
HPLC-DAD-APCI-MS as a Tool for Carotenoid Assessment of Wild and Cultivated Cherry Tomatoes

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
2.2. Samples and Reagents
2.3. Carotenoid Conventional Extraction Method
2.4. HPLC-DAD-MS Analysis
2.5. Statistical Analysis
3. Results
3.1. Carotenoid Characterization by HPLC-DAD-MS
3.2. Relationship between Carotenoid Levels and Tomato Accessions
4. Discussion
4.1. Carotenoid Characterization by HPLC-DAD-MS
4.2. Relationship between Carotenoid Levels and Tomato Accessions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2014, 8, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Gong, M.; Bassi, A. Carotenoids from microalgae: A review of recent developments. Biotechnol. Adv. 2016, 34, 1396–1412. [Google Scholar] [CrossRef]
- Fiedor, J.; Burda, K. Potential role of carotenoids as antioxidants in human health and disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, pharmacology and treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef] [Green Version]
- Saini, R.K.; Keum, Y.-S. Carotenoid extraction methods: A review of recent developments. Food Chem. 2018, 240, 90–103. [Google Scholar] [CrossRef]
- Poojary, M.M.; Barba, F.J.; Aliakbarian, B.; Donsì, F.; Pataro, G.; Dias, D.A.; Juliano, P. Innovative alternative technologies to extract carotenoids from microalgae and seaweeds. Mar. Drugs 2016, 14, 214. [Google Scholar] [CrossRef]
- Shi, J. Lycopene in tomatoes: Chemical and physical properties affected by food processing. Crit. Rev. Biotechnol. 2000, 20, 293–334. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, D.; Ferruzzi, M.G. Identification and quantification of carotenoids and tocochromanols in sorghum grain by high-performance liquid chromatography. In Sorghum; Zhao, Z.-Y., Dahlberg, J., Eds.; Humana Press: New York, NY, USA, 2019; pp. 141–151. [Google Scholar]
- Sen, S. The chemistry and biology of lycopene: Antioxidant for human health. Int. J. Adv. Life Sci. Res. 2019, 2, 8–14. [Google Scholar] [CrossRef]
- Mendelová, A.; Fikselová, M.; Mendel, L. Carotenoids and lycopene content in fresh and dried tomato fruits and tomato juice. Acta Univ. Agric. Silvic. Mendel. Brun. 2013, 61, 1329–1337. [Google Scholar] [CrossRef] [Green Version]
- Sahu, K.K.; Chattopadhyay, D. Genome-wide sequence variations between wild and cultivated tomato species revisited by whole genome sequence mapping. BMC Genom. 2017, 18, 430. [Google Scholar] [CrossRef] [Green Version]
- Lado, J.; Zacarias, L.; Rodrigo, M.J. Regulation of Carotenoid biosynthesis during fruit development. Carotenoids Nat. 2016, 79, 161–198. [Google Scholar] [CrossRef]
- Mariutti, L.R.; Mercadante, A.Z. Carotenoid Esters in Foods: Physical, Chemical and Biological Properties (Carotenoid Esters in Foods); Royal Society of Chemistry: London, UK, 2019. [Google Scholar]
- Londoño-Giraldo, L.; Baena-Pedroza, A.; Martinez-Seidel, F.; Corpas-Iguarán, E.; Taborda-Ocampo, G. Gone wild: Integration of antioxidative, physicochemical, volatilomic and sensorial profiles ratify rustic relatives of cherry tomato as ideal mating partners. Sci. Hortic. 2020, 277, 109814. [Google Scholar] [CrossRef]
- Londoño-Giraldo, L.M.; Gonzalez, J.; Baena, A.M.; Tapasco, O.; Corpas, E.J.; Taborda, G. Selection of promissory crops of wild cherry-type tomatoes using physicochemical parameters and antioxidant contents. Bragantia 2020, 79, 169–179. [Google Scholar] [CrossRef]
- Olives Barba, A.I.; Camara Hurtado, M.; Sanchez Mata, M.C.; Fernandez Ruiz, V.; Lopez Saenz de Tejada, M. Application of a UV-vis detection-HPLC method for a rapid determination of lycopene and b-carotene in vegetables. Food Chem. 2006, 95, 328–336. [Google Scholar] [CrossRef]
- Bueno, M.; Vitali, C.; Martínez, J.D.S.; Mendiola, J.A.; Cifuentes, A.; Ibañez, E.; Herrero, M. Compressed CO2 technologies for the recovery of carotenoid-enriched extracts from Dunaliella salina with potential neuroprotective activity. ACS Sustain. Chem. Eng. 2020, 8. [Google Scholar] [CrossRef]
- Gallego, R.; Martínez, M.; Cifuentes, A.; Ibáñez, E.; Herrero, M. Development of a green downstream process for the valorization of Porphyridium cruentum biomass. Molecules 2019, 24, 1564. [Google Scholar] [CrossRef] [Green Version]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. Carotenoids; Springer: Berlin, Germany, 2004. [Google Scholar]
- van Breemen, R.B.; Dong, L.; Pajkovic, N.D. Atmospheric pressure chemical ionization tandem mass spectrometry of carotenoids. Int. J. Mass Spectrom. 2012, 312, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Sreelakshmi, Y.; Sharma, R. A rapid and sensitive method for determination of carotenoids in plant tissues by high performance liquid chromatography. Plant Methods 2015, 11, 5. [Google Scholar] [CrossRef] [Green Version]
- Mercadante, A.; Rodrigues, D.; Petry, F.; Barros Mariutti, L. Carotenoid esters in foods—A review and practical directions on analysis and occurrence. Food Res. Int. 2017, 99, 830–850. [Google Scholar] [CrossRef]
- Dzakovich, M.; Gas-Pascual, E.; Orchard, C.J.; Sari, E.N.; Riedl, K.; Schwartz, S.J.; Francis, D.M.; Cooperstone, J.L. Analysis of tomato carotenoids: Comparing extraction and chromatographic methods. J. AOAC Int. 2019, 102, 1069–1079. [Google Scholar] [CrossRef]
- Ghislain, T.; Faure, P.; Michels, R. Detection and monitoring of PAH and oxy-PAHs by high resolution mass spectrometry: Comparison of ESI, APCI and APPI source detection. J. Am. Soc. Mass Spectrom. 2012, 23, 530–536. [Google Scholar] [CrossRef] [Green Version]
- Stommel, J.R.; Haynes, K.G. Inheritance of betacarotene content in the wild tomato species Lycopersicon cheesmanii. J. Hered. 1994, 85, 401–404. [Google Scholar] [CrossRef]
- Lewinsohn, E.; Sitrit, Y.; Bar, E.; Azulay, Y.; Ibdah, M.; Meir, A.; Yosef, E.; Zamir, D.; Tadmor, Y. Carotenoid pigmentation affects the volatile composition of tomato and watermelon fruits, as revealed by comparative genetic analyses. J. Agric. Food Chem. 2005, 53, 3142–3148. [Google Scholar] [CrossRef] [PubMed]
- Ciriminna, R.; Fidalgo, A.; Meneguzzo, F.; Ilharco, L.; Pagliaro, M. Lycopene: Emerging production methods and applications of a valued carotenoid. ACS Sustain. Chem. Eng. 2015, 4, 643–650. [Google Scholar] [CrossRef]
- Liu, Y.; Sherry Roof, S.; Ye, Z.; Barry, C.; van Tuinen, A.; Vrebalov, J.; Bowler, C.; Giovannoni, J. Manipulation of light signal transduction as a means of modifying fruit nutritional quality in tomato. Proc. Natl. Acad. Sci. USA 2004, 101, 9897–9902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambla, J.L.; Tikunov, Y.M.; Monforte, A.J.; Bovy, A.G.; Granell, A. The expanded tomato fruit volatile landscape. J. Exp. Bot. 2014, 65, 4613–4623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronen, G.; Carmel-Goren, L.; Zamir, D.; Hirschberg, J. An alternative pathway to β-carotene formation in plant chromoplasts discovered by map-based cloning of Beta and old-gold color mutations in tomato. Proc. Natl. Acad. Sci. USA 2000, 97, 11102–11107. [Google Scholar] [CrossRef] [Green Version]
- Fraser, P.D.; Truesdale, M.R.; Bird, C.R.; Schuch, W.; Bramley, P.M. Carotenoid biosynthesis during tomato fruit development. Plant Physiol. 1994, 105, 405–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronen, G.; Cohen, M.; Zamir, D.; Hirschberg, J. Regulation of carotenoid biosynthesis during tomato fruit development: Expression of the gene for lycopene epsilon-cyclase is down-regulated during ripening and is elevated in the mutant Delta. Plant J. 1999, 17, 341–351. [Google Scholar] [CrossRef]
- Sajilata, M.; Singhal, R.; Kamat, M. The Carotenoid pigment zeaxanthin—A review. Compr. Rev. Food Sci. Food Saf. 2008, 7, 29–49. [Google Scholar] [CrossRef]
- Ma, L.; Yan, S.-F.; Huang, Y.-M.; Lu, X.-R.; Qian, F.; Pang, H.-L.; Xu, X.-R.; Zou, Z.; Dong, P.-C.; Xiao, X.; et al. Effect of lutein and zeaxanthin on macular pigment and visual function in patients with early age-related macular degeneration. Ophthalmology 2012, 119, 2290–2297. [Google Scholar] [CrossRef] [PubMed]
- Giorio, G.; Yildirim, A.; Stigliani, A.L.; D’Ambrosio, C. Elevation of lutein content in tomato: A biochemical tug-of-war between lycopene cyclases. Metab. Eng. 2013, 20, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Lewinsohn, E.; Sitrit, Y.; Bar, E.; Azulay, Y.; Ibdah, M.; Meir, A.; Yosef, E.; Zamir, D.; Tadmor, Y. Not just colors—Carotenoid degradation as a link between pigmentation and aroma in tomato and watermelon fruit. Trends Food Sci. Technol. 2005, 16, 407–415. [Google Scholar] [CrossRef]
- Havaux, M. Carotenoids as membrane stabilizers in chloroplasts. Trends Plant. Sci. 1998, 3, 147–151. [Google Scholar] [CrossRef]
- Aizawa, K.; Liu, C.; Tang, S.; Veeramachaneni, S.; Hu, K.-Q.; Smith, D.E.; Wang, X.-D. Tobacco carcinogen induces both lung cancer and non-alcoholic steatohepatitis and hepatocellular carcinomas in ferrets which can be attenuated by lycopene supplementation. Int. J. Cancer 2016, 139, 1171–1181. [Google Scholar] [CrossRef]
- Rugkong, A.; McQuinn, R.; Giovannoni, J.J.; Rose, J.K.C.; Watkins, C.B. Expression of ripening-related genes in cold-stored tomato fruit. Postharvest Biol. Technol. 2011, 61, 1–14. [Google Scholar] [CrossRef]
- Tieman, D.; Zhu, G.; Resende, M.F.R., Jr.; Lin, T.; Nguyen, C.; Bies, D.; Rambla, J.L.; Beltran, K.S.O.; Taylor, M.; Zhang, B.; et al. A chemical genetic roadmap to improved tomato flavour. Science 2017, 355, 391–394. [Google Scholar] [CrossRef]
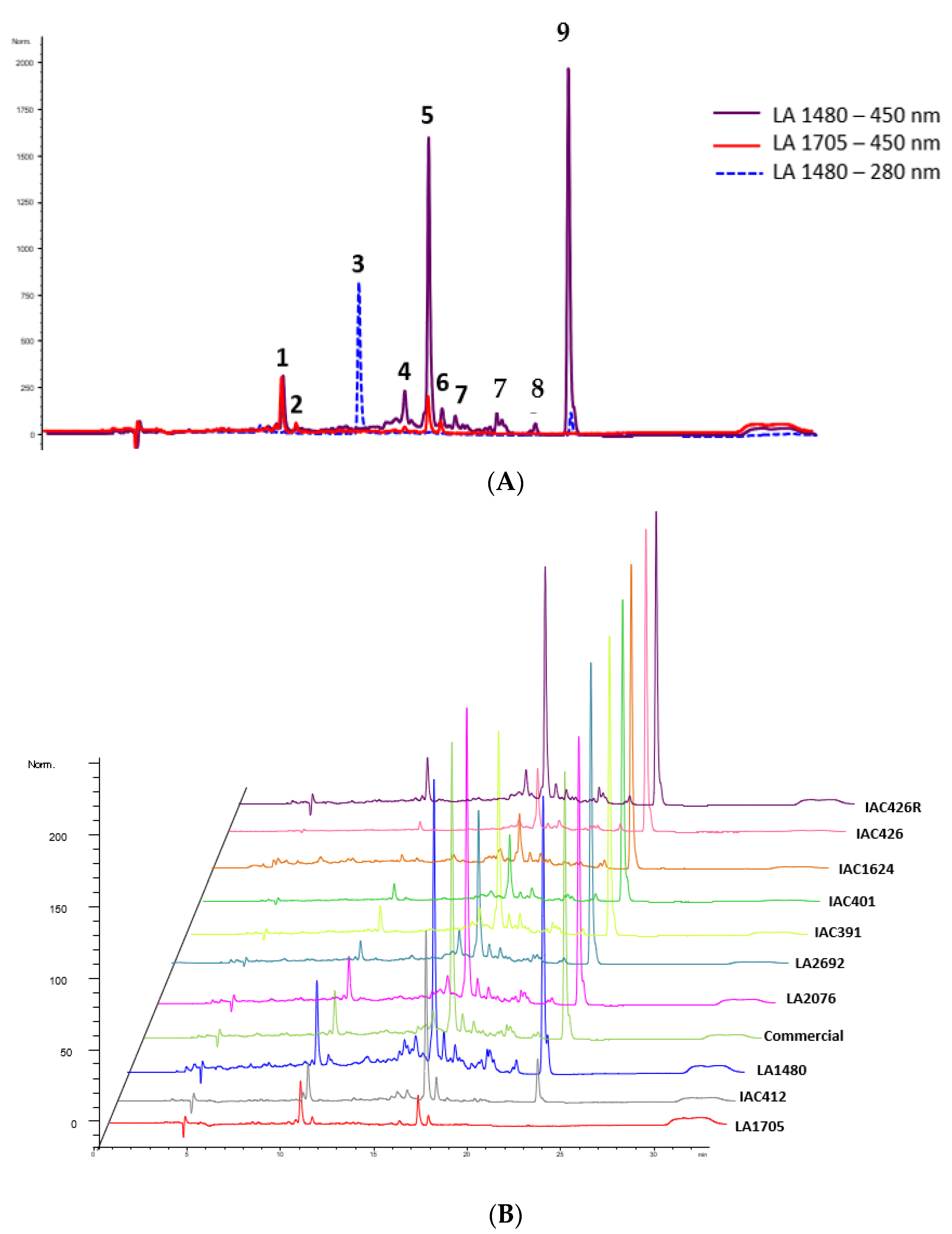


{kind=link}
{kind=link}
| Carotenoids | r2 | Linearity Range
(µg/mL) | LOD
(µg/mL) | LOQ
(µg/mL) |
|---|---|---|---|---|
| Lutein | 0.993 | 4.3–51.2 | 3.66 × 10−2 | 1.22 × 10−1 |
| Zeaxanthin | 0.995 | 2.3–61.2 | 5.18 × 10−3 | 1.72 × 10−2 |
| all-trans-β-carotene | 1.000 | 0.2–26.1 | 8.11 × 10−2 | 2.70 × 10−1 |
| Lycopene | 0.999 | 1.3–99.6 | 7.32 × 10−2 | 2.44 × 10−1 |
| Peak | Carotenoids | RT (min) | UV/Vis Max (nm) | [M+H]+ m/z |
|---|---|---|---|---|
| 1 | Lutein a | 10.11 | 445, 474 | 569.4 |
| 2 | Zeaxanthin a | 10.72 | 425, 449, 476 | 569.6 |
| 3 | phytoene | 11.68 | 281 | |
| 4 | all-trans-α-carotene | 15.36 | 424, 446, 475 | 537.4 |
| 5 | all-trans-β-carotene a | 16.36 | 452, 477 | 537.6 |
| 6 | carotenoid 1 | 16.90 | 426, 448, 475 | 537.5 |
| 7 | δ-carotene | 17.49 | 432, 457, 488 | 537.5 |
| 8 | 1,2-dihydrolycopene | 20.75 | 440, 468, 502 | 537.4 |
| 9 | trans-lycopene a | 22.16 | 446, 472, 503 | 537.4 |
| Accession | Luteine | α-Carotene | β-Carotene | δ-Carotene | Carotenoid 1 | Zeaxanthin | Lycopene | 1,2-Dihydrolycopene | Total Carotenoids |
|---|---|---|---|---|---|---|---|---|---|
| IAC426 | 1.79 ± 0.18 | 0.259 ± 0.036 a | 0.802 ± 0.090 a | 0.259 ± 0.036 a,b | 0.131 ± 0.011 a | trace | 3.80 ± 0.50 a | 65 ± 27 | 5.31 ± 0.60 a |
| IAC1624 | 1.81 ± 0.14 | 0.100 ± 0.040 e | 0.458 ± 0.022 b | 0.100 ± 0.040 d | 0.080 ± 0.004 b,c,d | trace | 0.80 ± 0.30 e,f | trace | 1.54 ± 0.30 d |
| LA1705 | 1.80 ± 0.10 | 0.001 ± 0.00 f | 0.061 ± 0.003 c | ND e | NDf | 34.3 ± 5.7 | ND g | trace | 0.09 ± 0.01 f |
| IAC401 | 1.72 ± 0.20 | 0.074 ± 0.003 c,d,e | 0.504 ± 0.08 b | 0.151 ± 0.011 d | 0.085 ± 0.013 b | trace | 2.00 ± 0.30 b,c | trace | 2.81 ± 0.40 c |
| IAC391 | 1.86 ± 0.06 | 0.190 ± 0.004 b,c | 0.887 ± 0.037 a | 0.122 ± 0.014 a,b,c | 0.119 ± 0.007 b,c | trace | 1.30 ± 0.00 c,d,e | trace | 2.61 ± 0.04 c |
| IAC412 | 1.94 ± 0.15 | 0.127 ± 0.004 d,e | 0.478 ± 0.047 b | 0.038 ± 0.003 b,c,d | 0.0105 ± 0.008 e,f | trace | 0.05 ± 0.01 f,g | trace | 0.79 ± 0.10 e,f |
| IAC426R | 1.76 ± 0.01 | 0.121 ± 0.00 d,e | 0.529 ± 0.001 b | 0.056 ± 0.001 d | 0.085 ± 0.001 d,e,f | trace | 0.70 ± 0.01 e,f,g | trace | 1.49 ± 0.02 d |
| LA1480 | 1.81 ± 0.05 | 0.280 ± 0.00 a | 0.493 ± 0.056 b | 0.142 ± 0.002 b,c,d | 0.107 ± 0.001 b | trace | 2.30 ± 0.09 b | trace | 3.32 ± 0.01 b |
| LA2076 | 2.13 ± 0.01 | 0.154 ± 0.002 c,d,e | 0.853 ± 0.00 a | 0.075 ± 0.00 b,c,d | 0.107 ± 0.00 c,d,e | trace | 0.90 ± 0.00 e | trace | 2.09 ± 0.01 c,d |
| LA2692 | 2.01 ± 0.12 | 0.247 ± 0.016 a,b | 0.860 ± 0.063 a | 0.1.12 ± 0.004 b | 0.144 ± 0.015 a | trace | 1.80 ± 0.20 b,c,d | trace | 3.16 ± 0.30 b |
| Commercial | 1.40 ± 0.24 | 0.164 ± 0.013 c,d | 0.384 ± 0.094 b | 0.077 ± 0.003 c,d | 0.095 ± 0.002 c,d,e | trace | 1.10 ± 0.10 d,e | trace | 1.82 ± 0.10 d |
| p-value | NS | *** | *** | *** | *** | *** | *** | ** | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Londoño-Giraldo, L.M.; Bueno, M.; Corpas-Iguarán, E.; Taborda-Ocampo, G.; Cifuentes, A. HPLC-DAD-APCI-MS as a Tool for Carotenoid Assessment of Wild and Cultivated Cherry Tomatoes. Horticulturae 2021, 7, 272. https://doi.org/10.3390/horticulturae7090272
Londoño-Giraldo LM, Bueno M, Corpas-Iguarán E, Taborda-Ocampo G, Cifuentes A. HPLC-DAD-APCI-MS as a Tool for Carotenoid Assessment of Wild and Cultivated Cherry Tomatoes. Horticulturae. 2021; 7(9):272. https://doi.org/10.3390/horticulturae7090272
Chicago/Turabian StyleLondoño-Giraldo, Lina M., Mónica Bueno, Eduardo Corpas-Iguarán, Gonzalo Taborda-Ocampo, and Alejandro Cifuentes. 2021. "HPLC-DAD-APCI-MS as a Tool for Carotenoid Assessment of Wild and Cultivated Cherry Tomatoes" Horticulturae 7, no. 9: 272. https://doi.org/10.3390/horticulturae7090272
APA StyleLondoño-Giraldo, L. M., Bueno, M., Corpas-Iguarán, E., Taborda-Ocampo, G., & Cifuentes, A. (2021). HPLC-DAD-APCI-MS as a Tool for Carotenoid Assessment of Wild and Cultivated Cherry Tomatoes. Horticulturae, 7(9), 272. https://doi.org/10.3390/horticulturae7090272