
Alternative Growing Media Under the Same Fertigation Scheme Affected Mineral Accumulation and Physiological Parameters in Grapevine Cultivars



Abstract
1. Introduction
2. Materials and Methods
2.1. Grapevine’s Cultivars and Growing Media Preparation
2.2. Plant Growth and Physiology Attributes
2.3. Total Phenolics, Total Flavonoids, and Antioxidant Activity
2.4. Hydrogen Peroxide Content and Lipid Peroxidation
2.5. Plant Mineral Content
2.6. Statistical Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- International Organization of Vine and Wine State of the World Vine and Wine Sector 2021. Int. Organ. Vine Wine Intergov. Organ. 2022, 1–19. Available online: https://www.oiv.int/sites/default/files/documents/OIV_State_of_the_world_Vine_and_Wine_sector_in_2022_2.pdf (accessed on 4 March 2025).
- Santillán, D.; Sotés, V.; Iglesias, A.; Garrote, L. Adapting viticulture to climate change in the Mediterranean region: Evaluations accounting for spatial differences in the producers-climate interactions. BIO Web Conf. 2019, 12, 01001. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Xylia, P.; Litskas, V.; Mandoulaki, A.; Antoniou, D.; Boyias, T.; Stavrinides, M.; Tzortzakis, N. Drought stress and soil management practices in grapevines in Cyprus under the threat of climate change. J. Water Clim. Chang. 2018, 9, 703–712. [Google Scholar] [CrossRef]
- Litskas, V.D.; Irakleous, T.; Tzortzakis, N.; Stavrinides, M.C. Determining the carbon footprint of indigenous and introduced grape varieties through Life Cycle Assessment using the island of Cyprus as a case study. J. Clean. Prod. 2017, 156, 418–425. [Google Scholar] [CrossRef]
- Frioni, T.; Sabbatini, P.; Tombesi, S.; Norrie, J.; Poni, S.; Gatti, M.; Palliotti, A. Effects of a biostimulant derived from the brown seaweed Ascophyllum nodosum on ripening dynamics and fruit quality of grapevines. Sci. Hortic. 2018, 232, 97–106. [Google Scholar] [CrossRef]
- Copper, A.W.; Johnson, T.E.; Danner, L.; Bastian, S.E.P.; Collins, C. Preliminary Sensory and chemical profiling of Cypriot wines made from indigenous grape varieties Xynisteri, Maratheftiko and Giannoudhi and acceptability to Australian consumers. OENO One 2019, 53, 229–248. [Google Scholar] [CrossRef]
- Koufos, G.; Mavromatis, T.; Koundouras, S.; Fyllas, N.M.; Jones, G.V. Viticulture-climate relationships in Greece: The impacts of recent climate trends on harvest date variation. Int. J. Climatol. 2014, 34, 1445–1459. [Google Scholar] [CrossRef]
- Fraga, H.; García de Cortázar Atauri, I.; Malheiro, A.C.; Santos, J.A. Modelling climate change impacts on viticultural yield, phenology and stress conditions in Europe. Glob. Chang. Biol. 2016, 22, 3774–3788. [Google Scholar] [CrossRef]
- Tzortzakis, N.; Chrysargyris, A.; Aziz, A. Adaptive response of a native mediterranean grapevine cultivar upon short-term exposure to drought and heat stress in the context of climate change. Agronomy 2020, 10, 249. [Google Scholar] [CrossRef]
- Shah, S.U.; Ayub, Q.; Hussain, I.; Khan, S.M.; Ali, S.; Khan, M.A.; Haq, N.; Mehmood, A.; Khan, T.; Brahmi, N.C. Effect of different growing media on survival and growth of Grape (Vitus vinifera) cuttings. J. Adv. Nutr. Sci. Technol. 2021, 1, 117. [Google Scholar] [CrossRef]
- Alikhani, L.; Ansari, K.; Jamnezhad, M.; Tabatabaie, Z. The effect of different mediums and cuttings on growth and rooting of pomegranate cuttings. Iran. J. Plant Physiol. 2011, 1, 199–203. [Google Scholar]
- Waite, H.; Whitelaw-Weckert, M.; Torley, P. Grapevine propagation: Principles and methods for the production of high-quality grapevine planting material. N. Zeal. J. Crop Hortic. Sci. 2015, 43, 144–161. [Google Scholar] [CrossRef]
- Ausari, P.K.; Soni, N.; Kanpure, R.N.; Ninama, N.; Bhandari, J. Effect of Different Growing Media on Hardwood Cutting of Grapes (Vitis vinifera L.) cv. Pusa Navrang. Int. J. Plant Soil Sci. 2023, 35, 978–988. [Google Scholar] [CrossRef]
- Zareei, E.; Zaare-Nahandi, F.; Hajilou, J.; Oustan, S. Eliciting effects of magnetized solution on physiological and biochemical characteristics and elemental uptake in hydroponically grown grape (Vitis vinifera L. cv. Thompson Seedless). Plant Physiol. Biochem. 2021, 167, 586–595. [Google Scholar] [CrossRef]
- Savvas, D.; Gruda, N. Application of soilless culture technologies in the modern greenhouse industry—A review. Eur. J. Hortic. Sci. 2018, 83, 280–293. [Google Scholar] [CrossRef]
- Ferrón-Carrillo, F.; Cunha-Chiamolera, T.P.L.d.; Urrestarazu, M. Effect of ammonium nitrogen on pepper grown under soilless culture. J. Plant Nutr. 2021, 45, 113–122. [Google Scholar] [CrossRef]
- Specht, K.; Siebert, R.; Hartmann, I.; Freisinger, U.B.; Sawicka, M.; Werner, A.; Thomaier, S.; Henckel, D.; Walk, H.; Dierich, A. Urban agriculture of the future: An overview of sustainability aspects of food production in and on buildings. Agric. Hum. Values 2014, 31, 33–51. [Google Scholar] [CrossRef]
- Huang, L.; Gu, M.; Yu, P.; Zhou, C.; Liu, X. Biochar and vermicompost amendments affect substrate properties and plant growth of basil and tomato. Agronomy 2020, 10, 224. [Google Scholar] [CrossRef]
- Wystalska, K.; Malińska, K.; Sobik-Szołtysek, J.; Dróżdż, D.; Meers, E. Properties of Poultry-Manure-Derived Biochar for Peat Substitution in Growing Media. Materials 2023, 16, 6392. [Google Scholar] [CrossRef] [PubMed]
- Hirschler, O.; Thrän, D. Peat Substitution in Horticulture: Interviews with German Growing Media Producers on the Transformation of the Resource Base. Horticulturae 2023, 9, 919. [Google Scholar] [CrossRef]
- Herrera, F.; Castillo, J.E.; Chica, A.F.; López Bellido, L. Use of municipal solid waste compost (MSWC) as a growing medium in the nursery production of tomato plants. Bioresour. Technol. 2008, 99, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Niu, G.; Starman, T.; Volder, A.; Gu, M. Poinsettia Growth and Development Response to Container Root Substrate with Biochar. Horticulturae 2018, 4, 1. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Prasad, M.; Tzortzakis, N. Wood-Based Biochar Ratio Used for Partial Peat Replacement in Growing Media for Antirrhinum majus Pot Production. Agriculture 2024, 14, 1860. [Google Scholar] [CrossRef]
- Mehmood, T.; Ahmad, W.; Shafique Ahmad, K.; Shafi, J.; Asif Shehzad, M.; Aqeel Sarwar, M. Comparative Effect of Different Potting Media on Vegetative and Reproductive Growth of Floral Shower (Antirrhinum majus L.). Univers. J. Plant Sci. 2013, 1, 104–111. [Google Scholar] [CrossRef]
- Prasad, M.; Chrysargyris, A.; McDaniel, N.; Kavanagh, A.; Gruda, N.S.; Tzortzakis, N. Plant nutrient availability and pH of biochars and their fractions, with the possible use as a component in a growing media. Agronomy 2020, 10, 10. [Google Scholar] [CrossRef]
- Gruda, N. Current and future perspective of growing media in Europe. Acta Hortic. 2012, 960, 37–43. [Google Scholar] [CrossRef]
- Messiga, A.J.; Hao, X.; Ziadi, N.; Dorais, M. Reducing peat in growing media: Impact on nitrogen content, microbial activity, and CO2 and N2O emissions. Can. J. Soil Sci. 2021, 102, 77–87. [Google Scholar] [CrossRef]
- Ekbic, H.B.; Yaman, E.; Özenç, D.B.; Ekbic, E. Effects of Hazelnut Husk Compost and Tea Residue Compost on Quality and Performance of 5 Bb American Grapevine Rootstock Saplings. Acta Sci. Pol. Hortorum Cultus 2022, 21, 15–23. [Google Scholar] [CrossRef]
- Saqib, M.; Razzaq, K.; Ullah, S.; Hussain, A.; Rajwana, I.A.; Naz, A.; Akhtar, G.; Amin, M.; Faried, H.N.; Zafar, M.S.; et al. Standardization of Growing Media for Grapes Nursery Production. J. Plant Environ. 2021, 3, 115–124. [Google Scholar] [CrossRef]
- Ronga, D.; Francia, E.; Allesina, G.; Pedrazzi, S.; Zaccardelli, M.; Pane, C.; Tava, A.; Bignami, C. Valorization of Vineyard By-Products to Obtain Composted Digestate and Biochar Suitable for Nursery Grapevine (Vitis vinifera L.) Production. Agronomy 2019, 9, 420. [Google Scholar] [CrossRef]
- Amir, M.; Hussain, S.; Martin, S.; Asad Latif, M.; Basit, A.; Ahmad, N.; Khan, F.; Ullah, W.; Ullah, R.; Abbas, M. Different media effect on various cultivars of grapes (Vitis vinifera) cuttings. J. Xi’an Shiyou Univ. 2023, 19, 468–484. [Google Scholar]
- Vrontis, D.; Paliwoda, S.J. Branding and the Cyprus wine industry. J. Brand Manag. 2008, 16, 145–159. [Google Scholar] [CrossRef]
- Tzortzakis, N.G.; Economakis, C.D. Shredded maize stems as an alternative substrate medium: Effect on growth, flowering and yield of tomato in soilless culture. J. Veg. Sci. 2005, 11, 57–70. [Google Scholar] [CrossRef]
- Marinou, E.; Chrysargyris, A.; Tzortzakis, N. Use of sawdust, coco soil and pumice in hydroponically grown strawberry. Plant Soil Environ. 2013, 59, 452–459. [Google Scholar] [CrossRef]
- Tzortzakis, N.; Pitsikoulaki, G.; Stamatakis, A.; Chrysargyris, A. Ammonium to Total Nitrogen Ratio Interactive Effects with Salinity Application on Solanum lycopersicum Growth, Physiology, and Fruit Storage in a Closed Hydroponic System. Agronomy 2022, 12, 386. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Charalambous, S.; Xylia, P.; Litskas, V.; Stavrinides, M.; Tzortzakis, N. Assessing the biostimulant effects of a novel plant-based formulation on tomato crop. Sustainability 2020, 12, 8432. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of Chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Meyers, K.J.; Watkins, C.B.; Pritts, M.P.; Liu, R.H. Antioxidant and Antiproliferative Activities of Strawberries. J. Agric. Food Chem. 2003, 51, 6887–6892. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Czemerys, R. Antioxidant activity and phenolic compounds in 32 selected herbs. Food Chem. 2007, 105, 940–949. [Google Scholar] [CrossRef]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef]
- De Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; Abreu, C.E.B.D.; Gomes-Filho, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Farooq, M.; Kakar, K.; Golly, M.K.; Ilyas, N.; Zib, B.; Khan, I.; Khan, S.; Khan, I.; Saboor, A.; Bakhtiar, M. Comparative Effect of Potting Media on Sprouting and Seedling Growth of Grape Cuttings. Int. J. Environ. Agric. Res. 2018, 4, 82–89. [Google Scholar]
- Popescu, G.C.; Popescu, M. Effects of different potting growing media for Petunia grandiflora and Nicotiana alata Link & Otto on photosynthetic capacity, leaf area, and flowering potential. Chil. J. Agric. Res. 2015, 75, 21–26. [Google Scholar]
- Santos, C.V. Regulation of chlorophyll biosynthesis and degradation by salt stress in sunflower leaves. Sci. Hortic. 2004, 103, 93–99. [Google Scholar] [CrossRef]
- Bresson, J.; Vasseur, F.; Dauzat, M.; Koch, G.; Granier, C.; Vile, D. Quantifying spatial heterogeneity of chlorophyll fluorescence during plant growth and in response to water stress. Plant Methods 2015, 11, 23. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Xylia, P.; Antoniou, O.; Tzortzakis, N. Climate change due to heat and drought stress can alter the physiology of Maratheftiko local cyprian grapevine variety. J. Water Clim. Chang. 2018, 9, 715–727. [Google Scholar] [CrossRef]
- Heyman, L.; Chrysargyris, A.; Demeestere, K.; Tzortzakis, N.; Höfte, M. Responses to drought stress modulate the susceptibility to Plasmopara viticola in Vitis vinifera self-rooted cuttings. Plants 2021, 10, 273. [Google Scholar] [CrossRef]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent developments in enzymatic antioxidant defence mechanism in plants with special reference to abiotic stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef]
- Pintać, D.; Četojević-Simin, D.; Berežni, S.; Orčić, D.; Mimica-Dukić, N.; Lesjak, M. Investigation of the chemical composition and biological activity of edible grapevine (Vitis vinifera L.) leaf varieties. Food Chem. 2019, 286, 686–695. [Google Scholar] [CrossRef]
- Dorion, S.; Ouellet, J.C.; Rivoal, J. Glutathione metabolism in plants under stress: Beyond reactive oxygen species detoxification. Metabolites 2021, 11, 641. [Google Scholar] [CrossRef] [PubMed]
- Tzortzakis, N.; Chrysargyris, A. Olive-mill and grape-mill residue impact the growth, physiology and nutrient status of grapevines young cuttings. Sustain. Chem. Pharm. 2024, 37, 101362. [Google Scholar] [CrossRef]
- Singh, A.; Satheeshkumar, P.K. Reactive Oxygen Species (ROS) and ROS Scavengers in Plant Abiotic Stress Response. In Stress Biology in Photosynthetic Organisms; Mishra, A.K., Ed.; Springer: Singapore, 2024; pp. 44–64. ISBN 9789819718825. [Google Scholar]
- Agarwal, P.; Saha, S.; Hariprasad, P. Agro-industrial-residues as potting media: Physicochemical and biological characters and their influence on plant growth. Biomass Convers. Biorefin. 2021, 13, 9601–9624. [Google Scholar] [CrossRef]
- Dubey, T.; Mishra, D.S. Effect of rooting media and different polythene wrappers on air layering of pomegranate (Punica granatum) cv. Bhagwa. Int. J. Adv. Biochem. Res. 2024, 8, 401–406. [Google Scholar] [CrossRef]
- Cerovic, Z.G.; Masdoumier, G.; Ghozlen, N.B.; Latouche, G. A new optical leaf-clip meter for simultaneous non-destructive assessment of leaf chlorophyll and epidermal flavonoids. Physiol. Plant. 2012, 146, 251–260. [Google Scholar] [CrossRef]
- Singh, K.K.; Chauhan, J.S. A review on vegetative propagation of grape (Vitis vinifera L.) through cutting. Glob. J. Biosci. Biotechnol. 2020, 9, 50–55. [Google Scholar]
- Khan, F.; Siddique, A.B.; Shabala, S.; Zhou, M.; Zhao, C. Phosphorus Plays Key Roles in Regulating Plants’ Physiological Responses to Abiotic Stresses. Plants 2023, 12, 2861. [Google Scholar] [CrossRef]
- Cetner, M.D.; Kalaji, H.M.; Borucki, W.; Kowalczyk, K. Phosphorus deficiency affects the i-step of chlorophyll a fluorescence induction curve of radish. Photosynthetica 2020, 58, 671–681. [Google Scholar] [CrossRef]
- Méndez, A.; Paz-Ferreiro, J.; Gil, E.; Gascó, G. The effect of paper sludge and biochar addition on brown peat and coir based growing media properties. Sci. Hortic. 2015, 193, 225–230. [Google Scholar] [CrossRef]
- Abad, M.; Noguera, P.; Puchades, R.; Maquieira, A.; Noguera, V. Physico-chemical and chemical properties of some coconut coir dusts for use as a peat substitute for containerised ornamental plants. Bioresour. Technol. 2002, 82, 241–245. [Google Scholar] [CrossRef]
- Tangolar, S.; Tangolar, S.; Tarım, G.; Ada, M.; Torun, A.A.; Ertargın, E. The effects of different nitrogen and potassium levels on yield and quality of two early grape cultivars grown in different soilless media. Acta Hortic. 2019, 1242, 349–356. [Google Scholar] [CrossRef]
- Baronti, S.; Montagnoli, A.; Beatrice, P.; Danieli, A.; Maienza, A.; Vaccari, F.P.; Casini, D.; Di Gennaro, S.F. Above- and below-ground morpho-physiological traits indicate that biochar is a potential peat substitute for grapevine cuttings nursery production. Sci. Rep. 2024, 14, 17185. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, F.; Mogensen, L.; Smith, A.M.; Larsen, S.U.; Knudsen, M.T. Greenhouse gas emissions from bio-based growing media: A life-cycle assessment. Sci. Total Environ. 2024, 907, 167977. [Google Scholar] [CrossRef] [PubMed]
- Papafotiou, M.; Kargas, G.; Lytra, I. Olive-mill waste compost as a growth medium component for foliage potted plants. HortScience 2005, 40, 1746–1750. [Google Scholar] [CrossRef]
- Jamal Uddin, A.F.M.; Rakibuzzaman, M.; Raisa, I.; Maliha, M.; Husna, M.A. Impact of natural substances and synthetic hormone on grapevine cutting. J. Biosci. Agric. Res. 2020, 25, 2069–2074. [Google Scholar] [CrossRef]
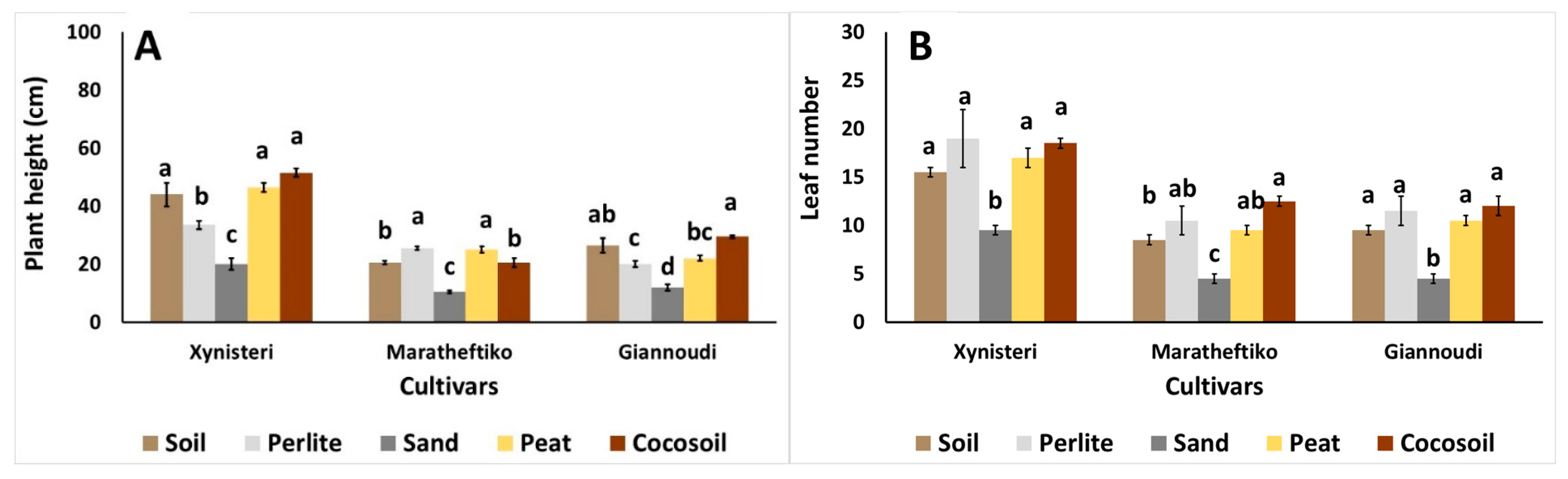

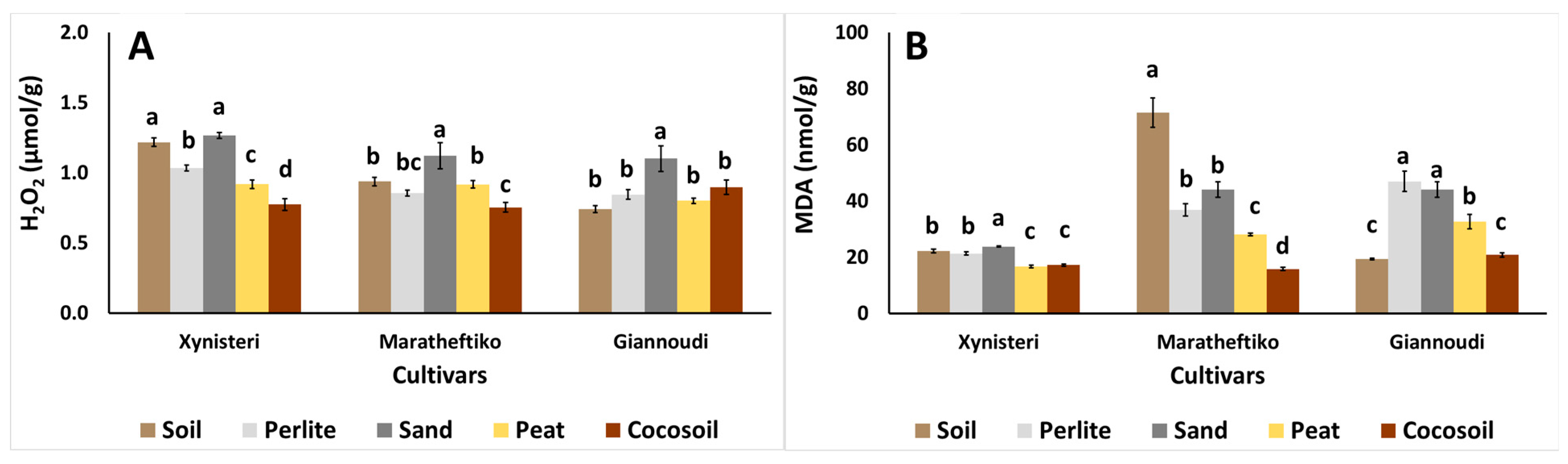
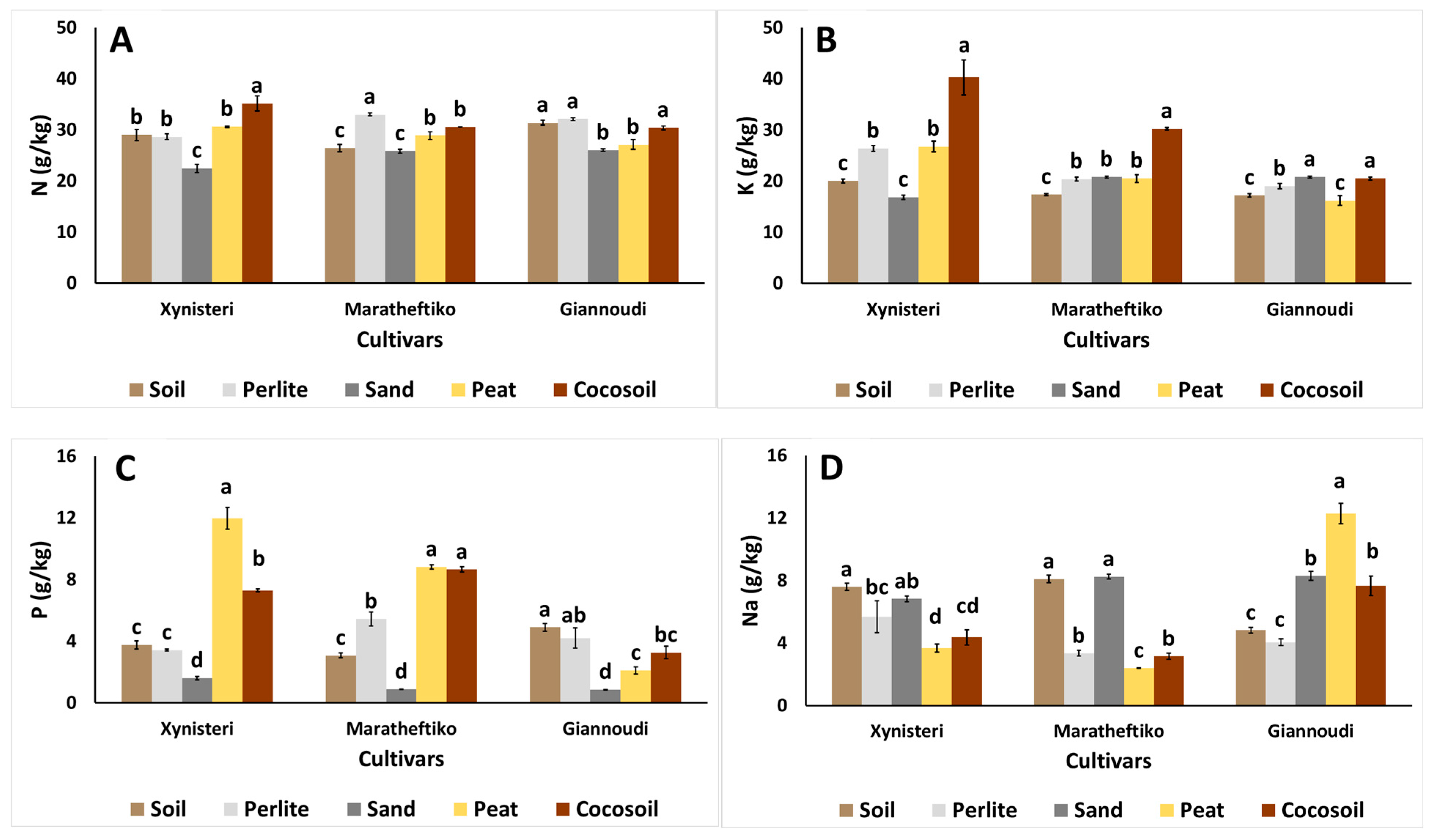

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SPAD | Chl Fluorescence (Fv/Fm) | Chl a | Chl b | Total Chls | Total Car | Chl a:Chl b | Total Car/Total Chls | ||
|---|---|---|---|---|---|---|---|---|---|
| Xynisteri | Soil | 34.80 ± 1.88 ab | 0.771 ± 0.015 | 1.27 ± 0.03 | 0.54 ± 0.02 | 1.81 ± 0.05 | 0.18 ± 0.002 b | 2.33 ± 0.02 | 1.102 ± 0.003 |
| Perlite | 38.12 ± 1.31 a | 0.767 ± 0.028 | 1.18 ± 0.05 | 0.49 ± 0.03 | 1.68 ± 0.09 | 0.16 ± 0.008 b | 2.39 ± 0.06 | 0.099 ± 0.005 | |
| Sand | 34.35 ± 1.46 ab | 0.770 ± 0.010 | 1.17 ± 0.01 | 0.52 ± 0.01 | 1.69 ± 0.02 | 0.17 ± 0.008 b | 2.26 ± 0.03 | 0.098 ± 0.004 | |
| Peat | 32.19 ± 0.54 b | 0.775 ± 0.009 | 1.27 ± 0.08 | 0.57 ± 0.05 | 1.84 ± 0.13 | 0.21 ± 0.006 a | 2.25 ± 0.09 | 0.117 ± 0.009 | |
| Cocosoil | 35.57 ± 0.99 ab | 0.815 ± 0.024 | 1.26 ± 0.08 | 0.54 ± 0.05 | 1.80 ± 0.14 | 0.18 ± 0.011 b | 2.31 ± 0.07 | 0.104 ± 0.005 | |
| Maratheftiko | Soil | 34.69 ± 1.72 a | 0.756 ± 0.004 b | 1.14 ± 0.13 b | 0.53 ± 0.08 b | 1.68 ± 0.22 b | 0.15 ± 0.002 c | 2.16 ± 0.10 a | 0.093 ± 0.011 a |
| Perlite | 32.92 ± 1.44 ab | 0.786 ± 0.005 a | 1.57 ± 0.01 a | 0.91 ± 0.01 a | 2.48 ± 0.02 a | 0.15 ± 0.001 c | 1.72 ± 0.00 b | 0.061 ± 0.000 b | |
| Sand | 18.61 ± 1.81 c | 0.605 ± 0.010 c | 1.16 ± 0.02 b | 0.52 ± 0.01 b | 1.68 ± 0.02 b | 0.17 ± 0.002 b | 2.22 ± 0.01 a | 0.103 ± 0.001 a | |
| Peat | 32.20 ± 1.27 ab | 0.778 ± 0.003 a | 1.19 ± 0.05 b | 0.53 ± 0.03 b | 1.73 ± 0.09 b | 0.19 ± 0.005 a | 2.24 ± 0.06 a | 0.107 ± 0.003 a | |
| Cocosoil | 29.77 ± 1.15 b | 0.777 ± 0.007 a | 1.10 ± 0.04 b | 0.47 ± 0.03 b | 1.57 ± 0.08 b | 0.15 ± 0.004 c | 2.35 ± 0.06 a | 0.100 ± 0.007 a | |
| Giannoudi | Soil | 32.79 ± 0.87 a | 0.800 ± 0.035 | 1.32 ± 0.06 a | 0.60 ± 0.06 a | 1.92 ± 0.12 a | 0.21 ± 0.002 a | 2.23 ± 0.11 ab | 0.111 ± 0.005 bc |
| Perlite | 39.69 ± 1.44 a | 0.772 ± 0.008 | 0.95 ± 0.00 c | 0.38 ± 0.01 b | 1.33 ± 0.00 c | 0.16 ± 0.006 c | 2.45 ± 0.04 a | 0.121 ± 0.002 ab | |
| Sand | 18.76 ± 6.21 b | 0.509 ± 0.154 | 1.17 ± 0.01 ab | 0.52 ± 0.00 ab | 1.69 ± 0.01 ab | 0.17 ± 0.001 c | 2.22 ± 0.01 b | 0.099 ± 0.000 c | |
| Peat | 33.49 ± 0.62 a | 0.775 ± 0.008 | 1.09 ± 0.02 bc | 0.45 ± 0.01 ab | 1.55 ± 0.03 bc | 0.19 ± 0.001 ab | 2.41 ± 0.03 ab | 0.127 ± 0.003 a | |
| Cocosoil | 31.58 ± 1.59 a | 0.761 ± 0.014 | 1.07 ± 0.12 bc | 0.48 ± 0.05 ab | 1.54 ± 0.17 bc | 0.18 ± 0.013 bc | 2.23 ± 0.05 ab | 0.117 ± 0.005 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tzortzakis, N.; Chrysargyris, A. Alternative Growing Media Under the Same Fertigation Scheme Affected Mineral Accumulation and Physiological Parameters in Grapevine Cultivars. Horticulturae 2025, 11, 479. https://doi.org/10.3390/horticulturae11050479
Tzortzakis N, Chrysargyris A. Alternative Growing Media Under the Same Fertigation Scheme Affected Mineral Accumulation and Physiological Parameters in Grapevine Cultivars. Horticulturae. 2025; 11(5):479. https://doi.org/10.3390/horticulturae11050479
Chicago/Turabian StyleTzortzakis, Nikolaos, and Antonios Chrysargyris. 2025. "Alternative Growing Media Under the Same Fertigation Scheme Affected Mineral Accumulation and Physiological Parameters in Grapevine Cultivars" Horticulturae 11, no. 5: 479. https://doi.org/10.3390/horticulturae11050479
APA StyleTzortzakis, N., & Chrysargyris, A. (2025). Alternative Growing Media Under the Same Fertigation Scheme Affected Mineral Accumulation and Physiological Parameters in Grapevine Cultivars. Horticulturae, 11(5), 479. https://doi.org/10.3390/horticulturae11050479