
A Preliminary Comparison of Antioxidants of Tomato Fruit Grown Under Organic and Conventional Systems



{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Plant Preparation
2.2. Total Antioxidant Activity
2.3. Enzyme Assay
2.4. Statistical Analysis
3. Results
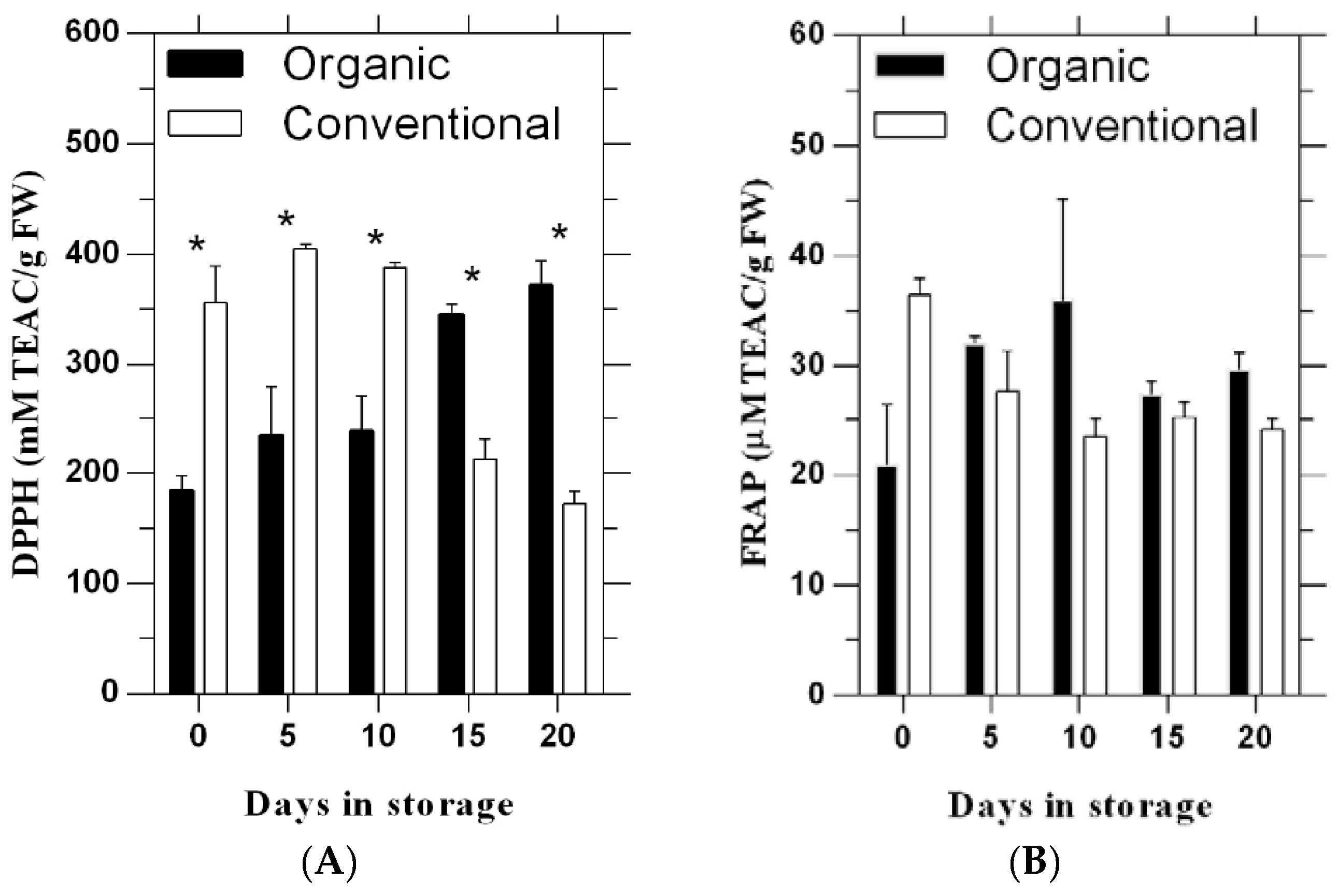
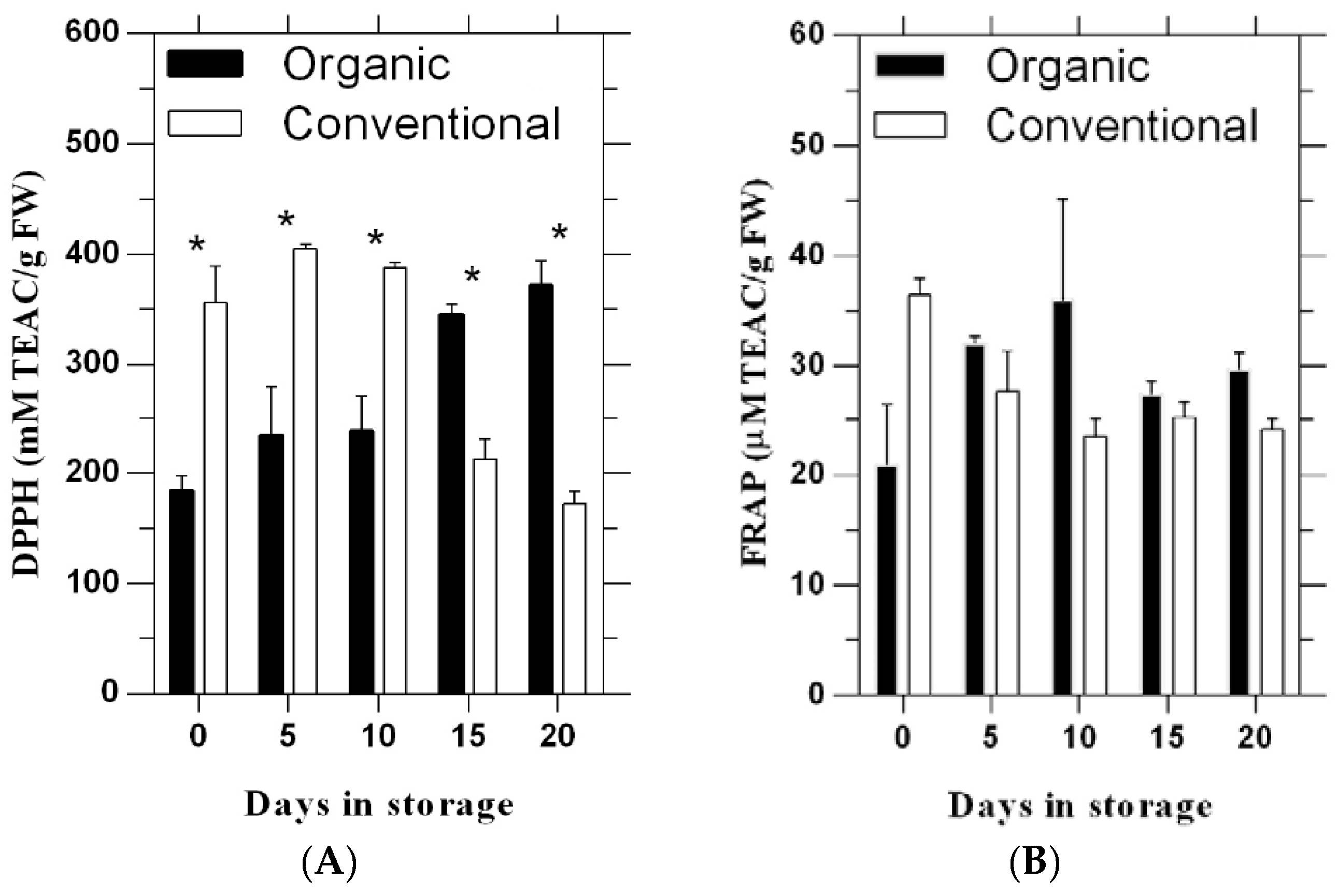
3.1. 2,2′-Diphenyl-l-Picrylhydrazyl (DPPH)
3.2. Ferric Reducing Antioxidant Power (FRAP)
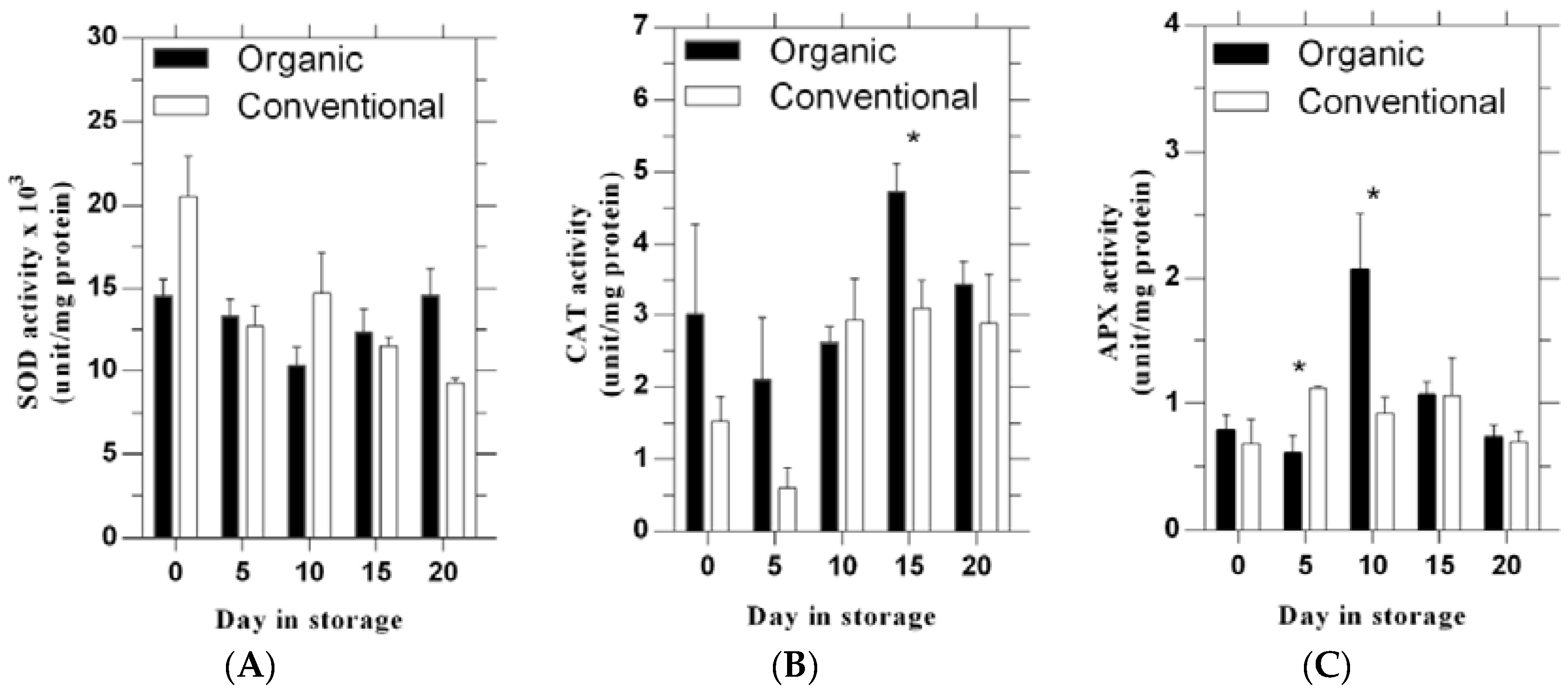
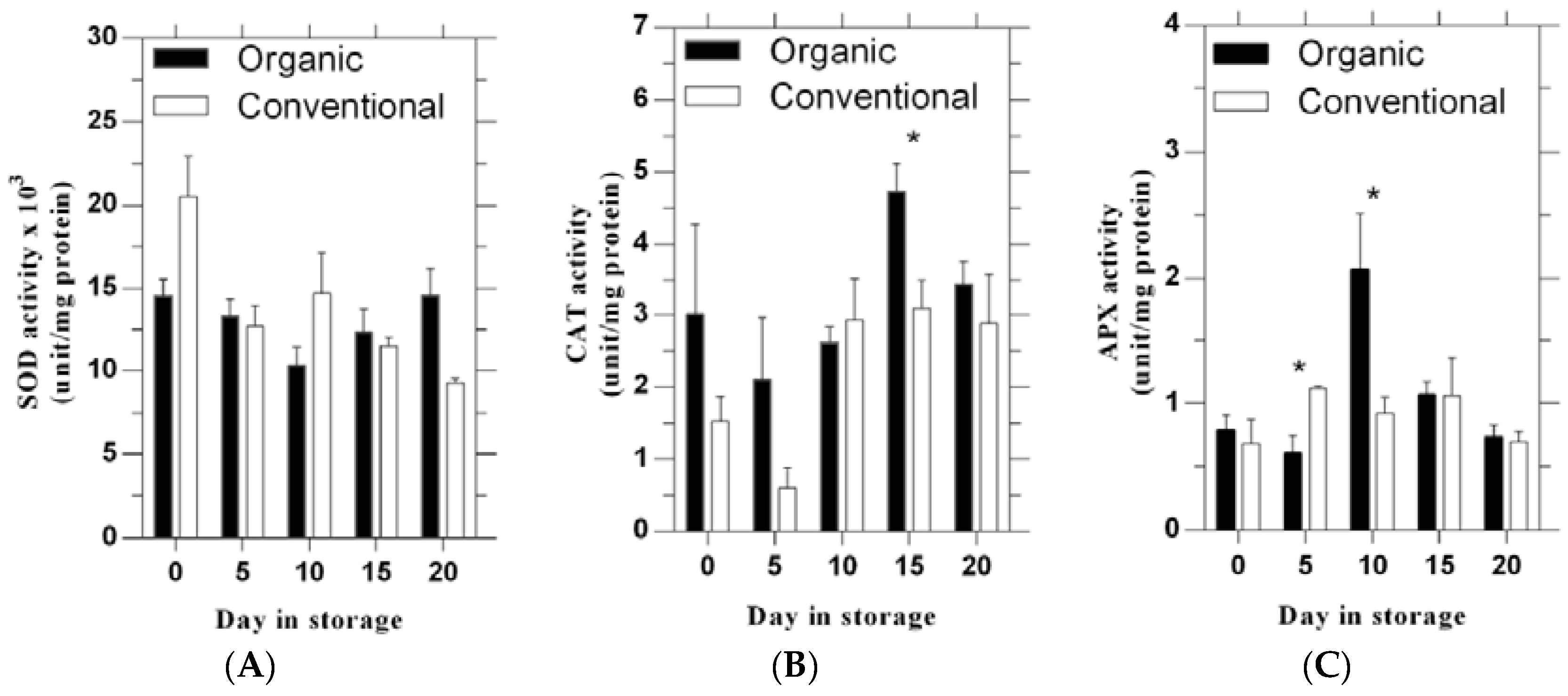
3.3. The Activity of SOD, CAT and APX
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhao, X.; Carey, E.E.; Wang, W.; Rajashekar, C.B. Does organic production enhance phytochemical content of fruits and vegetables? Current knowledge and prospects for research. Horttechnology 2006, 16, 449–456. [Google Scholar]
- Lombardi-Boccia, G.; Lucarini, M.; Lanzi, S.; Aguzzi, A.; Cappelloni, M. Nutrients and antioxidant molecules in yellow plums (Prunus domestica L.) from conventional and organic productions: A comparative study. J. Agric. Food Chem. 2004, 52, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Parka, Y.S.; Imb, M.H.; Choic, J.H.; Yimc, S.H.; Leontowiczd, H.; Leontowiczd, M.; Suhaje, M.; Gorinstein, S. The effects of ethylene treatment on the bioactivity of conventional and organic growing “Hayward” kiwi fruits. Sci. Hortic. 2013, 164, 589–595. [Google Scholar] [CrossRef]
- Parka, Y.S.; Ham, K.S.; Kang, S.G.; Park, Y.K.; Namiesnik, J.; Leontowicz, H.; Leontowicz, M.; Ezra, A.; Trakhtenberg, S.; Gorinstein, S. Organic and conventional kiwi fruits, myths versus reality: Antioxidant, antiproliferative, and health effects. J. Agric. Food Chem. 2012, 60, 6984–6993. [Google Scholar] [CrossRef] [PubMed]
- Worthington, V. Nutritional quality of organic versus conventional fruits, vegetables, and grains. J. Altern. Complement. Med. 2001, 7, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Winter, C.; Davis, S. Organic foods. J. Food Sci. 2006, 71, 117–124. [Google Scholar] [CrossRef]
- Mehdizadeh, M.; Darbandi, E.I.; Naseri-Rad, H.; Tobeh, A. Growth and yield of tomato (Lycopersicon esculentum Mill.) as influenced by different organic fertilizers. Int. J. Agron. Plant Prod. 2013, 4, 734–738. [Google Scholar]
- Reganold, J.P.; Andrews, P.K.; Reeve, J.R.; Carpenter-Boggs, L.; Schadt, C.W.; Alldredge, J.R.; Ross, C.F.; Davies, N.M.; Zhou, J. Fruits and soil quality of organic and conventional strawberry agro ecosystems. PLoS ONE 2010, 5, e12346. [Google Scholar] [CrossRef]
- Toor, R.K.; Geoffrey, P.S.; Anuschka, H. Influence of different types of fertilizers on the major antioxidant components of tomatoes. J. Food Compos. Anal. 2006, 19, 20–27. [Google Scholar] [CrossRef]
- Cano, A.; Acosta, M.; Arnao, M.B. Hydrophilic and lipophilic antioxidant activity changes during on-vine ripening of tomatoes (Lycopersicon esculentum Mill.). Postharvest Biol. Technol. 2003, 28, 59–65. [Google Scholar] [CrossRef]
- Foyer, C.H.; Shigeoka, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Rani, P.; Unni, M.; Karthikeyan, J. Evaluation of antioxidant properties of berries. Indian J. Clin. Biochem. 2004, 19, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Bowler, C.; van Montagu, M.; Inze, D. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Biol. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Abreu, I.A.; Cabelli, D.E. Superoxide dismutases—A review of the metal-associated mechanistic variations. Biochim. Biophys. Acta 2010, 1804, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Willekens, H.; Chamnongpol, S.; Davey, M.; Schraudner, M.; Langebartels, C.; van Montagu, M.; Inzé, D.; van Camp, W. Catalase is a sink for H2O2 and is indispensable for stress defence in C3 plants. EMBO J. 1997, 16, 4806–4816. [Google Scholar] [CrossRef] [PubMed]
- Kangasjärvi, S.; Lepistö, A.; Hännikäinen, K.; Piippo, M.; Luomala, E.M.; Aro, E.M.; Rintamäki, E. Diverse roles for chloroplast stromal and thylakoid-bound ascorbate peroxidases in plant stress responses. Biochem. J. 2008, 412, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.; Mølgaard, J.P. Organic agriculture: Does it enhance or reduce the nutritional value of plant foods? J. Sci. Food Agric. 2001, 81, 924–931. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of free radical method to evaluate antioxidant activity. Lebensm. Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I.; Marschner, H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplast. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Bartoli, C.G.; Simontacchi, M.; Guiamet, J.J.; Montadi, E.; Puntarulo, S. Antioxidant enzymes and lipid peroxidation during aging of Chrysanthemum morifolium RAM petals. Plant Sci. 1995, 104, 161–168. [Google Scholar] [CrossRef]
- Kato, M.; Shimizu, S. Chlorophyll metabolism in higher plants, VII. Chlorophyll degradation in senescence tobacco leaves, phenolic-dependent peroxidative degradation. Can. J. Bot. 1987, 65, 729–735. [Google Scholar] [CrossRef]
- Guil-Guerrero, J.L.; Rebolloso-Fuentes, M.M. Nutrient composition and antioxidant activity of eight tomato (Lycopersicon esculentum) varieties. J. Food Compos. Anal. 2009, 22, 123–129. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Pokorny, J.; Yanishlieva, N.; Gordon, M. Antioxidants in Food: Practical Application; CRC Press: New York, NY, USA, 2001; p. 380. [Google Scholar]
- Jin, P.; Wang, S.Y.; Wang, C.Y.; Zheng, Y. Effect of cultural system and storage temperature on antioxidant capacity and phenolic compounds in strawberries. Food Chem. 2011, 124, 262–270. [Google Scholar] [CrossRef]
- Faller, A.L.K.; Fialho, E. Polyphenol content and antioxidant capacity in organic and conventional plant food. J. Food Compos. Anal. 2010, 23, 561–568. [Google Scholar] [CrossRef]
- Bogs, J.; Bunning, M.; Stushnoff, C. Influence of biologically enhanced organic production on antioxidant and sensory qualities of (Malus x domestica Borkh. cv. Braeburn) apples. Organ. Agric. 2012, 2, 117–126. [Google Scholar] [CrossRef]
- Riahi, A.; Hdider, C. Bioactive compounds and antioxidant activity of organically grown tomato (Solanum lycopersicum L.) cultivars as affected by fertilization. Sci. Hortic. 2013, 151, 90–96. [Google Scholar] [CrossRef]
- Stracke Berenike, A.; Rüfer, C.E.; Bub, A.; Seifert, S.; Weibel, F.P.; Kunz, C.; Watzl, B. No effect of the farming system (organic/conventional) on the bioavailability of apple (Malus domestica Bork. cultivar Golden Delicious) polyphenols in healthy men: A comparative study. Eur. J. Nutr. 2010, 49, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Salandanan, K.; Bunning, M.; Stonaker, F.; Kulen, O.; Kendall, P.; Stushnoff, C. Comparative analysis of antioxidant properties and fruit quality attributes of organically and conventionally grown melons (Cucumis melo L.). Hortscience 2009, 44, 1825–1832. [Google Scholar]
- Oliveira, A.B.; Moura, C.F.H.; Gomes-Filho, E.; Marco, C.A.; Urban, L.; Miranda, M.R.A. The Impact of organic farming on quality of tomatoes is associated to increased oxidative stress during fruit development. PLoS ONE 2013, 8, e56354. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uthairatanakij, A.; Aiamla-or, S.; Jitareerat, P.; Maneenoi, A. A Preliminary Comparison of Antioxidants of Tomato Fruit Grown Under Organic and Conventional Systems. Horticulturae 2017, 3, 21. https://doi.org/10.3390/horticulturae3010021
Uthairatanakij A, Aiamla-or S, Jitareerat P, Maneenoi A. A Preliminary Comparison of Antioxidants of Tomato Fruit Grown Under Organic and Conventional Systems. Horticulturae. 2017; 3(1):21. https://doi.org/10.3390/horticulturae3010021
Chicago/Turabian StyleUthairatanakij, Apiradee, Sukanya Aiamla-or, Pongphen Jitareerat, and Ashariya Maneenoi. 2017. "A Preliminary Comparison of Antioxidants of Tomato Fruit Grown Under Organic and Conventional Systems" Horticulturae 3, no. 1: 21. https://doi.org/10.3390/horticulturae3010021
APA StyleUthairatanakij, A., Aiamla-or, S., Jitareerat, P., & Maneenoi, A. (2017). A Preliminary Comparison of Antioxidants of Tomato Fruit Grown Under Organic and Conventional Systems. Horticulturae, 3(1), 21. https://doi.org/10.3390/horticulturae3010021