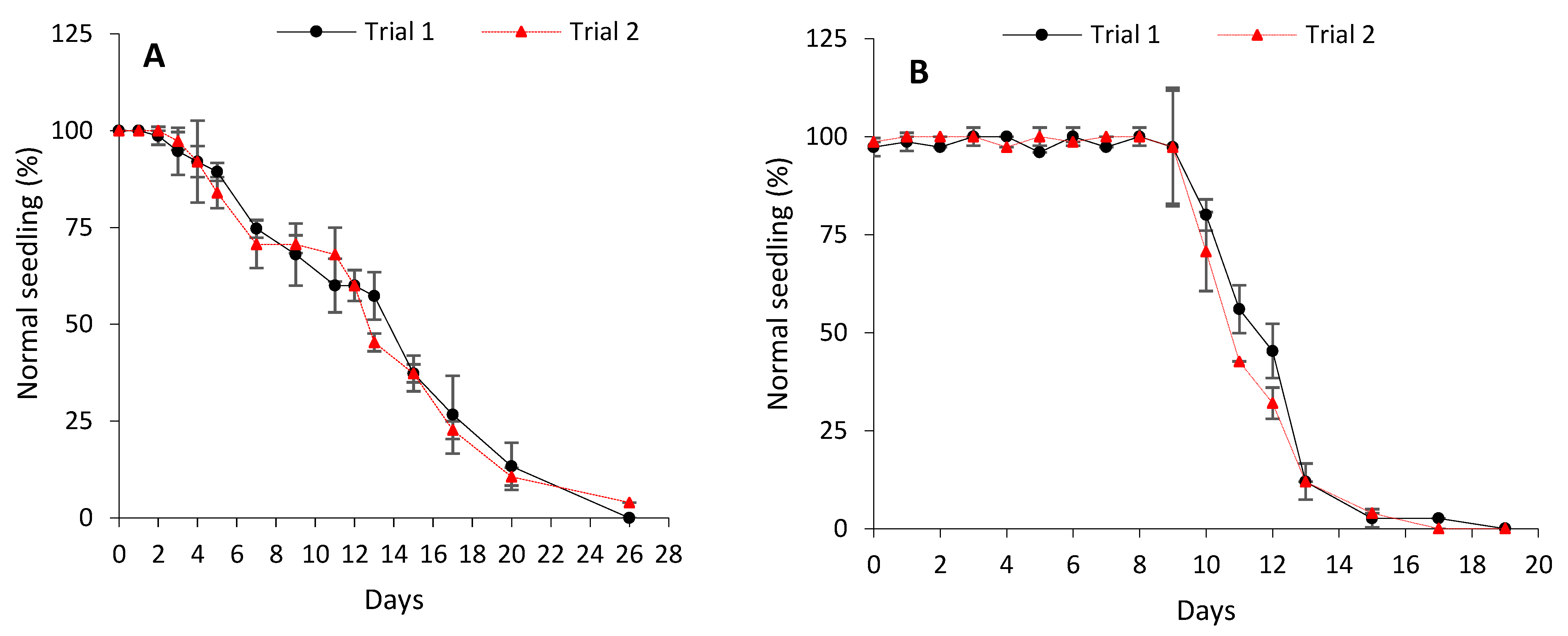
3.1. Characterisation of Seed Ageing Rates and Patterns via Controlled Deterioration
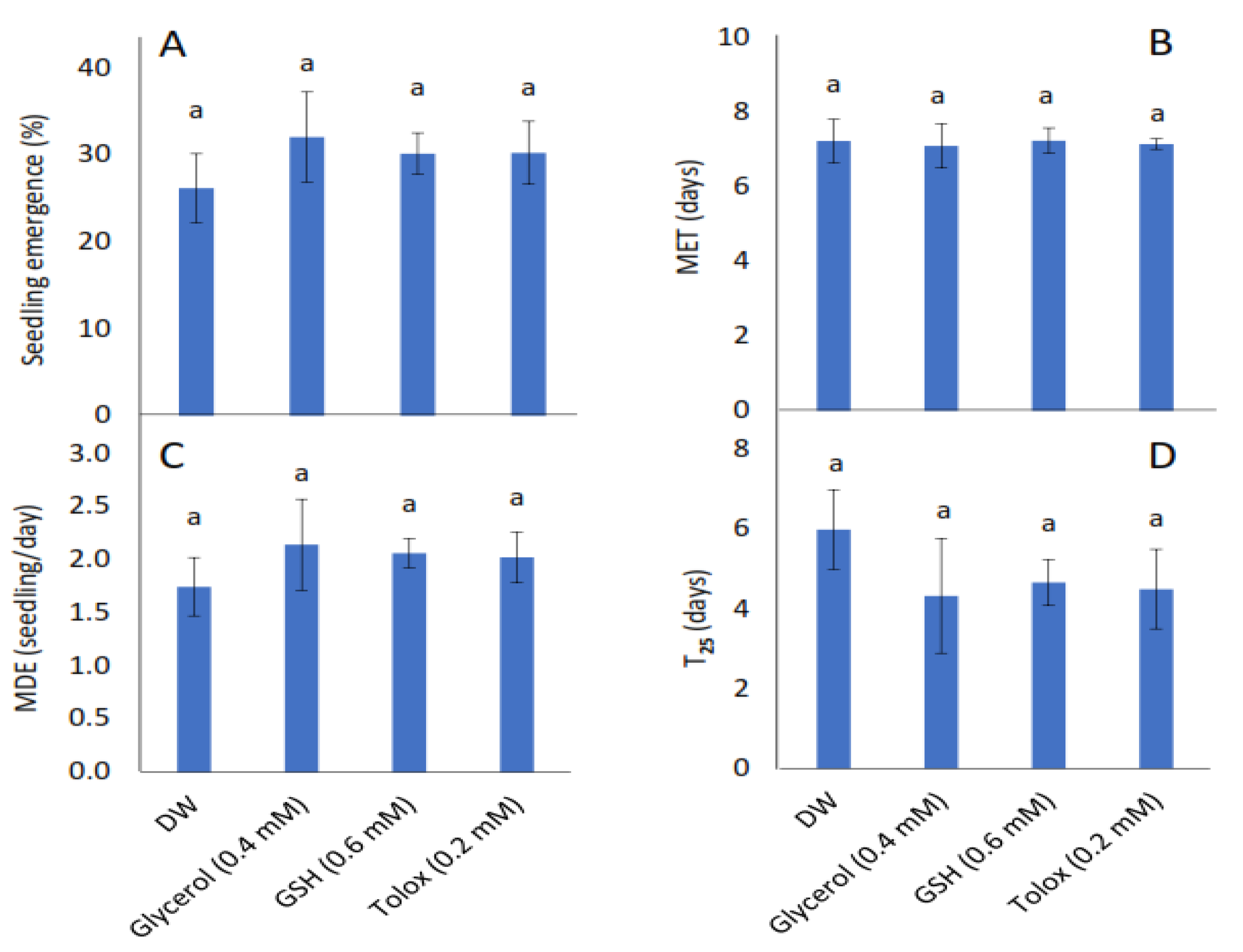
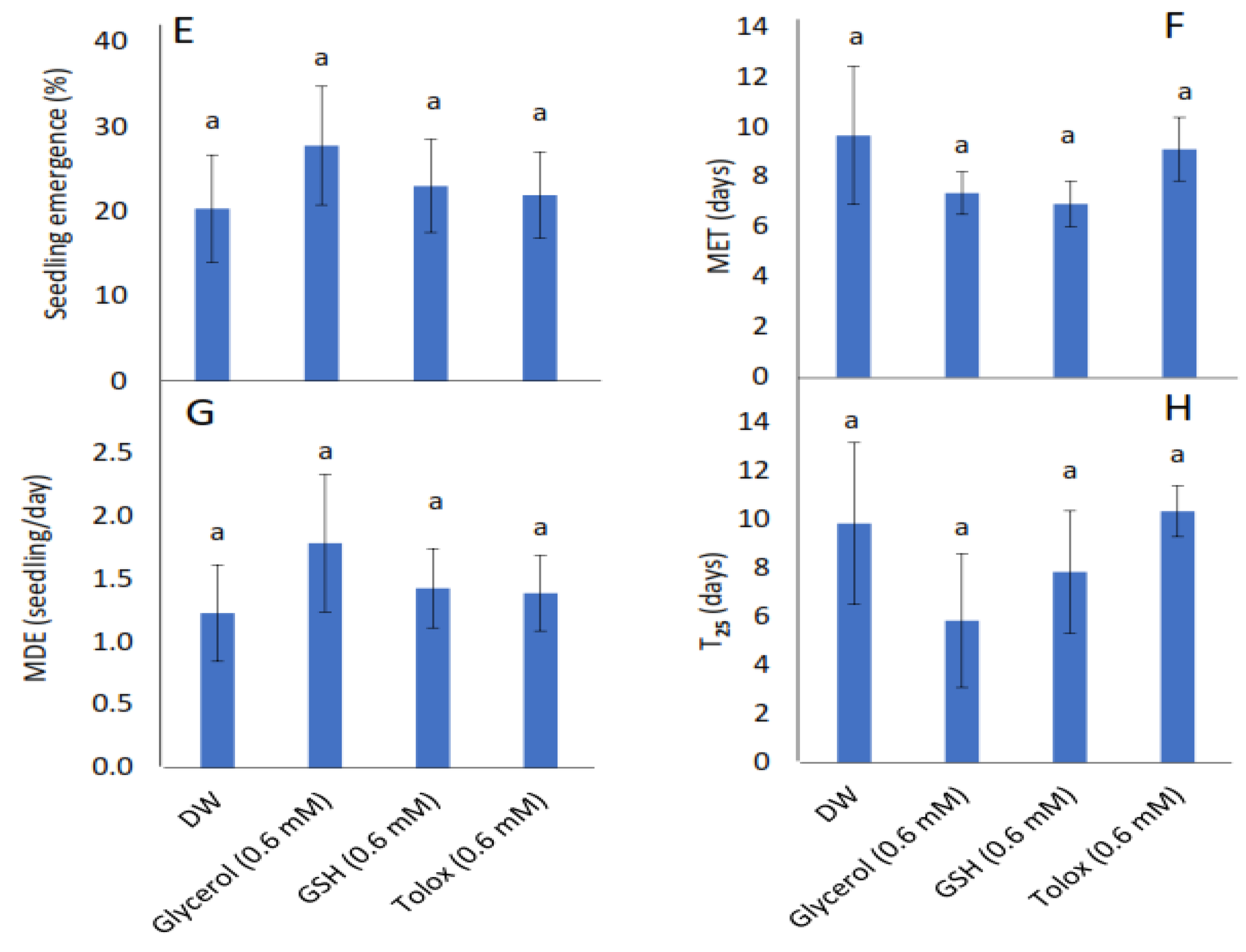
3.2. Effect of Exogenous Application of Antioxidants on Seedling Emergence Parameters of Controlled Deteriorated Cabbage and Lettuce Seeds
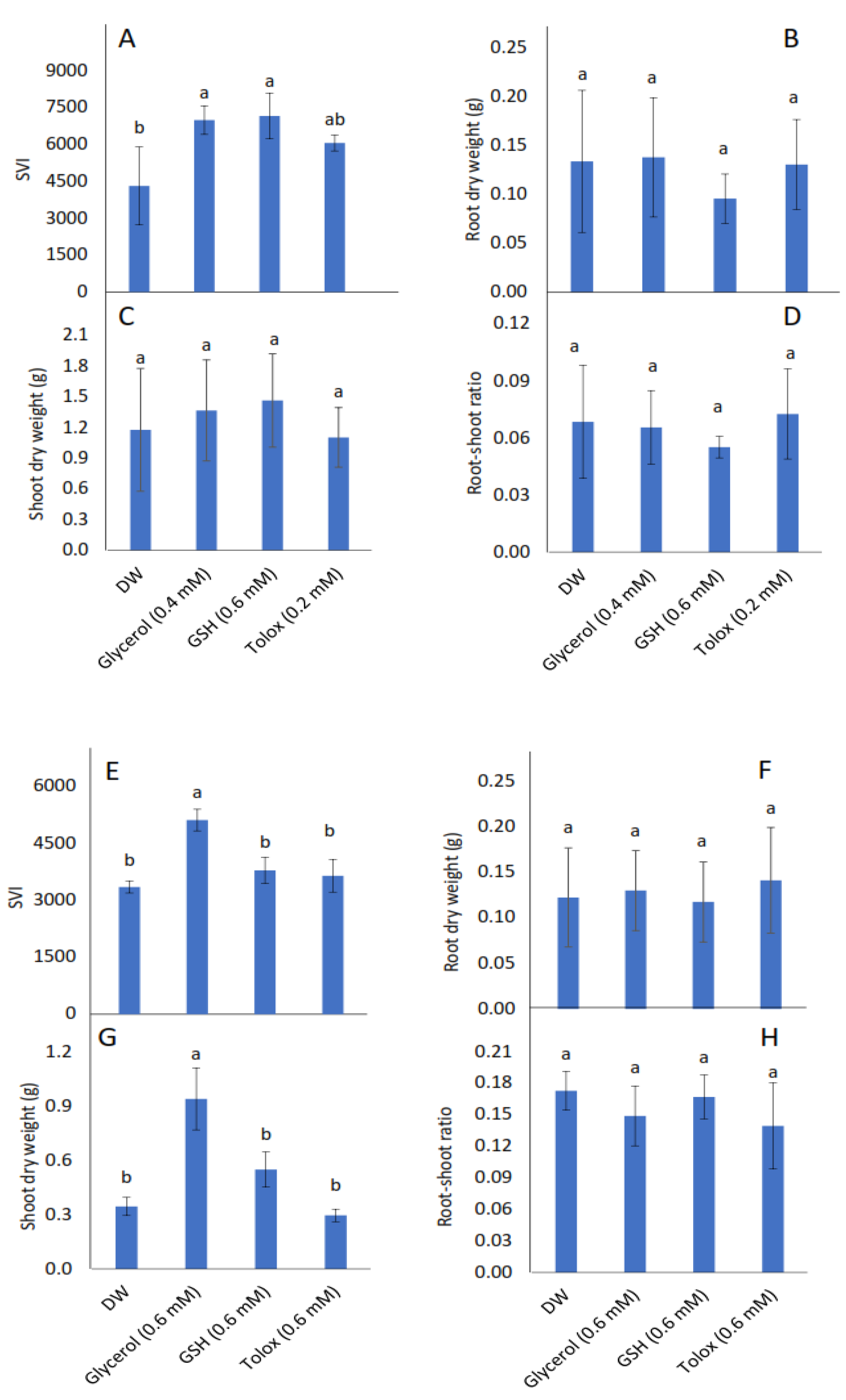
3.3. Effect of Exogenous Application of Antioxidants on Seedling Vigour and Biomass Accumulation of Seedlings Produced from Controlled Deteriorated Cabbage and Lettuce Seeds
3.4. Effect of Exogenous Application of Antioxidants on Leaf Area, Leaf Area Ratio, and Total Chlorophyll Content of Leaves from Seedlings Produced from Controlled Deteriorated Cabbage and Lettuce Seeds
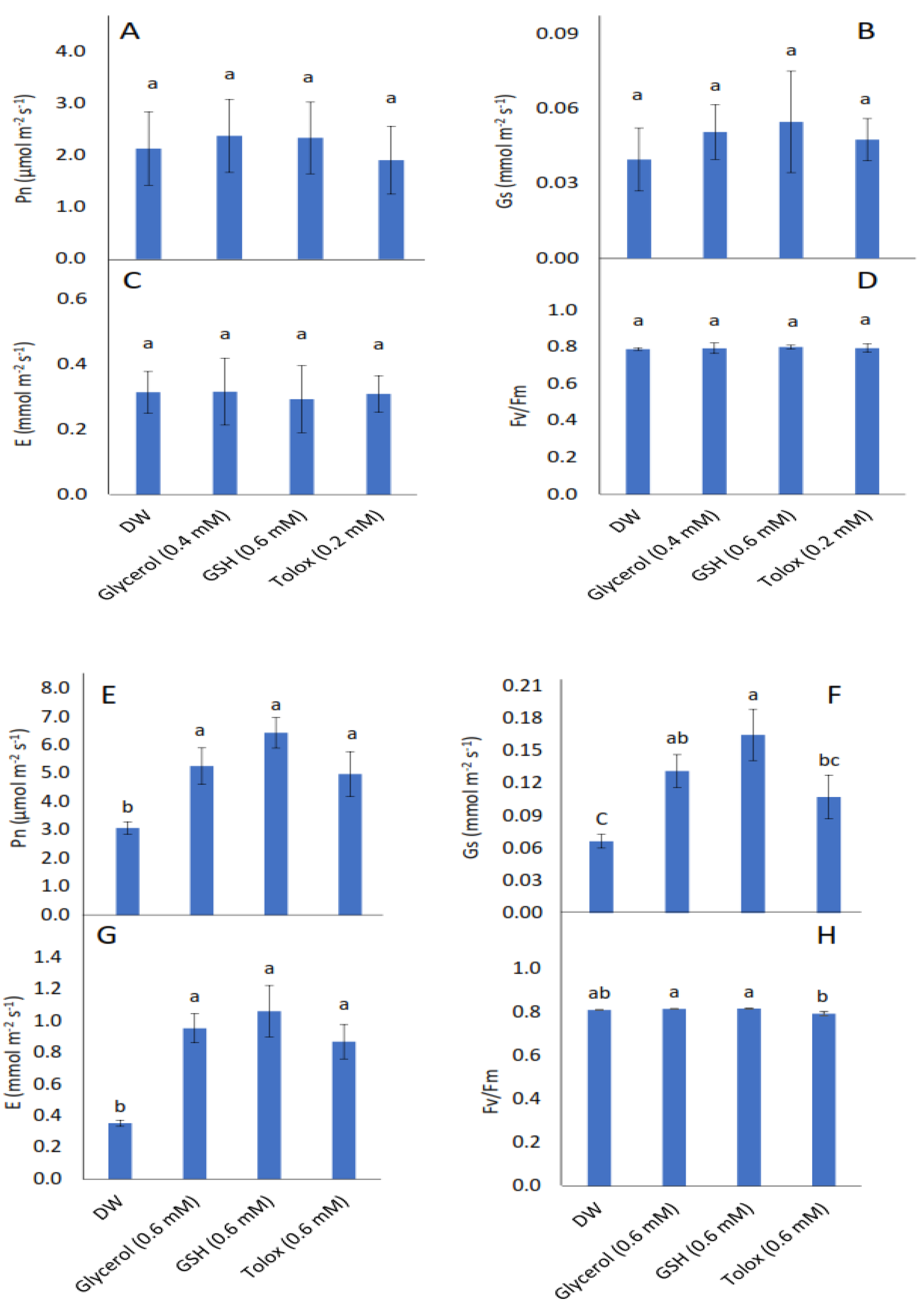
3.5. Effect of Exogenous Application of Antioxidants on Photosynthetic Rate, Gas Exchange, and Photochemistry of Seedlings Produced from Controlled Deteriorated Cabbage and Lettuce Seeds
4. Discussion
4.1. Differences in Rates and Patterns of Ageing in Cabbage and Lettuce Seeds Subjected to Controlled Deterioration
4.2. Influence of Exogenous Antioxidant Invigoration of Aged Cabbage and Lettuce Seeds on Subsequent Seedling Emergence, Growth, Gas Exchange, and Photochemistry
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rakshit, A.; Singh, H.B. (Eds.) Advances in Seed Priming; Springer: Singapore, 2018; ISBN 978-981-13-0031-8. [Google Scholar]
- Johnson, R.R.; Wax, L.M. Relationship of soybean germination and vigor tests to field performance. Agron. J. 1978, 70, 273–278. [Google Scholar] [CrossRef]
- Heydecker, W. Vigour. In Viability of Seeds; Roberts, E.H., Ed.; Springer: Dordrecht, The Netherlands, 1972; pp. 209–252. [Google Scholar]
- Hall, R.D.; Wiesner, L.E. Relationship between seed vigor tests and field performance of ‘regar’ meadow bromegrass. Crop Sci. 1990, 30, 967–970. [Google Scholar] [CrossRef]
- Mehrabadi, M.; Bandani, A.R. Assessing of α-amylase activity of midgut in wheat bug Eurygaster Maura. Am. J. Appl. Sci. 2009, 6, 478–483. [Google Scholar] [CrossRef] [Green Version]
- Matthews, S.; Collins, M.T. The effect of seed condition and fungicidal dressings on the field emergence of barley. In Proceedings of the 7th British Insecticide and Fungicide Conference, Brighton, UK, 19–22 November 1973; British Crop Protection Council: Nottingham, UK; pp. 135–141.
- Perry, D.A.; Harrison, J.G. Effects of seed deterioration and seed-bed environment on emergence and yield of spring-sown barley. Ann. Appl. Biol. 1977, 86, 291–300. [Google Scholar] [CrossRef]
- Stormonth, D.A.; Doling, D.A. The significance of seed vigour in cereals. Arab. Farm. 1979, 6, 42–46. [Google Scholar]
- Mirmazloum, I.; Kiss, A.; Erdélyi, É.; Ladányi, M.; Németh, É.Z.; Radácsi, P. The Effect of osmopriming on seed germination and early seedling characteristics of Carum carvi L. Agriculture 2020, 10, 94. [Google Scholar] [CrossRef] [Green Version]
- Kolasinska, K.; Szyrmer, J.; Dul, S. Relationship between laboratory seed qality tests and field emergence of common bean seed. Crop Sci. 2000, 40, 470–475. [Google Scholar] [CrossRef]
- Lubbe, E.; Rodda, N. Sershen Effects of greywater irrigation on germination, growth and photosynthetic characteristics in selected African leafy vegetables. Water SA 2016, 42, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Wilson Jr, D.O.; McDonald Jr, M.B. Lipid peroxidation model of seed ageing. Seed Sci. Technol. 1986, 14, 269–300. [Google Scholar]
- Chang, S.M.; Sung, J.M. Deteriorative changes in primed sweet corn seeds during storage. Seed Sci. Technol. 1998, 26, 613–625. [Google Scholar]
- Sahu, B.; Sahu, A.K.; Thomas, V.; Naithani, S.C. Reactive oxygen species, lipid peroxidation, protein oxidation and antioxidative enzymes in dehydrating Karanj (Pongamia pinnata) seeds during storage. South African J. Bot. 2017, 112, 383–390. [Google Scholar] [CrossRef]
- Adetunji, A.E.; Sershen; Varghese, B.; Pammenter, N.W. Effects of inorganic salt solutions on vigour, viability, oxidative metabolism and germination enzymes in aged cabbage and lettuce seeds. Plants 2020, 9, 1164. [Google Scholar] [CrossRef] [PubMed]
- Mira, S.; Estrelles, E.; González-Benito, M.E.; Corbineau, F. Biochemical changes induced in seeds of Brassicaceae wild species during ageing. Acta Physiol. Plant. 2011, 33, 1803–1809. [Google Scholar] [CrossRef]
- Adetunji, A.E.; Sershen; Varghese, B.; Pammenter, N. Effects of exogenous application of five antioxidants on vigour, viability, oxidative metabolism and germination enzymes in aged cabbage and lettuce seeds. South African J. Bot. 2021, 137, 85–97. [Google Scholar] [CrossRef]
- Coolbear, P.; Francis, A.; Grierson, D. The effect of low temperature pre-sowing treatment on the germination performance and membrane integrity of artificially aged tomato seeds. J. Exp. Bot. 1984, 35, 1609–1617. [Google Scholar] [CrossRef]
- Khan, M.M.; Iqbal, M.J.; Abbas, M.; Usman, M. Effect of ageing on viability, vigour and chromosomal damage in pea (Pisum sativum L.) seeds. Pakistan J. Agric. Sci. 2003, 40, 50–54. [Google Scholar]
- Berjak, P.; Villiers, T.A. Ageing in plant embryos. New Phytol. 1972, 71, 1069–1074. [Google Scholar] [CrossRef]
- Merritt, D.J.; Senaratna, T.; Touchell, D.H.; Dixon, K.W.; Sivasithamparam, K. Seed ageing of four Western Australian species in relation to storage environment and seed antioxidant activity. Seed Sci. Res. 2003, 13, 155–165. [Google Scholar] [CrossRef]
- Shaban, M. Review on physiological aspects of seed deterioration. Int. J. Agric. Crop Sci. 2013, 6, 627–631. [Google Scholar]
- Yan, H.-F.; Mao, P.-S.; Sun, Y.; Li, M.-L. Impacts of ascorbic acid on germination, antioxidant enzymes and ultrastructure of embryo cells of aged Elymus sibiricus seeds with different moisture contents. Int. J. Agric. Biol. 2016, 18, 176–183. [Google Scholar] [CrossRef]
- Draganić, I.; Lekić, S. Seed priming with antioxidants improves sunflower seed germination and seedling growth under unfavorable germination conditions. Turkish J. Agric. For. 2012, 36, 421–428. [Google Scholar] [CrossRef]
- Xia, F.; Cheng, H.; Chen, L.; Zhu, H.; Mao, P.; Wang, M. Influence of exogenous ascorbic acid and glutathione priming on mitochondrial structural and functional systems to alleviate aging damage in oat seeds. BMC Plant Biol. 2020, 20, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabarti, M.C.; Le, N.; Paik, C.H.; De Graff, W.G.; Carrasquillo, J.A. Prevention of radiolysis of monoclonal antibody during labeling. J. Nucl. Med. 1996, 37, 1384–1388. [Google Scholar]
- Yatim, R.M.; Kannan, T.P.; Ab Hamid, S.S. Effect of gamma radiation on the expression of mRNA growth factors in glycerol cryopreserved human amniotic membrane. Cell Tissue Bank. 2016, 17, 643–651. [Google Scholar] [CrossRef]
- Sershen; Berjak, P.; Pammenter, N.W.; Wesley-Smith, J. The effects of various parameters during processing for cryopreservation on the ultrastructure and viability of recalcitrant zygotic embryos of Amaryllis belladonna. Protoplasma 2012, 249, 155–169. [Google Scholar] [CrossRef]
- Roopa, K.S.; Geetha, N.P.; Sharathchandra, R.G.; Pushpalatha, H.G.; Sudisha, J.; Amruthesh, K.N.; Prakash, H.S.; Shetty, H.S. Osmopriming enhances pearl millet growth and induces downy mildew disease resistance. Arch. Phytopathol. Plant Prot. 2009, 42, 979–987. [Google Scholar] [CrossRef]
- Malik, S.; Ashraf, M. Exogenous application of ascorbic acid stimulates growth and photosynthesis of wheat (Triticum aestivum L.) under drought. Soil Environ. 2012, 31, 72–77. [Google Scholar]
- Shah, T.; Latif, S.; Khan, H.; Munsif, F.; Nie, L. Ascorbic acid priming enhances seed germination and seedling growth of winter wheat under low temperature due to late sowing in Pakistan. Agronomy 2019, 9, 757. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Barsa, S.M.A.; Wahid, A. Priming of field-sown rice seed enhances germination, seedling establishment, allometry and yield. Plant Growth Regul. 2006, 49, 285–294. [Google Scholar] [CrossRef]
- Ahmad, I.; Basra, S.M.A.; Wahid, A. Exogenous application of ascorbic acid, salicylic acid and hydrogen peroxide improves the productivity of hybrid maize at low temperature stress. Int. J. Agric. Biol. 2014, 16, 825–830. [Google Scholar]
- Luera, P.; Wahl-Villarreal, K.; Christoffersen, B.O.; Treviño, A.; Soti, P.; Gabler, C.A. Article effects of scarification, phytohormones, soil type, and warming on the germination and/or seedling performance of three tamaulipan thornscrub forest species. Plants 2021, 10, 1489. [Google Scholar] [CrossRef] [PubMed]
- Komba, C.G.; Brunton, B.J.; Hampton, J.G. Accelerated ageing vigour testing of kale (Brassica oleracea L. var. acephala DC) seed. Seed Sci. Technol. 2006, 34, 205–208. [Google Scholar] [CrossRef]
- TeKrony, D.M. Accelerated aging test: Principles and procedures. Seed Technol. 2005, 27, 135–146. [Google Scholar]
- Winston, P.W.; Bates, D.H. Saturated solutions for the control of humidity in biological research. Ecology 1960, 41, 232–237. [Google Scholar] [CrossRef]
- Ellis, R.H.; Roberts, E.H. Improved equations for the prediction of seed longevity. Ann. Bot. 1980, 45, 13–30. [Google Scholar] [CrossRef]
- Bam, R.; Hong, T.; Ellis, R.; Kumaga, F.; Asiedu, E. Storage behaviour of two contrasting upland rice genotypes. Ghana J. Agric. Sci. 2008, 41, 113–120. [Google Scholar] [CrossRef]
- Crawford, A.D.; Plummer, J.A.; Probert, R.J.; Steadman, K.J. The influence of cone age on the relative longevity of Banksia seeds. Ann. Bot. 2011, 107, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Patil, H.Y.; Mutanal, S.M.; Mokashi, M.V.; Ghatanatti, S.M. Germination and vigor index of different sources of Pongamia pinnata (L.) Pierre. Int. J. For. Crop Improv. 2017, 8, 8–11. [Google Scholar] [CrossRef]
- Mahajan, G.; Sarlach, R.S.; Japinder, S.; Gill, M.S. Seed priming effects on germination, growth and yield of dry direct-seeded rice. J. Crop Improv. 2011, 25, 409–417. [Google Scholar] [CrossRef]
- Patil, V.M.P.; Shivanna, H.; Surendra, P.; Manjunath, G.O.; Krishna, A.; Dasar, G. V Variability studies for seed and seedling traits in Pongamia pinnata (L.) Pierre. Karnataka J. Agric. Sci 2011, 24, 201–203. [Google Scholar]
- Soltani, E.; Ghaderi-Far, F.; Baskin, C.C.; Baskin, J.M. Problems with using mean germination time to calculate rate of seed germination. Aust. J. Bot. 2015, 63, 631–635. [Google Scholar] [CrossRef]
- Hussain, S.; Khaliq, A.; Matloob, A.; Wahid, M.A.; Afzal, I. Germination and growth response of three wheat cultivars to NaCl salinity. Soil Environ. 2013, 32, 36–43. [Google Scholar]
- Jamil, M.; Rehman, S.; Rha, E.S. Salinity effect on plant growth, PSII photochemistry and chlorophyll content in sugar beet (Beta vulgaris L.) and cabbage (Brassica oleracea capitata L.). Pakistan J. Bot. 2007, 39, 753–760. [Google Scholar]
- Abdul-Baki, A.A.; Anderson, J.D. Vigor determination in soybean seed by multiple criteria. Crop Sci. 1973, 13, 630–633. [Google Scholar] [CrossRef]
- Kitajima, M.; Butler, W.L. Quenching of chlorophyll fluorescence and primary photochemistry in chloroplasts by dibromothymoquinone. Biochim. Biophys. Acta 1975, 376, 105–115. [Google Scholar] [CrossRef]
- Sayed, O.H. Chlorophyll fluorescence as a tool in cereal crop research. Photosynthetica 2003, 41, 321–330. [Google Scholar] [CrossRef]
- Moradi, F.; Ismail, A.M. Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging systems to salt stress during seedling and reproductive stages in rice. Ann. Bot. 2007, 99, 1161–1173. [Google Scholar] [CrossRef] [Green Version]
- Probert, R.J.; Daws, M.I.; Hay, F.R. Ecological correlates of ex situ seed longevity: A comparative study on 195 species. Ann. Bot. 2009, 104, 57–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.-S.; Jeon, Y.-A.; Lee, Y.-Y.; Lee, S.-Y.; Kim, Y.-G. Comparison of seed viability among 42 species stored in a genebank. Korean J. Crop Sci. 2013, 58, 432–438. [Google Scholar] [CrossRef]
- Xu, L.; Xin, X.; Yin, G.; Zhou, J.; Zhou, Y.; Lu, X. Timing for antioxidant-priming against rice seed ageing: Optimal only in non-resistant stage. Sci. Rep. 2020, 10, 13294. [Google Scholar] [CrossRef]
- Ellis, R.H.; Nasehzadeh, M.; Hanson, J.; Woldemariam, Y. Medium-term seed storage of 50 genera of forage legumes and evidence-based genebank monitoring intervals. Genet. Resour. Crop Evol. 2018, 65, 607–623. [Google Scholar] [CrossRef] [Green Version]
- Walters, C.; Fleming, M.B.; Hill, L.M.; Dorr, E.J.; Richards, C.M. Stress–response relationships related to ageing and death of orthodox seeds: A study comparing viability and RNA integrity in soya bean (Glycine max) cv. Williams 82. Seed Sci. Res. 2020, 30, 161–172. [Google Scholar] [CrossRef]
- Butler, L.H.; Hay, F.R.; Ellis, R.H.; Smith, R.D.; Murray, T.B. Priming and re-drying improve the survival of mature seeds of Digitalis purpurea during storage. Ann. Bot. 2009, 103, 1261–1270. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.; TeKrony, D.M.; Egli, D.B.; Cornelius, P.L.; Rucker, M. Survival characteristics of corn seed during storage: I. Normal distribution of seed survival. Crop Sci. 1999, 39, 1394–1400. [Google Scholar] [CrossRef]
- Jatoi, S.A.; Afzal, M.; Nasim, S.; Anwar, R. Seed deterioration study in pea, using accelerated ageing techniques. Pakistan J. Biol. Sci. 2001, 4, 1490–1494. [Google Scholar] [CrossRef] [Green Version]
- Walters, C.; Wheeler, L.M.; Grotenhuis, J.M. Longevity of seeds stored in a genebank: Species characteristics. Seed Sci. Res. 2005, 15, 1–20. [Google Scholar] [CrossRef]
- Saxena, O.P.; Singh, G.; Pakeeraiah, T.; Pandey, N. Seed deterioration studies in some vegetable seeds. Acta Hortic. 1987, 215, 39–44. [Google Scholar] [CrossRef]
- Kochanek, J.; Buckley, Y.M.; Probert, R.J.; Adkins, S.W.; Steadman, K.J. Pre-zygotic parental environment modulates seed longevity. Austral Ecol. 2010, 35, 837–848. [Google Scholar] [CrossRef]
- Hay, F.; Klin, J.; Probert, R. Can a post-harvest ripening treatment extend the longevity of Rhododendron L. seeds? Sci. Hortic. 2006, 111, 80–83. [Google Scholar] [CrossRef]
- Roberts, E.H.; Ellis, R.H. Water and seed survival. Ann. Bot. 1989, 63, 39. [Google Scholar] [CrossRef]
- Walters, C. Understanding the mechanisms and kinetics of seed aging. Seed Sci. Res. 1998, 8, 223–244. [Google Scholar] [CrossRef]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. Moisture content and the longevity of seeds of Phaseolus vulgaris. Ann. Bot. 1990, 66, 341–348. [Google Scholar] [CrossRef]
- Copeland, L.O.; McDonald, M.B. Seed longevity and deterioration. In Principles of Seed Science and Technology; Springer: Boston, MA, USA, 1999; pp. 181–220. [Google Scholar]
- Simon, E.W. Phospholipids and plant membrane permeability. New Phytol. 1974, 73, 377–420. [Google Scholar] [CrossRef]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. Seed moisture content, storage, viability and vigour. Seed Sci. Res. 1991, 1, 275–279. [Google Scholar] [CrossRef] [Green Version]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. Survival and vigour of lettuce (Lactuca sativa L.) and sunflower (Helianthus annuus L.) seeds stored at low and very-low moisture contents. Ann. Bot. 1995, 76, 521–534. [Google Scholar] [CrossRef]
- Daws, M.I.; Lydall, E.; Chmielarz, P.; Leprince, O.; Matthews, S.; Thanos, C.A.; Pritchard, H.W. Developmental heat sum influences recalcitrant seed traits in Aesculus hippocastanum across Europe. New Phytol. 2004, 162, 157–166. [Google Scholar] [CrossRef]
- Wang, Y.; Mu, C.; Hou, Y.; Li, X. Optimum harvest time of Vicia cracca in relation to high seed quality during pod development. Crop Sci. 2008, 48, 709–715. [Google Scholar] [CrossRef]
- TeKrony, D.M. Precision is an essential component in seed vigour testing. Seed Sci. Technol. 2003, 31, 435–447. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.A.; Tabassum, R.; Afzal, I. Enhancing the performance of direct seeded fine rice by seed priming. Plant Prod. Sci. 2006, 9, 446–456. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, I.; Khaliq, T.; Ahmad, A.; Basra, S.M.A.; Hasnain, Z.; Ali, A. Effect of seed priming with ascorbic acid, salicylic acid and hydrogen peroxide on emergence, vigor and antioxidant activities of maize. African J. Biotechnol. 2012, 11, 1127–1132. [Google Scholar] [CrossRef]
- Yousof, F.I.; Mersal, I.F.; El-Emam, A.A.M. Effect of soaking rice (Oryza sativa, L.) seed in some anitioxidants solutions on germination and seedling vigor under different salinity levels. J. Plant Prod. 2010, 1, 279–290. [Google Scholar] [CrossRef]
- Brocklehurst, P.A.; Dearman, J. A comparison of different chemicals for osmotic treatment of vegetable seed. Ann. Appl. Biol. 1984, 105, 391–398. [Google Scholar] [CrossRef]
- Dolatabadian, A.; Ali, S.; Modarres, M. Effect of the ascorbic acid, pyridoxine and hydrogen peroxide treatments on germination, catalase activity, protein and malondialdehyde content of three oil seeds. Not. Bot. Horti Agrobot. Cluj-Napoca 2008, 36, 61–66. [Google Scholar] [CrossRef]
- Weraduwage, S.M.; Chen, J.; Anozie, F.C.; Morales, A.; Weise, S.E.; Sharkey, T.D. The relationship between leaf area growth and biomass accumulation in Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gifford, R.M.; Thorne, J.H.; Hitz, W.D.; Giaquinta, R.T. Crop productivity and photoassimilate partitioning. Science (801-–808). 1984, 225, 801–808. [Google Scholar] [CrossRef]
- Koester, R.P.; Skoneczka, J.A.; Cary, T.R.; Diers, B.W.; Ainsworth, E.A. Historical gains in soybean (Glycine max Merr.) seed yield are driven by linear increases in light interception, energy conversion, and partitioning efficiencies. J. Exp. Bot. 2014, 65, 3311–3321. [Google Scholar] [CrossRef]
- Vu, J.C.V.; Niedz, R.P.; Yelenosky, G. Glycerol stimulation of chlorophyll synthesis, embryogenesis, and carboxylation and sucrose metabolism enzymes in nucellar callus of “Hamlin” sweet orange. Plant Cell. Tissue Organ Cult. 1993, 33, 75–80. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Xu, Y.-F.; Ookawa, T.; Ishihara, K. Analysis of the photosynthetic characteristics of the high-yielding rice cultivar Takanari. Japanese J. Crop Sci. 1997, 66, 616–623. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, A.J.; Davies, W.J. The coupled response of stomatal conductance to photosynthesis and transpiration. J. Exp. Bot. 1998, 49, 399–406. [Google Scholar] [CrossRef]
- Miyashita, K.; Tanakamaru, S.; Maitani, T.; Kimura, K. Recovery responses of photosynthesis, transpiration, and stomatal conductance in kidney bean following drought stress. Environ. Exp. Bot. 2005, 53, 205–214. [Google Scholar] [CrossRef]
- Hayat, S.; Yadav, S.; Wani, A.S.; Irfan, M.; Ahmad, A. Nitric oxide effects on photosynthetic rate, growth, and antioxidant activity in tomato. Int. J. Veg. Sci. 2011, 17, 333–348. [Google Scholar] [CrossRef]
- Kusumi, K.; Hirotsuka, S.; Kumamaru, T.; Iba, K. Increased leaf photosynthesis caused by elevated stomatal conductance in a rice mutant deficient in SLAC1, a guard cell anion channel protein. J. Exp. Bot. 2012, 63, 5635–5644. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.; Chen, K. The regulatory role of silicon on water relations, photosynthetic gas exchange, and carboxylation activities of wheat leaves in field drought conditions. Acta Physiol. Plant. 2012, 34, 1589–1594. [Google Scholar] [CrossRef]
- Kumutha, D.; Ezhilmathi, K.; Sairam, R.K.; Srivastava, G.C.; Deshmukh, P.S.; Meena, R.C. Waterlogging induced oxidative stress and antioxidant activity in pigeonpea genotypes. Biol. Plant. 2009, 53, 75–84. [Google Scholar] [CrossRef]
- Miyake, C.; Asada, K. Thylakoid-bound ascorbate peroxidase in spinach chloroplasts and photoreduction of its primary oxidation product monodehydroascorbate radicals in thylakoids. Plant Cell Physiol. 1992, 33, 541–553. [Google Scholar] [CrossRef]
- Zhang, S.; Weng, J.; Pan, J.; Tu, T.; Yao, S.; Xu, C. Study on the photo-generation of superoxide radicals in Photosystem II with EPR spin trapping techniques. Photosynth. Res. 2003, 75, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Dolatabadian, A.; Modarres Sanavy, S.A.M.; Sharifi, M. Alleviation of water deficit stress effects by foliar application of ascorbic acid on Zea mays L. J. Agron. Crop Sci. 2009, 195, 347–355. [Google Scholar] [CrossRef]
- Zhao, H.J.; Zou, Q. Protective Effects of exogenous antioxidants and phenolic compounds on photosynthesis of wheat leaves under high irradiance and oxidative stress. Photosynthetica 2002, 40, 523–527. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox sensing and signalling associated with reactive oxygen in chloroplasts, peroxisomes and mitochondria. Physiol. Plant. 2003, 119, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Shigeoka, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CD Level | Cabbage Seed Deterioration Rate (Probit/Day) | Lettuce Seed Deterioration Rate (Probit/Day) |
|---|---|---|
| P75 | 0.13 ± 0.01 b | 0.47 ± 0.02 a |
| P50 | 0.15 ± 0.01 b | 0.47 ± 0.02 a |
| P25 | 0.15 ± 0.01 b | 0.48 ± 0.02 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adetunji, A.E.; Sershen; Varghese, B.; Pammenter, N.W. Exogenous Antioxidants Enhance Seedling Growth and Yield of Artificially Aged Cabbage and Lettuce Seeds. Horticulturae 2021, 7, 274. https://doi.org/10.3390/horticulturae7090274
Adetunji AE, Sershen, Varghese B, Pammenter NW. Exogenous Antioxidants Enhance Seedling Growth and Yield of Artificially Aged Cabbage and Lettuce Seeds. Horticulturae. 2021; 7(9):274. https://doi.org/10.3390/horticulturae7090274
Chicago/Turabian StyleAdetunji, Ademola Emmanuel, Sershen, Boby Varghese, and Norman W. Pammenter. 2021. "Exogenous Antioxidants Enhance Seedling Growth and Yield of Artificially Aged Cabbage and Lettuce Seeds" Horticulturae 7, no. 9: 274. https://doi.org/10.3390/horticulturae7090274
APA StyleAdetunji, A. E., Sershen, Varghese, B., & Pammenter, N. W. (2021). Exogenous Antioxidants Enhance Seedling Growth and Yield of Artificially Aged Cabbage and Lettuce Seeds. Horticulturae, 7(9), 274. https://doi.org/10.3390/horticulturae7090274