

Biofortification of Arugula Microgreens Through Supplemental Blue Light
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
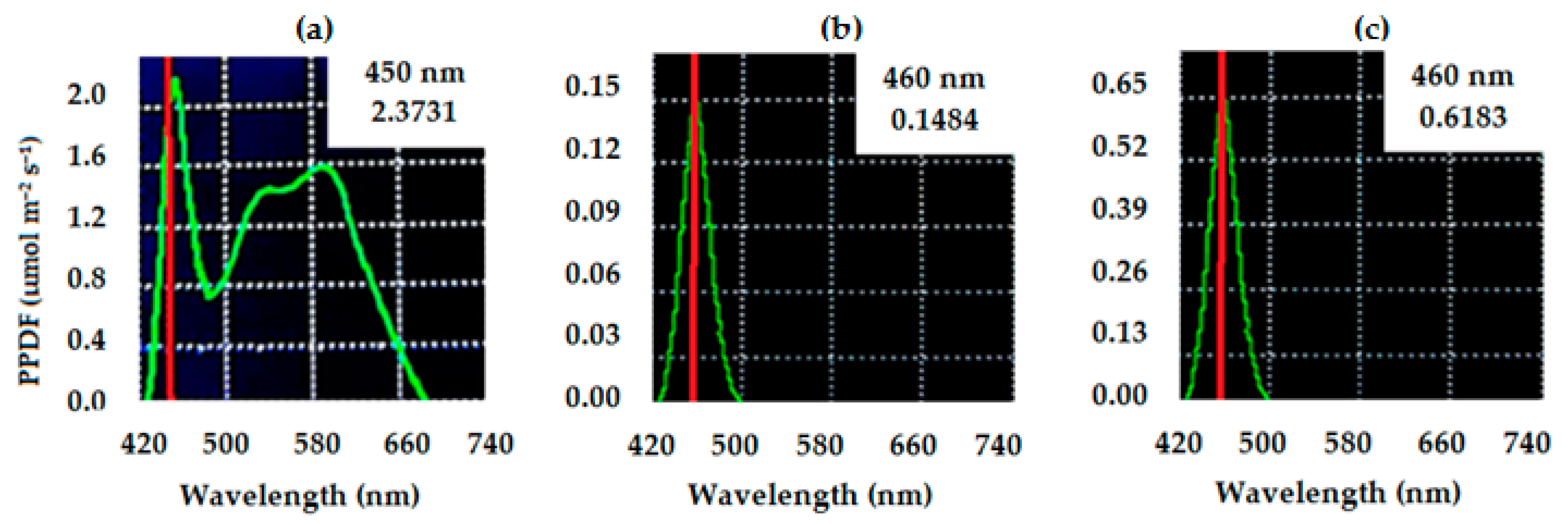
2.2. Treatments and Experimental Design
2.3. Experimental Conditions
2.4. Evaluated Characteristics
- (a)
- Hypocotyl length (cm): A total of 50 seedlings were randomly and carefully cut at the level of the substrate, which, prior to sowing, had its surface flattened and the seeds distributed over it. After cutting, the seedlings were placed on the surface of the laboratory table and measured with a graduated ruler.
- (b)
- Cotyledon area (cm2 plant−1): The cotyledons from the 50 seedlings per experimental unit, used to measure the hypocotyl length, were separated from the seedlings and passed through an LI 3100 electronic bench area meter (LI-COR, Tucson, AZ, USA) to quantify the cotyledon area.
- (c)
- Yield (g m−2): The microgreens from one tray were harvested by cutting the hypocotyl close to the substrate and immediately weighed on a scale with 0.01 g precision.
- (d)
- Chlorophyll and carotenoid contents (μg g−1 fresh mass, FM): A hole punch was used to collect the discs of cotyledons with diameters of 0.5 cm, totaling 0.025 to 0.030 g of fresh mass. The leaf discs were placed in Eppendorf Tubes® with 1.5 mL of 80% acetone solution. The Eppendorf Tubes® were protected from light and placed in a refrigerator, where the samples remained for 48 h at 5–7 °C. Then, the samples were left at room temperature, and readings were performed at 663 nm for chlorophyll a, 647 nm for chlorophyll b and 470 nm for carotenoids with a Du Series 600 spectrophotometer (Beckman®, Brea, CA, USA). Subsequently, the pigment contents were calculated using the formulas proposed by Lichtenthaler [27]. The analyses were performed in triplicate.
- (e)
- Vitamin C (mg 100 g−1): The ascorbic acid content was determined by weighing 2 g of fresh microgreens and adding 10 mL of 0.5% cold oxalic acid solution. Then, the mixture was filtered. Amounts of 1 mL of the filtered extract and 4 mL of 0.5% oxalic acid were transferred to an Erlenmeyer® flask, mixed and titrated with 2,6-dichloroindophenol sodium salt up to the turning point, as described by Strohecker and Henning [28]. The analyses were performed in triplicate.
- (f)
- Phenolic compounds (mg gallic acid equivalent (GAE) 100 g−1 FM): This was determined by the spectrophotometry of an extract obtained according to Larrauri et al. [29] with the use of 70% acetone and 50% methanol solutions and subsequent addition of the Folin–Ciocalteu reagent, being based on an alkaline oxidation–reduction reaction where the phenolate ion was oxidized and the Folin reagent was reduced. A total of 300 mg of cotyledon FM was used. After the reaction, a blue color was produced, and the reading was performed at an absorbance of 700 nm, being expressed as the GAE. The analyses were performed in triplicate.
- (g)
- Antioxidant power (%): A total of 1 g of the sample was weighed in a dark environment and placed in a beaker, where 5 mL of 50% methanol was added and the mixture was homogenized and left to rest for 60 min. Subsequently, it was centrifuged at 15,000× g for 15 min, and supernatant 1 was placed in a 10 mL volumetric flask. In the second step, 4 mL of 70% acetone was added to the residue of the first extraction and homogenized, left at rest for 60 min; then, this residue was centrifuged at 15,000× g for 15 min, and supernatant 2 was collected in the same 10 mL volumetric flask. Subsequently, the flask was completed with distilled water, resulting in the extract. In the third step, 0.1 mL of the extract and 3.9 mL of the 0.06 mM DPPH radical were added to a test tube and homogenized in a tube shaker. Methyl alcohol was used as a blank to calibrate the spectrophotometer. Readings were taken at 515 nm, as described by Larrauri et al. [29]. The analyses were performed in triplicate.
- (h)
- Macronutrient (g kg−1), iron and zinc (mg kg−1) contents: The aerial parts of the microgreens were washed in deionized water and dried in an oven at 65 °C. After drying using the method described by Miyasawa et al. [30], the samples were prepared for reading of the N content using the Kjeldahl method; P and S by spectrophotometry; and K, Ca, Mg, Fe and Zn by atomic absorption.
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ampim, P.A.; Obeng, E.; Olvera-Gonzalez, E. Indoor vegetable production: An alternative approach to increasing cultivation. Plants 2022, 11, 2843. [Google Scholar] [CrossRef] [PubMed]
- United Nations (UN). End Hunger, Achieve Food Security and Improved Nutrition and Promote Sustainable Agriculture. Available online: https://sustainabledevelopment.un.org/sdg2 (accessed on 22 November 2022).
- Shibaeva, T.G.; Sherudilo, E.G.; Rubaeva, A.A.; Titov, A.F. Continuous LED lighting enhances yield and nutritional value of four genotypes of Brassicaceae microgreens. Plants 2022, 11, 176. [Google Scholar] [CrossRef]
- Horizon Grand View Research. China Microgreens Market Size & Outlook, 2023–2030. 2025. Available online: https://www.grandviewresearch.com/horizon/outlook/microgreens-market/china (accessed on 6 February 2025).
- Mendes, F.Q.; Purquerio, L.F.V.; Carvalho, R.F.; Cecílio Filho, A.B. White light intensities for maximum yield and quality of arugula microgreens. Pesqui. Agropecu. Trop. 2024, 54, e79951. [Google Scholar] [CrossRef]
- Xiao, Z.; Rausch, S.; Luo, Y.; Sun, J.; Yu, L.; Wang, Q.; Chein, P.; Stommel, J. Microgreens of Brassicaceae: Genetic diversity of phytochemical concentrations and antioxidant capacity. LWT 2019, 101, 731–737. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, Z.; Ager, E.; Kong, L.; Tan, L. Nutritional quality and health benefits of microgreens, a crop of modern agriculture. J. Future Foods 2021, 1, 58–66. [Google Scholar] [CrossRef]
- Lanoue, J.; Louis, S.; Little, C.; Hao, X. Continuous lighting can improve yield and reduce energy costs while increasing or maintaining nutritional contents of microgreens. Front. Plant Sci. 2022, 13, 983222. [Google Scholar] [CrossRef]
- Kusuma, P.; Pattison, P.M.; Bugbee, B. From physics to fixtures to food: Current and potential LED efficacy. Hortic. Res. 2020, 7, 56. [Google Scholar] [CrossRef]
- Yang, X.; Xu, H.; Shao, L.; Li, T.; Wang, Y.; Wang, R. Response of photosynthetic capacity of tomato leaves to different LED light wavelength. Environ. Exp. Bot. 2018, 150, 161–171. [Google Scholar] [CrossRef]
- Nguyen, T.K.L.; Cho, K.M.; Lee, H.Y.; Cho, D.Y.; Lee, G.O.; Jang, S.N.; Lee, Y.; Kim, D.; Son, K.-H. Effects of white LED lighting with specific shorter blue and/or green wavelength on the growth and quality of two lettuce cultivars in a vertical farming system. Agronomy 2021, 11, 2111. [Google Scholar] [CrossRef]
- Medelo, M.J.Y.; Alves, T.N.; Ribera, L.M.; Silva, L.N.C.; Carvalho, R.F.; Calori, A.H.; Cecilio Filho, A.B. Harvest time, photoperiod and white light irradiance on yield of red cabbage microgreens in plant factory. Chil. J. Agric. Res. 2025, 85, 181–192. [Google Scholar] [CrossRef]
- Pennisi, G.; Pistillo, A.; Orsini, F.; Cellini, A.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Crepaldi, A.; Gianquinto, G.; Marcelis, L.F. Optimal light intensity for sustainable water and energy use in indoor cultivation of lettuce and basil under red and blue LEDs. Sci. Hortic. 2020, 272, 109508. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Sams, C.E.; Morrow, R.C. Blue wavelengths from LED lighting increase nutritionally important metabolites in specialty crops. HortScience 2015, 50, 1285–1288. [Google Scholar] [CrossRef]
- Kang, W.H.; Park, J.S.; Park, K.S.; Son, J.E. Leaf photosynthetic rate, growth, and morphology of lettuce under different fractions of red, blue, and green light from light-emitting diodes (LEDs). Hortic. Environ. Biote 2016, 57, 573–579. [Google Scholar] [CrossRef]
- Kang, L.K.; Huang, Y.J.; Lim, W.T.; Hsu, P.H.; Hwang, P.A. Growth, pigment content, antioxidant activity, and phytoene desaturase gene expression in Caulerpa lentillifera grown under different combinations of blue and red light-emitting diodes. J. Appl. Phycol. 2020, 32, 1971–1982. [Google Scholar] [CrossRef]
- Zheng, L.; He, H.; Song, W. Application of light-emitting diodes and the effect of light quality on horticultural crops: A review. HortScience 2019, 54, 1656–1661. [Google Scholar] [CrossRef]
- Zhang, X.; Bian, Z.; Yuan, X.; Chen, X.; Lu, C. A review on the effects of light-emitting diode (LED) light on the nutrients of sprouts and microgreens. Trends Food Sci. Technol. 2020, 99, 203–216. [Google Scholar] [CrossRef]
- Brazaitytė, A.; Miliauskienė, J.; Vaštakaitė-Kairienė, V.; Sutulienė, R.; Laužikė, K.; Duchovskis, P.; Małek, S. Effect of different ratios of blue and red led light on brassicaceae microgreens under a controlled environment. Plants 2021, 10, 801. [Google Scholar] [CrossRef]
- Gödecke, T.; Stein, A.J.; Qaim, M. The global burden of chronic and hidden hunger: Trend and determinants. Glob. Food Sec. 2018, 18, 21–29. [Google Scholar] [CrossRef]
- Buturi, C.V.; Mauro, R.P.; Fogliano, V.; Leonardi, C.; Giuffrida, F. Mineral biofortification of vegetables as a tool to improve human diet. Foods 2021, 10, 223. [Google Scholar] [CrossRef]
- Kong, Y.; Masabni, J.; Niu, G. Effect of temperature variation and blue and red leds on the elongation of arugula and mustard microgreens. Horticulturae 2023, 9, 608. [Google Scholar] [CrossRef]
- Son, K.-H.; Oh, M.-M. Leaf shape, growth, and antioxidant phenolic compounds of two lettuce cultivars grown under various combinations of blue and red light-emitting diodes. HortScience 2013, 48, 988–995. [Google Scholar] [CrossRef]
- Johnson, R.E.; Kong, Y.; Zheng, Y. Elongation growth mediated by blue light varies with light intensities and plant species: A comparison with red light in arugula and mustard seedlings. Environ. Exp. Bot. 2020, 169, 103898. [Google Scholar] [CrossRef]
- Ouzounis, T.; Fretté, X.; Ottosen, C.O.; Rosenqvist, E. Spectral effects of LEDs on chlorophyll fluorescence and pigmentation in Phalaenopsis ‘Vivien’and ‘Purple Star’. Physiol. Plant 2015, 154, 314–327. [Google Scholar] [CrossRef]
- Furlani, P.R.; Bolonhezi, D.; Silveira, L.C.P.; Faquin, V. Nutrição mineral de hortaliças, preparo e manejo de soluções nutritivas. Inf. Agropecu. 1999, 20, 90–98. [Google Scholar]
- Lichtenthaler, H.K. [34] Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1987; Volume 148, pp. 350–382. [Google Scholar] [CrossRef]
- Strohecker, R.; Henining, H.M. Análisis de Vitaminas: Métodos Comprobados; Paz Montalvo: Madrid, Spain, 1967; 428p. [Google Scholar]
- Larrauri, J.A.; Rupérez, P.; Saura-Calixto, F. Effect of drying temperature on the stability of polyphenols and antioxidant activity of red grape pomace peels. J. Agric. Food Chem. 1997, 45, 1390–1393. [Google Scholar] [CrossRef]
- Miyazawa, M.; Pavan, M.A.; Muraoka, T.; Carmo, C.A.F.S.; Mello, W.J. Análises químicas de tecido vegetal. In Manual de Análises Químicas de Solos, Plantas e Fertilizantes; Silva, F.C., Ed.; Embrapa Informação Tecnológica: Brasília, Brazil, 2009; pp. 193–233. [Google Scholar]
- Barbosa, J.C.; Maldonado Júnior, W. Experimentação Agronômica e AgroEstat: Sistema para Análises Estatísticas de Ensaios Agronômicos; Multipress: Jaboticabal, Brazil, 2015. [Google Scholar]
- Snowden, M.C.; Cope, K.R.; Bugbee, B. Sensitivity of seven diverse species to blue and green light: Interactions with photon flux. PLoS ONE 2016, 11, e0163121. [Google Scholar] [CrossRef]
- Noguchi, A.; Amaki, W. Effects of light quality on the growth and essential oil production in Mexican mint. In Proceedings of the VIII International Symposium on Light in Horticulture, East Lansing, MI, USA, 22–26 May 2016. [Google Scholar]
- Dou, H.; Niu, G.; Gu, M.; Masabni, J.G. Effects of light quality on growth and phytonutrient accumulation of herbs under controlled environments. Horticulturae 2017, 3, 36. [Google Scholar] [CrossRef]
- Landi, M.; Zivcak, M.; Sytar, O.; Brestic, M.; Allakhverdiev, S.I. Plasticity of photosynthetic processes and the accumulation of secondary metabolites in plants in response to monochromatic light environments: A review. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148131. [Google Scholar] [CrossRef]
- Akbari, B.; Baghaei-Yazdi, N.; Bahmaie, M.; Mahdavi Abhari, F. The role of plant-derived natural antioxidants in reduction of oxidative stress. BioFactors 2022, 48, 611–633. [Google Scholar] [CrossRef]
- Zivcak, M.; Brestic, M.; Kalaji, H.M.; Govindjee. Photosynthetic responses of sun-and shade-grown barley leaves to high light: Is the lower PSII connectivity in shade leaves associated with protection against excess of light? Photosynth. Res. 2014, 119, 339–354. [Google Scholar] [CrossRef]
- Brunetti, C.; Guidi, L.; Sebastiani, F.; Tattini, M. Isoprenoids and phenylpropanoids are key components of the antioxidant defense system of plants facing severe excess light stress. Environ. Exp. Bot. 2015, 119, 54–62. [Google Scholar] [CrossRef]
- Petroutsos, D.; Tokutsu, R.; Maruyama, S.; Flori, S.; Greiner, A.; Magneschi, L.; Cusant, L.; Kottke, T.; Mittag, M.; Hegemann, P.; et al. A blue-light photoreceptor mediates the feedback regulation of photosynthesis. Nature 2016, 537, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Zivcak, M.; Brestic, M.; Botyanszka, L.; Chen, Y.E.; Allakhverdiev, S.I. Phenotyping of isogenic chlorophyll-less bread and durum wheat mutant lines in relation to photoprotection and photosynthetic capacity. Photosynth. Res. 2019, 139, 239–251. [Google Scholar] [CrossRef]
- Hamdani, S.; Khan, N.; Perveen, S.; Qu, M.; Jiang, J.; Govindjee; Zhu, X.G. Changes in the photosynthesis properties and photoprotection capacity in rice (Oryza sativa) grown under red, blue, or white light. Photosynth. Res. 2019, 139, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zuo, Z.; Wang, X.; Gu, L.; Yoshizumi, T.; Yang, Z.; Yang, L.; Liu, Q.; Han, Y.-J.; Kim, J.-I.; et al. Photoactivation and inactivation of Arabidopsis cryptochrome 2. Science 2016, 354, 343–347. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Q.; Wang, X.; Zuo, Z.; Oka, Y.; Lin, C. New insights into the mechanisms of phytochrome–cryptochrome coaction. New Phytol. 2018, 217, 547–551. [Google Scholar] [CrossRef]
- Samuolienė, G.; Brazaitytė, A.; Jankauskienė, J.; Viršilė, A.; Sirtautas, R.; Novičkovas, A.; Sakalauskienė, S.; Sakalauskienė, J.; Duchovskis, P. LED irradiance level affects growth and nutritional quality of Brassica microgreens. Cent. Eur. J. Biol. 2013, 8, 1241–1249. [Google Scholar] [CrossRef]

{kind=link}
| Blue Light Irradiation Time (h) | ||||
|---|---|---|---|---|
| PPFD | 1 | 2 | 1 | 2 |
| (µmol m−2 s−1) | HL (cm) | CA (cm2 plant−1) | ||
| 5 | 8.37 Aa | 7.85 Ba * | 2.43 Aa | 1.39 Ba * |
| 20 | 8.30 Aa | 7.15 Bb * | 2.45 Aa | 1.16 Bb * |
| Control | 8.38 | 2.48 | ||
| Blue Light Irradiation Time (h) | ||||
|---|---|---|---|---|
| PPFD | 1 | 2 | 1 | 2 |
| (µmol m−2 s−1) | Yield (g m−2) | Phenols (mg GAE 100 g−1) | ||
| 5 | 3907.40 Aa | 3044.75 Ba * | 178.41 Ab | 179.08 Ab |
| 20 | 3926.57 Aa | 2877.21 Bb * | 202.15 Ba * | 249.17 Aa * |
| Control | 3933.79 | 166.88 | ||
| Blue Light Irradiation Time (h) | ||||
|---|---|---|---|---|
| PPFD | 1 | 2 | 1 | 2 |
| (µmol m−2 s−1) | Chlor a (µg g−1) | Chlor b (µg g−1) | ||
| 5 | 1.70 Ab * | 2.05 Ab * | 1.20 Ab * | 1.31 Ab * |
| 20 | 2.81 Aa * | 2.68 Aa * | 1.76 Aa * | 2.12 Aa * |
| Control | 0.84 | 0.65 | ||
| Car (µg g−1) | Vit C (mg 100g−1) | |||
| 5 | 0.70 Ab * | 0.84 Ab * | 80.64 Bb * | 95.47 Ab * |
| 20 | 1.22 Aa * | 1.24 Aa * | 103.68 Ba * | 153.73 Aa * |
| Control | 0.33 | 36.84 | ||
| AOP (%) | ||||
| 5 | 73.18 Bb * | 75.57 Ab * | ||
| 20 | 84.09 Ba * | 85.65 Aa * | ||
| Control | 67.00 | |||
| Blue Light Irradiation Time (h) | ||||
|---|---|---|---|---|
| PPFD | 1 | 2 | 1 | 2 |
| (µmol m−2 s−1) | N (g kg−1) | P (g kg−1) | ||
| 5 | 47.4 Aa * | 46.2 Aa * | 21.3 Aa * | 19.6 Bb * |
| 20 | 46.8 Aa * | 47.2 Aa * | 21.0 Aa * | 20.6 Aa * |
| Control | 44.5 | 17.9 | ||
| K (g kg−1) | Ca (g kg−1) | |||
| 5 | 36.8 Aa | 32.0 Aa | 14.8 Aa * | 15.4 Aa * |
| 20 | 33.8 Aa | 32.1 Aa | 15.6 Aa * | 14.4 Aa * |
| Control | 32.1 | 12.7 | ||
| Mg (g kg−1) | S (g kg−1) | |||
| 5 | 6.9 Aa | 7.0 Aa | 17.1 Aa | 17.5 Bb |
| 20 | 6.6 Aa | 7.3 Aa | 16.4 Aa | 16.4 Aa |
| Control | 6.8 | 16.8 | ||
| Fe (mg kg−1) | Zn (mg kg−1) | |||
| 5 | 433 Aa | 433 Aa | 134 Aa * | 134 Aa * |
| 20 | 495 Aa | 578 Aa | 130 Aa * | 152 Aa |
| Control | 497 | 161 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendes, F.Q.; Carvalho, R.F.; Souza, M.O.d.; Cecílio Filho, A.B. Biofortification of Arugula Microgreens Through Supplemental Blue Light. Horticulturae 2025, 11, 412. https://doi.org/10.3390/horticulturae11040412
Mendes FQ, Carvalho RF, Souza MOd, Cecílio Filho AB. Biofortification of Arugula Microgreens Through Supplemental Blue Light. Horticulturae. 2025; 11(4):412. https://doi.org/10.3390/horticulturae11040412
Chicago/Turabian StyleMendes, Franciele Quintino, Rogério Falleiros Carvalho, Manuela Oliveira de Souza, and Arthur Bernardes Cecílio Filho. 2025. "Biofortification of Arugula Microgreens Through Supplemental Blue Light" Horticulturae 11, no. 4: 412. https://doi.org/10.3390/horticulturae11040412
APA StyleMendes, F. Q., Carvalho, R. F., Souza, M. O. d., & Cecílio Filho, A. B. (2025). Biofortification of Arugula Microgreens Through Supplemental Blue Light. Horticulturae, 11(4), 412. https://doi.org/10.3390/horticulturae11040412