
Studies of Vegetative Growth, Inflorescence Development and Eco-Dormancy Formation of Abscission Layers in Streptocarpus formosus (Gesneriaceae)


Abstract
1. Introduction
2. Materials and Methods
2.1. Greenhouse Experiment
2.2. Plant Preparation
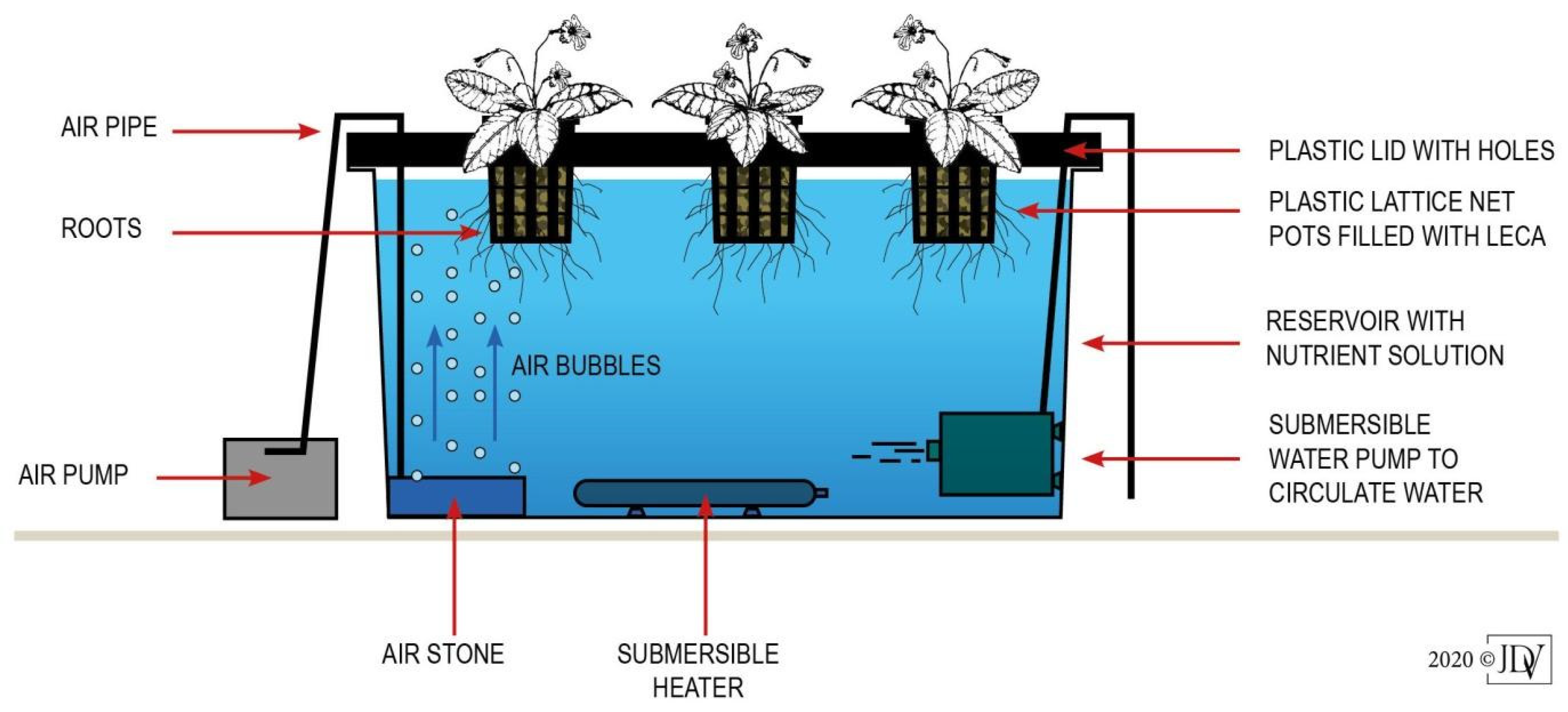
2.3. Hydroponic Cultivation
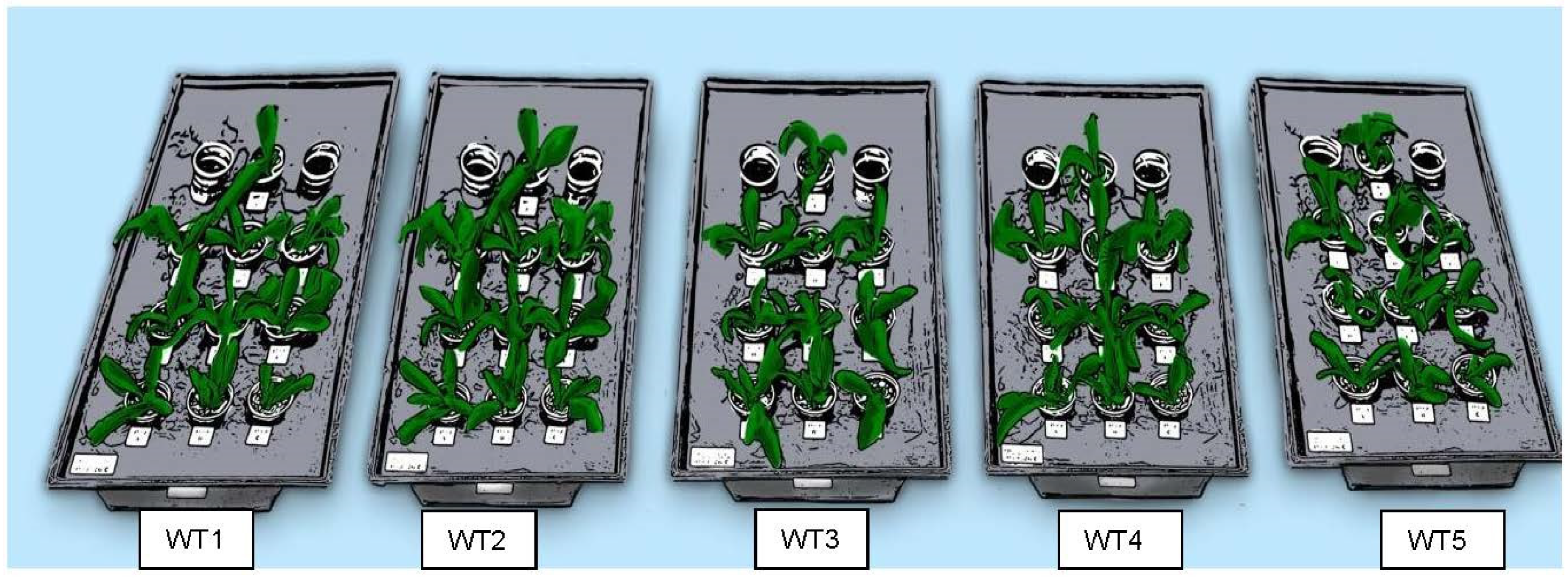
2.4. Water Temperature Treatments and Experimental Design
2.5. Vegetative Growth and Data Collection
2.6. Inflorescence Data Collection
2.7. Eco-Dormancy Data Collection
2.8. Statistical Analysis
3. Results
3.1. Total Leaf Number
3.2. Total Leaf Length
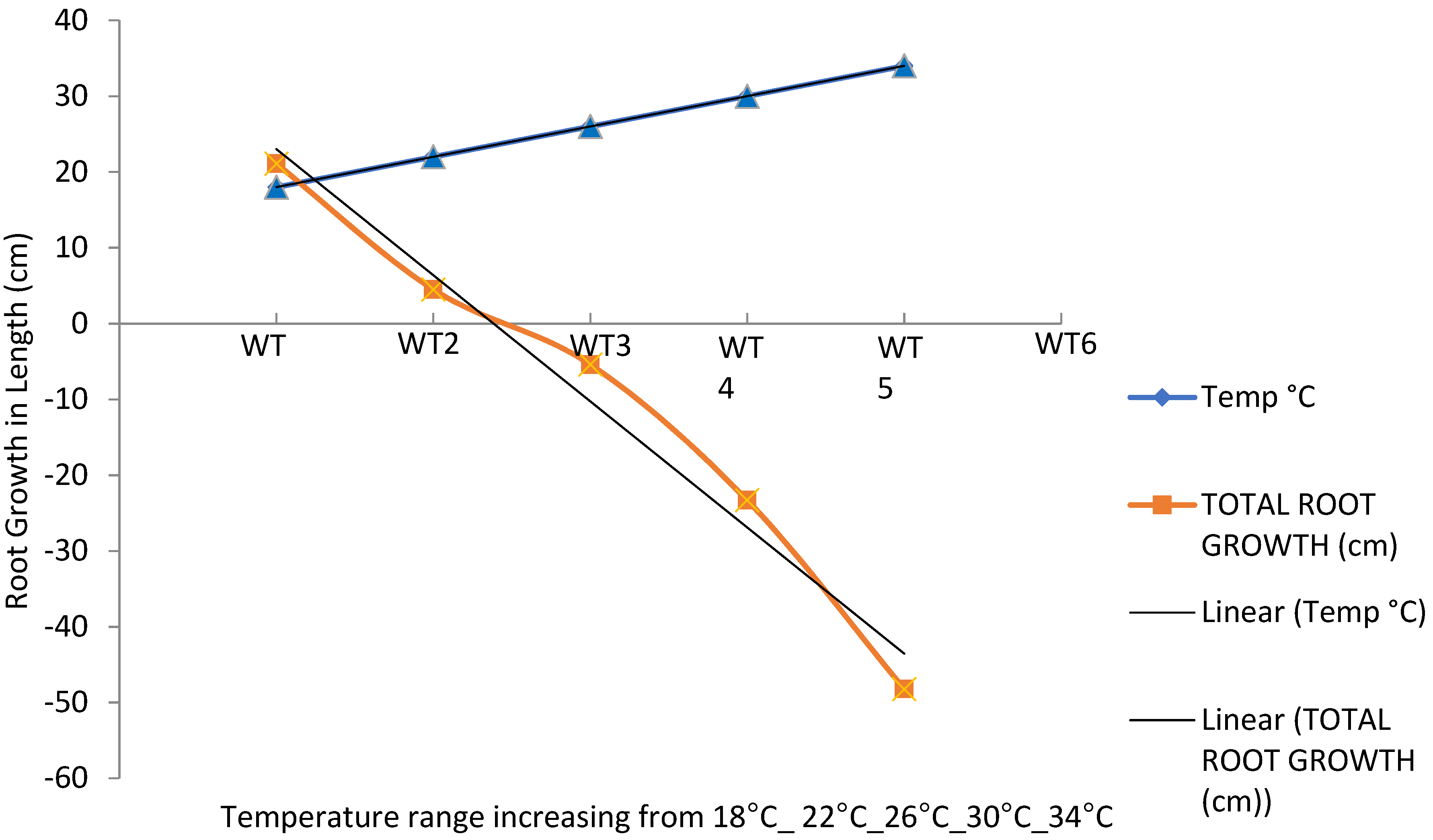
3.3. Total Root Growth
3.4. Total Fresh Weight
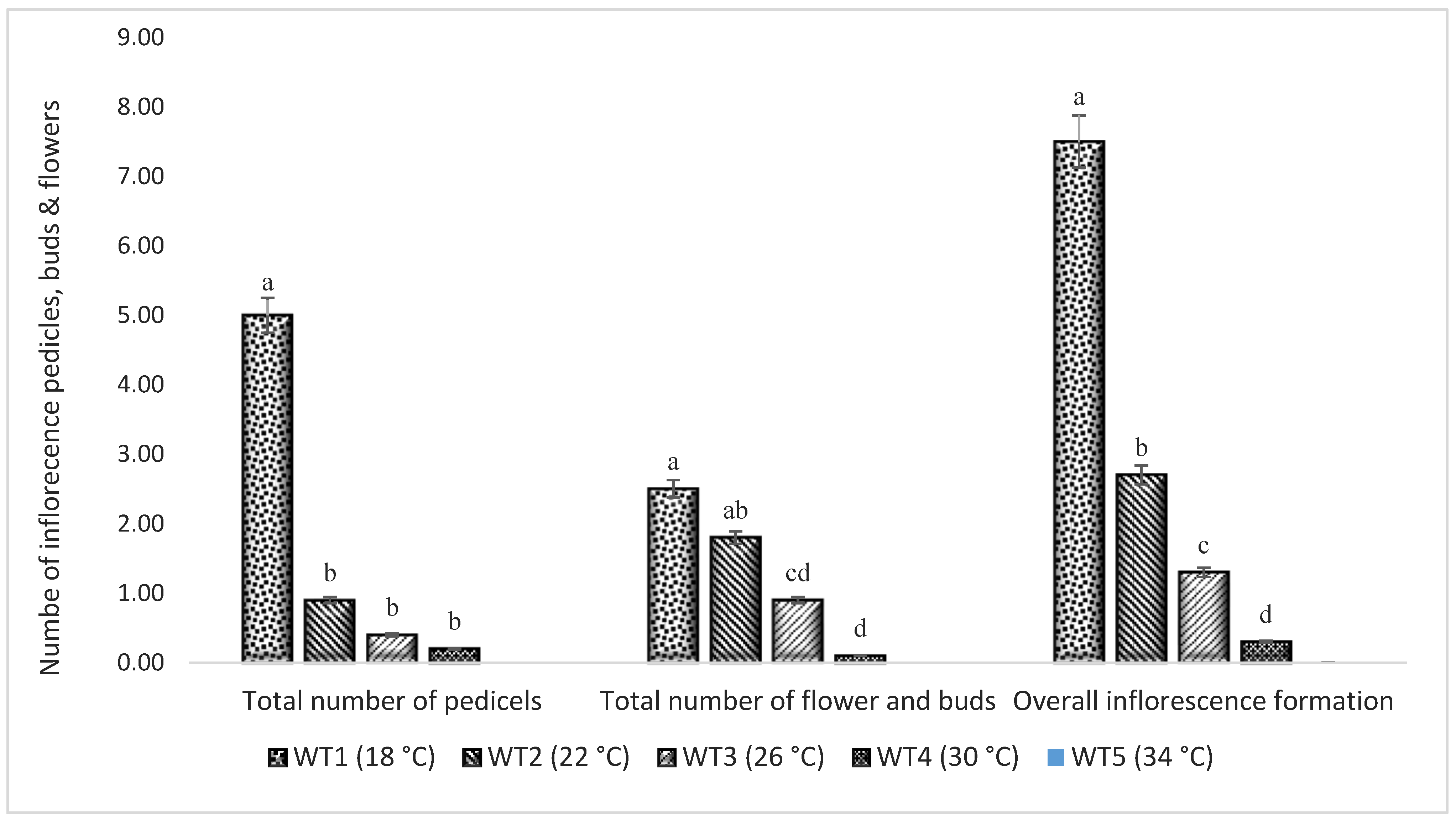
3.5. Flowering in Response to Five Different Temperature Regimes in Hydroponics
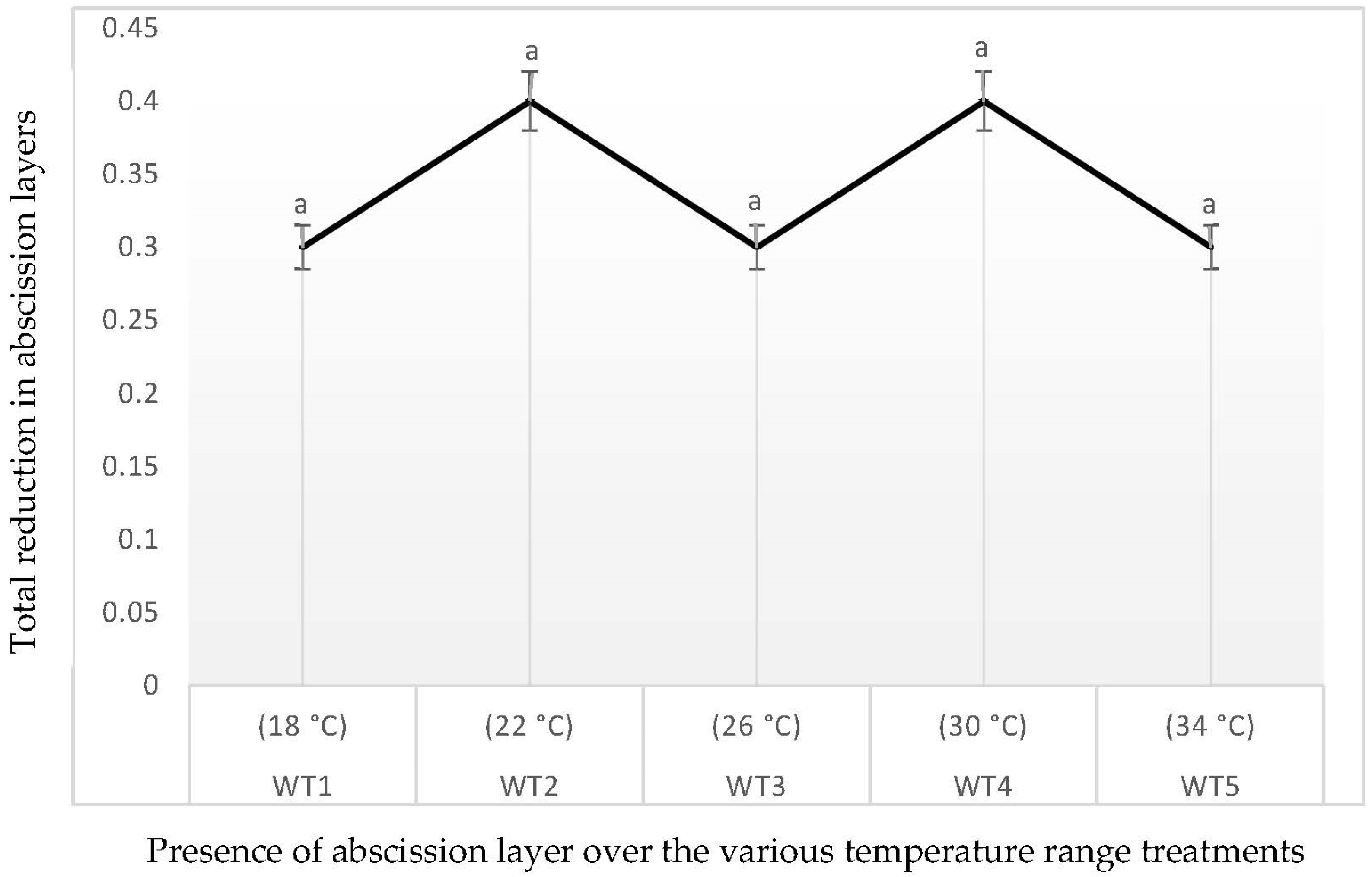
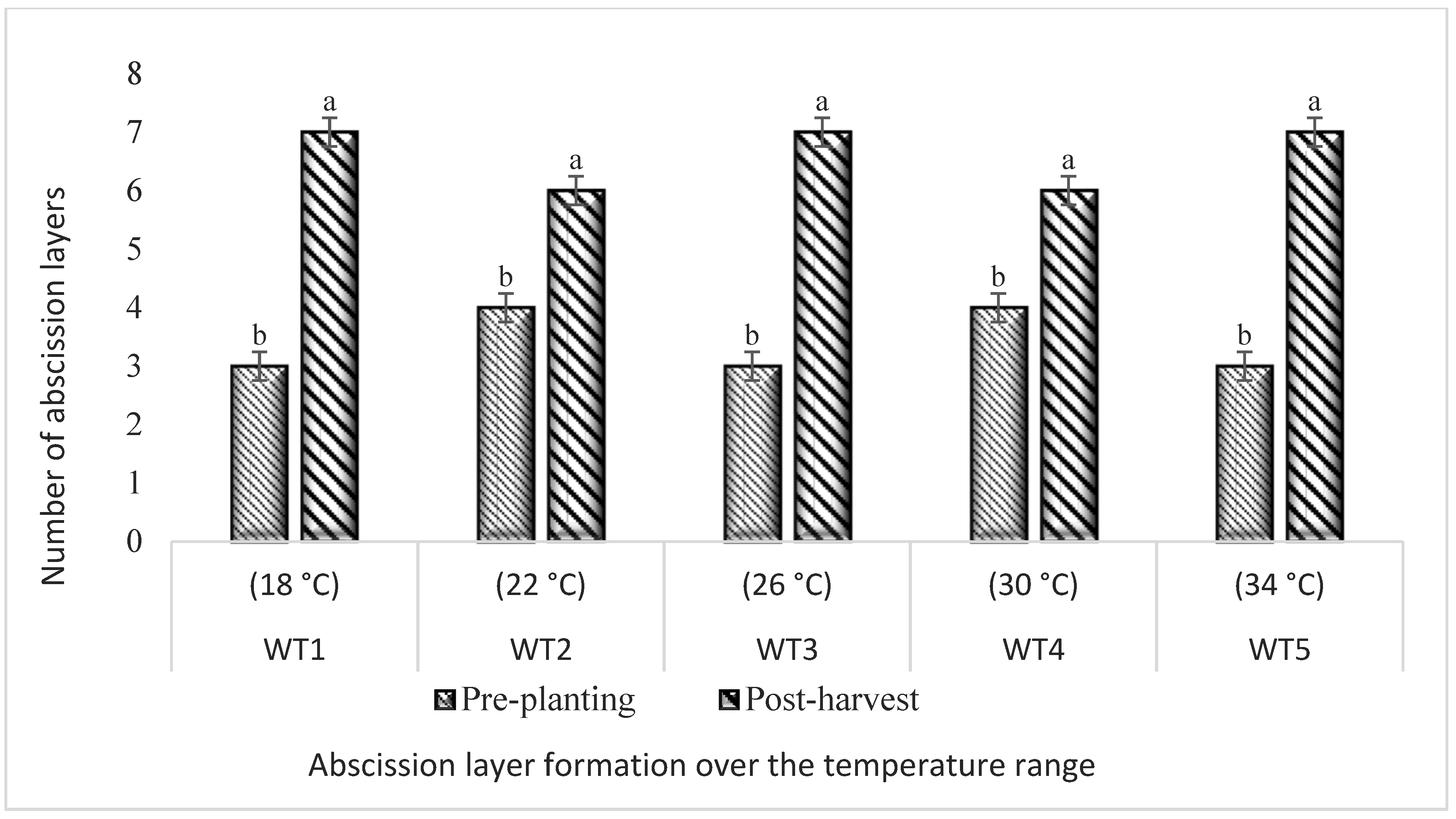
3.6. Reduction in Abscission Layers in Response to Five Different Temperature Regimes in Hydroponics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hillard, O.; Burtt, B. Streptocarpus, an African Plant Study; University of Natal Press: Pietermaritzburg, South Africa, 1971. [Google Scholar]
- Otieno, M.; Joshi, N.; Rutschmann, B. Flower visitors of Streptocarpus teitensis: Implications for conservation of a critically endangered African violet species in Kenya. PeerJ 2021, 9, 1–18. [Google Scholar] [CrossRef]
- van der Walt, L. Streptocarpus formosus. Veld Flora 2001, 87, 116–117. [Google Scholar]
- Pooley, E. A field Guide to Wild Flowers Kwazulu-Natal and the Eastern Region; Natal Flora Publications Trust: Durban, South Africa, 1998. [Google Scholar]
- Russo, S.E.; Kitajima, K. The Ecophysiology of Leaf Lifespan in Tropical Forests: Adaptive and Plastic Responses to Environmental Heterogeneity. In Tropical Tree Physiology; Goldstein, G., Santiago, L., Eds.; Springer: Cham, Switzerland, 2016; Volume 6, pp. 357–383. [Google Scholar]
- Van Doorn, W.G.; Woltering, E.J. Senescence and programmed cell death: Substance or semantics? J. Exp. Bot. 2004, 55, 2147–2153. [Google Scholar] [CrossRef] [PubMed]
- Lang, G.; Early, J.; Martin, G.; Darnell, R. Endo-, para-, and ecodormancy: Physiological terminology and classification for dormancy research. HortScience 1987, 22, 371–377. [Google Scholar]
- White, J.W. New and renewed pot plants- Streptocarpus. Pa. Flower Grow. Bull. 1975, 279, 469–472. [Google Scholar]
- Viljoen, C. Streptocarpus Saxorum/Plantz Africa; SANBI: Pretoria, South Africa, 2010. [Google Scholar]
- Blanchard, M.G.; Runkle, E.S. Intermittent light from a rotating high-pressure sodium lamp promotes flowering of long-day plants. HortScience 2010, 45, 236–241. [Google Scholar] [CrossRef]
- Nxawe, S.; Ndakidemi, P.A.; Laubscher, C.P. Possible effects of regulating hydroponic water temperature on plant growth, accumulation of nutrients and other metabolites. Afr. J. Biotechnol. 2010, 9, 9128–9134. [Google Scholar] [CrossRef]
- Sade, N.; Del Mar Rubio-Wilhelmi, M.; Umnajkitikorn, K.; Blumwald, E. Stress-induced senescence and plant tolerance to abiotic stress. J. Exp. Bot. 2018, 69, 845–853. [Google Scholar] [CrossRef]
- Mills, P.J.W.; Smith, I.E.; Marais, G. A greenhouse design for a cool subtropical climate with mild winters based on microclimatic measurements of protected environments. Acta Hortic. 1990, 281, 83–94. [Google Scholar] [CrossRef]
- Avila-Ospina, L.; Moison, M.; Yoshimoto, K.; Masclaux-Daubresse, C. Autophagy, plant senescence, and nutrient recycling. J. Exp. Bot. 2014, 65, 3799–3811. [Google Scholar] [CrossRef]
- Yan, Q.-Y.; Duan, Z.-Q.; Mao, J.-D.; Li, X.; Dong, F. Low Root Zone Temperature Limits Nutrient Effects on Cucumber Seedling Growth and Induces Adversity Physiological Response. J. Integr. Agric. 2013, 12, 1450–1460. [Google Scholar] [CrossRef]
- Al-Rawahy, M.S.; Al-Rawahy, S.A.; Al-Mulla, Y.A.; Nadaf, S.K. Effect of Cooling Root-Zone Temperature on Growth, Yield and Nutrient Uptake in Cucumber Grown in Hydroponic System During Summer Season in Cooled Greenhouse. J. Agric. Sci. 2019, 11. [Google Scholar] [CrossRef]
- Agung, P.P.; Yuliando, H. Soilless Culture System to Support Water Use Efficiency and Product Quality: A Review. Agric. Agric. Sci. Procedia 2015, 3, 283–288. [Google Scholar] [CrossRef]
- Phantong, P.; Machikowa, T.; Saensouk, P.; Muangsan, N. Comparing growth and physiological responses of Globba schomburgkii Hook. f. and Globba marantina L. under hydroponic and soil conditions. Emir. J. Food Agric. 2018, 30. [Google Scholar] [CrossRef]
- Pienaar, D.; Combrink, N.J.J. The effects of N-source, shading and root zone cooling on two Disa hybrids. S. Afr. J. Plant Soil 2007, 24, 166–171. [Google Scholar] [CrossRef]
- Moorby, J.; Graves, C.J. Root and air temperature effects on growth and yield of tomatoes and lettuce. Acta Hortic. 1980, 29–44. [Google Scholar] [CrossRef]
- Estiarte, M.; Penuelas, J. Alteration of the phenology of leaf senescence and fall in winter deciduous species by climate change: Effects on nutrient proficiency. Glob. Chang. Biol. 2015, 21, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Kooyers, N.J. The evolution of drought escape and avoidance in natural herbaceous populations. Plant Sci. 2015, 234, 155–162. [Google Scholar] [CrossRef]
- Addicott, F.T. Environmental Factors in the Physiology of Abscission. Plant Physiol. 1968, 43, 1471–1479. [Google Scholar]
- Noel, A.R.A.; van Staden, J. Phyllomorph Senescence in Streptocarpus molweniensis. Ann. Bot. 1975, 39, 921–929. [Google Scholar] [CrossRef]
- Marston, M. The morphology of a Streptocarpus hybrid and its regeneration from leaf cuttings. Sci. Hortic. 1964, 17, 114–120. [Google Scholar]
- Rohde, A.; Bhalerao, R.P. Plant dormancy in the perennial context. Trends Plant Sci. 2007, 12, 217–223. [Google Scholar] [CrossRef]
- Gawadi, A.G.; Avery, G.S. Leaf abscission and the so-called abscission layer. Am. J. Bot. 1950, 37, 172–180. [Google Scholar] [CrossRef]
- Matos, F.S.; Borges, L.P.; Müller, C. Ecophysiology Of Leaf Senescence. Agron. Agric. Sci. 2020, 3, 1–6. [Google Scholar] [CrossRef]
- Dhanasekaran, D.; Jasmine, M. Rooting Behavior of Certain Foliage Ornamentals Grown under Hydroponic Nutrient Solutions. J. Floric. 2019, 21. [Google Scholar]
- Sethi, V.P.; Sharma, S.K. Greenhouse heating and cooling using aquifer water. Energy 2007, 32, 1414–1421. [Google Scholar] [CrossRef]
- Grillas, S.; Lucas, M.; Bardopoulou, E.; Voulgari, M. Perlite based soilless culture systems: Current commercial application and prospects. Acta Hortic. 2001, 548, 105–113. [Google Scholar] [CrossRef]
- Calatayud, Á.; Gorbe, E.; Roca, D.; Martínez, P.F. Effect of two nutrient solution temperatures on nitrate uptake, nitrate reductase activity, NH4+ concentration and chlorophyll a fluorescence in rose plants. Environ. Exp. Bot. 2008, 64, 65–74. [Google Scholar] [CrossRef]
- Van Os, E.A. Recent advances in soilless culture in Europe. Acta Hortic. 2017, 1176, 1–8. [Google Scholar] [CrossRef]
- Asao, T.; Asaduzzaman, M.; Mondal, F.M. Horticultural research in Japan. Production of vegetables and ornamentals in hydroponics, constraints and control measures. Adv. Hortic. Sci. 2014, 28, 167–178. [Google Scholar] [CrossRef]
- Morard, P.; Lacoste, L.; Silvestre, J. Effect of oxygen deficiency on mineral nutrition of excised tomato roots. J. Plant Nutr. 2004, 27, 613–626. [Google Scholar] [CrossRef]
- Boudaghpour, S.; Nasir, K. A study on light expanded clay aggregate (LECA) in a geotechnical view and its application on greenhouse and greenroof cultivation. Int. J. Geol. 2008, 2, 59–63. [Google Scholar]
- Jones, J.B. Complete Guide for Growing Plants Hydroponically; CRS Press, Taylor and Francis Group, LLC: Abingdon, UK, 2014; ISBN 978-1-4398-7668. [Google Scholar]
- Nkcukankcuka, M.; Jimoh, M.O.; Griesel, G.; Laubscher, C.P. Growth characteristics, chlorophyll content and nutrients uptake in Tetragonia decumbens Mill. cultivated under different fertigation regimes in hydroponics. Crop Pasture Sci. 2021, 72, 1–12. [Google Scholar] [CrossRef]
- Sharma, N.; Acharya, S.; Kumar, K.; Singh, N.; Chaurasia, O.P. Hydroponics as an advanced technique for vegetable production: An overview. J. Soil Water Conserv. 2018, 17, 364. [Google Scholar] [CrossRef]
- Adams, P. Crop nutrition in hydroponics. Acta Hortic. 1993, 289–306. [Google Scholar] [CrossRef]
- Kratky, B.A. A suspended pot, non-circulating hydroponic method. Acta Hortic. 2004, 648, 83–89. [Google Scholar] [CrossRef]
- Faber, R.J.; Laubscher, C.P.; Rautenbach, F.; Jimoh, M.O. Variabilities in alkaloid concentration of Sceletium tortuosum (L.) N.E. Br in response to different soilless growing media and fertigation regimes in hydroponics. Heliyon 2020, 6, e05479. [Google Scholar] [CrossRef] [PubMed]
- Martens, D. Back to Basics: Streptocarpus. Gesnariads 2013, 63, 47–50. [Google Scholar]
- Uhl, R. Crop Culture Report: Streptocarpus Ladyslippers Series. Greenh. Prod. News 2012. [Google Scholar]
- Pennisi, B.V. The University of Georgia Cooperative Extension; The University of Georgia: Athens, GA, USA, 2009; p. 28. [Google Scholar]
- Kim, J.H.; Lee, A.K.; Roh, M.S.; Suh, J.K. The effect of irradiance and temperature on the growth and flowering of Sinningia cardinalis. Sci. Hortic. (Amst.) 2015, 194, 147–153. [Google Scholar] [CrossRef]
- World Weather Online Port Saint John’s. Eastern Cape, South Africa Weather Averages|Monthly Average High and Low Temperature|Average Precipitation and Rainfall Days. Available online: https://www.worldweatheronline.com/port-saint-johns-weather-averages/eastern-cape/za.aspx (accessed on 24 March 2021).
- Wang, X.; Zhang, W.; Miao, Y.; Gao, L. Root-Zone Warming Differently Benefits Mature and Newly Unfolded Leaves of Cucumis sativus L. Seedlings under Sub-Optimal Temperature Stress. PLoS ONE 2016, 11, e0155298. [Google Scholar] [CrossRef]
- Morgan, J.; Moustafa, A.; Tan, A. Factors affecting the growing-on stages of lettuce and chrysanthemum in nutrient solution culture. Acta Hortic. 1980, 253–262. [Google Scholar] [CrossRef]
- Jimoh, M.O.; Afolayan, A.J.; Lewu, F.B. Heavy metal uptake and growth characteristics of Amaranthus caudatus L. under five different soils in a controlled environment. Not. Bot. Horti Agrobot. 2020, 48, 417–425. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H.; Dickey, D.A. Principles and Procedures of Statistics: A Biometrical Approach, 3rd ed.; McGraw-Hill College: Pennsylvania, PA, USA, 1996; ISBN 978-0070610286. [Google Scholar]
- Wahome, P.K.; Oseni, T.; Masarirambi, M. Effects of different hydroponics systems and growing media on the vegetative growth, yield and cut flower quality of gypsophila (Gypsophila paniculata L.). J. Agric. Sci. 2011, 7, 692–698. [Google Scholar]
- Vogelezang, J.V.M. Effect of root-zone heating on growth, flowering and keeping quality of Saintpaulia. Sci. Hortic. (Amst.) 1988, 34, 101–113. [Google Scholar] [CrossRef]
- Greer, D.H.; Wünsche, J.N.; Norling, C.L.; Wiggins, H.N. Root-zone temperatures affect the phenology of bud break, flower cluster development, shoot extension growth and gas exchange of “Braeburn” (Malus domestica) apple trees. Tree Physiol. 2006, 26, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Moss, G.I.; Dalgleish, R. Increasing returns from roses with root-zone warming. J. Am. Soc. Hortic. Sci. 1984, 109, 893–898. [Google Scholar]
- Nxawe, S.; Laubscher, C.P.; Ndakidemi, P.A. Effect of regulated irrigation water temperature on hydroponics production of Spinach (Spinacia oleracea L.). Afr. J. Agric. Res. 2009, 4, 1442–1446. [Google Scholar]
- Stoltzfus, R.M.B.; Taber, H.G.; Aiello, A.S. Effect of increasing root-zone temperature on growth and nutrient uptake by “Gold Star” muskmelon plants. J. Plant Nutr. 1998, 21, 321–328. [Google Scholar] [CrossRef]
- Zeroni, M.; Gale, J. The effect of root temperature on the development, growth and yield of “Sonia” roses. Sci. Hortic. (Amst.) 1982, 18, 177–184. [Google Scholar] [CrossRef]
- Gislerod, H.R. Physical conditions of propagation media and their influence on the rooting of cuttings. Plant Soil 1983, 75, 1–14. [Google Scholar] [CrossRef]
- Andersen, C.P.; Sucoff, E.I.; Dixon, R.K. Effects of root zone temperature on root initiation and elongation in red pine seedlings. Can. J. For. Res. 1986, 16, 696–700. [Google Scholar] [CrossRef]
- Friis, K.; Christensen, O.V. Flowering of Centradenia inaequilateralis “Cascade” as influenced by temperature and photoperiod. Sci. Hortic. (Amst.) 1989, 41, 125–130. [Google Scholar] [CrossRef]
- Norby, R.J.; Hartz-Rubin, J.S.; Verbrugge, M.J. Phenological responses in maple to experimental atmospheric warming and CO2 enrichment. Glob. Chang. Biol. 2003, 9, 1792–1801. [Google Scholar] [CrossRef]
- Appelgren, M.; Heide, O.M. Regeneration in Streptocarpus Leaf Discs and its Regulation by Temperature and Growth Substances. Physiol. Plant 1972, 27, 417–423. [Google Scholar] [CrossRef]
- van Staden, J. Changes in endogenous cytokinin levels during abscission and senescence of Streptocarpus leaves. J. Exp. Bot. 1973, 24, 667–671. [Google Scholar] [CrossRef]
- Scott-Shaw, C.R.; Victor, J.E.; von Staden, L. Streptocarpus formosus (Hilliard & B.L.Burtt) T.J.Edwards; SANBI: Pretoria, South Africa, 2008. [Google Scholar]
- Maurya, J.P.; Bhalerao, R.P. Photoperiod- and temperature-mediated control of growth cessation and dormancy in trees: A molecular perspective. Ann. Bot. 2017, 120, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Adebooye, C.O.; Schmitz-Eiberger, M.; Lankes, C.; Noga, G.J. Inhibitory effects of sub-optimal root zone temperature on leaf bioactive components, photosystem II (PS II) and minerals uptake in Trichosanthes cucumerina L. Cucurbitaceae. Acta Physiol. Plant 2010, 32, 67–73. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S/N | Treatment Code | Treatment Description |
|---|---|---|
| 1 | WT1 | water temperature heated to 18 °C |
| 2 | WT2 | water temperature heated to 22 °C |
| 3 | WT3 | water temperature heated to 26 °C |
| 4 | WT4 | water temperature heated to 30 °C |
| 5 | WT5 | water temperature heated to 34 °C |
| Treatments | Temp. (°C) | ΔLeaf Number | ΔLeaf Length (cm) | ΔRoot Growth (cm) | ΔTotal Biomass (g) |
|---|---|---|---|---|---|
| WT1 | 18 | 17.7 ± 1.91 a | 36.22 ± 3.75 a | 21.05 ±2.28 a | 107.90 ± 21.07 a |
| WT2 | 22 | 13.3 ± 1.30 b | 15.85 ± 2.24 b | 0.45 ± 0.88 c | 18.67 ± 1.96 b |
| WT3 | 26 | 6.7 ± 0.91 d | 6.68 ± 1.68 c | −5.4 ± 1.02 c | 7.32 ± 1.71 b |
| WT4 | 30 | 3.3 ± 1.56 d | −2.18 ± 2.78 d | −23.3 ± 0.79 bc | −0.80 ± 2.16 b |
| WT5 | 34 | −2.8 ± 0.99 c | −11.76 ± 2.18 e | −48.2 ± 0.41 b | −3.70 ± 1.74 b |
| One-way ANOVA F-statistic | 34.2670 *** | 49.0178 *** | 69.6300 *** | 23.7484 *** | |
| Treatments | Temp. (°C) | Total Number of Buds and Flowers | Total Number of Pedicels |
|---|---|---|---|
| WT1 | 18 | 2.5 ± 0.81 a | 5.00 ± 1.11 a |
| WT2 | 22 | 1.8 ± 0.68 ab | 0.90 ± 0.23 b |
| WT3 | 26 | 0.9 ± 0.35 cd | 0.40 ± 0.22 b |
| WT4 | 30 | 0.1 ± 0.10 d | 0.20 ± 0.13 b |
| WT5 | 34 | 0.0 ± 0.00 d | 0.00 ± 0.00 b |
| One-way ANOVA F-statistic | 4.71716 ** | 16.33995 *** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations All authors have read and agreed to the published version of the manuscript.. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viljoen, C.C.; Jimoh, M.O.; Laubscher, C.P. Studies of Vegetative Growth, Inflorescence Development and Eco-Dormancy Formation of Abscission Layers in Streptocarpus formosus (Gesneriaceae). Horticulturae 2021, 7, 120. https://doi.org/10.3390/horticulturae7060120
Viljoen CC, Jimoh MO, Laubscher CP. Studies of Vegetative Growth, Inflorescence Development and Eco-Dormancy Formation of Abscission Layers in Streptocarpus formosus (Gesneriaceae). Horticulturae. 2021; 7(6):120. https://doi.org/10.3390/horticulturae7060120
Chicago/Turabian StyleViljoen, Cherise Christina, Muhali Olaide Jimoh, and Charles Petrus Laubscher. 2021. "Studies of Vegetative Growth, Inflorescence Development and Eco-Dormancy Formation of Abscission Layers in Streptocarpus formosus (Gesneriaceae)" Horticulturae 7, no. 6: 120. https://doi.org/10.3390/horticulturae7060120
APA StyleViljoen, C. C., Jimoh, M. O., & Laubscher, C. P. (2021). Studies of Vegetative Growth, Inflorescence Development and Eco-Dormancy Formation of Abscission Layers in Streptocarpus formosus (Gesneriaceae). Horticulturae, 7(6), 120. https://doi.org/10.3390/horticulturae7060120