Optimizing Planting Density for Increased Resource Use Efficiency in Baby-Leaf Production of Lettuce (Lactuca sativa L.) and Basil (Ocimum basilicum L.) in Vertical Farms

 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site and Growing Condition
2.2. Plant Material
2.3. Light Management
2.4. Nutrient Solution Management
2.5. Growth Analysis and Resource Use Efficiency
2.6. Stomatal Conductance
2.7. Relative Chlorophyll Content
2.8. Experimental Design and Treatments
2.9. Statistical Analysis
3. Results
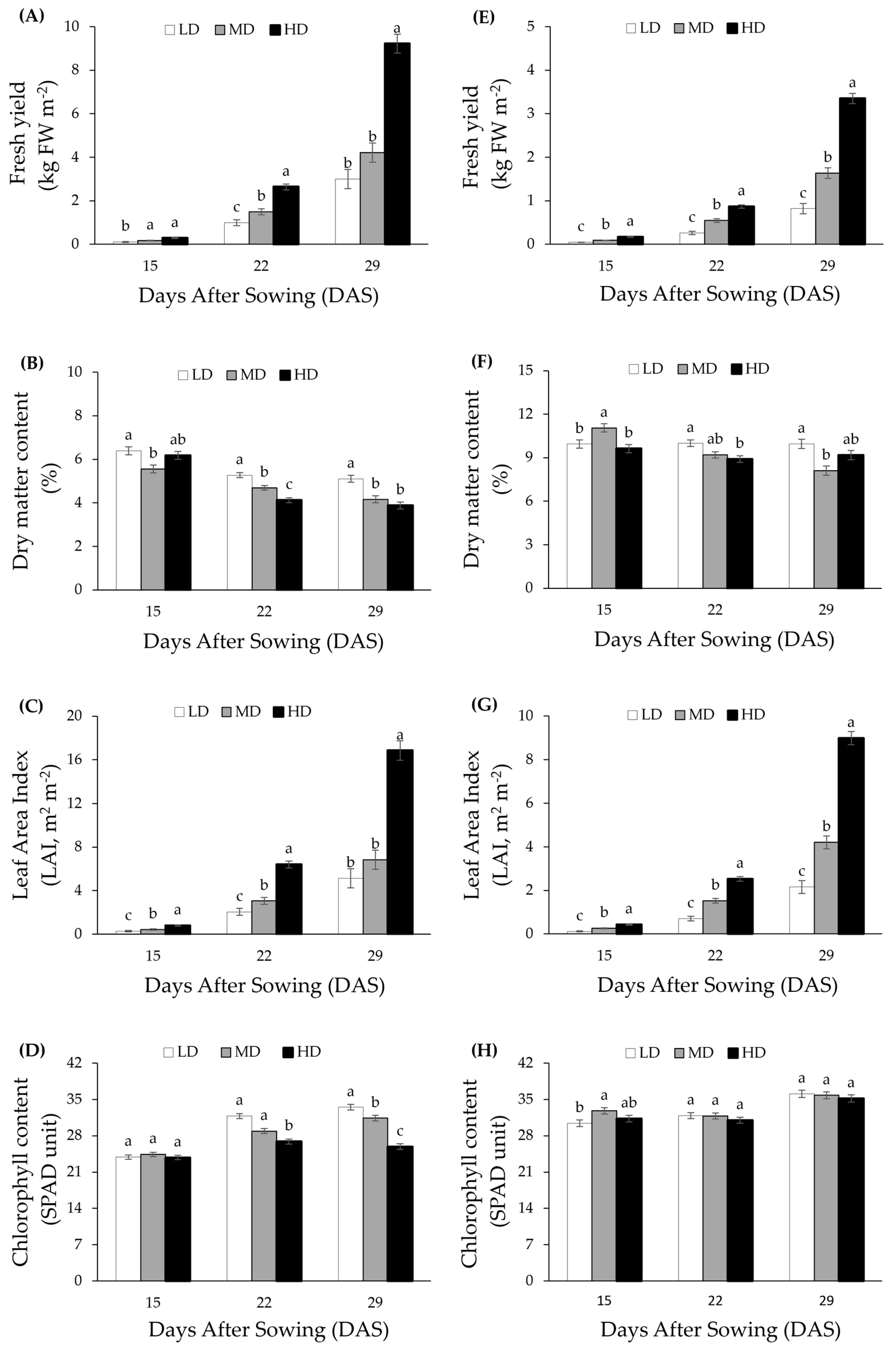
3.1. Effect of Planting Density on Biomass Production
3.2. Effect of Planting Density on Plant Morphological Response
3.3. Effect of Planting Density on Leaf Chlorophyll Content and Stomatal Conductance
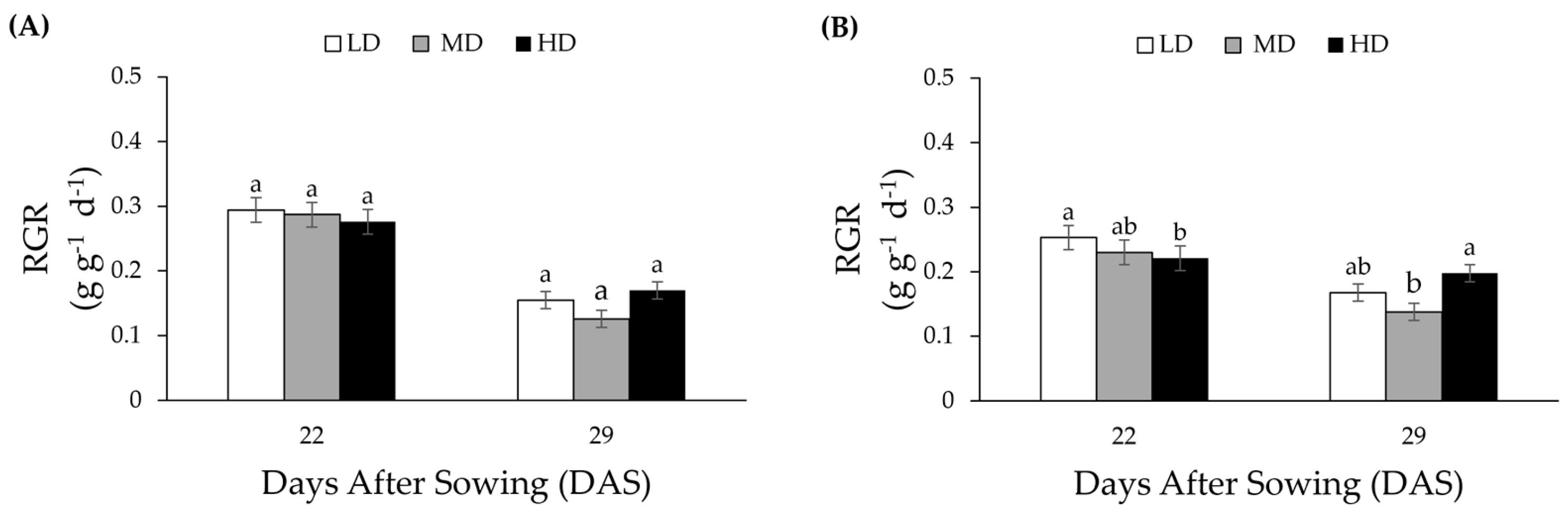
3.4. Effect of Planting Density on Relative Growth Rate
3.5. Effect of Planting Density on Light and Energy Use Efficiencies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kozai, T. Resource use efficiency of closed plant production system with artificial light: Concept, estimation and application to plant factory. Proc. Jpn. Acad. Ser. B 2013, 89, 447–461. [Google Scholar] [CrossRef]
- Martin, M. Life cycle management in vertical farming: Insights from vertical farming sustainability assessments. Acta Hortic. 2023, 1369, 235–242. [Google Scholar] [CrossRef]
- Miladinov, G. Impacts of population growth and economic development on food security in low-income and middle-income countries. Front. Hum. Dyn. 2023, 5, 1121662. [Google Scholar] [CrossRef]
- Carotti, L.; Pistillo, A.; Zauli, I.; Meneghello, D.; Martin, M.; Pennisi, G.; Gianquinto, G.; Orsini, F. Improving water use efficiency in vertical farming: Effects of growing systems, far-red radiation and planting density on lettuce cultivation. Agric. Water Manag. 2023, 285, 108365. [Google Scholar] [CrossRef]
- Van Delden, S.H.; SharathKumar, M.; Butturini, M.; Graamans, L.J.A.; Heuvelink, E.; Kacira, M.; Kaiser, E.; Klamer, R.S.; Klerkx, L.; Kootstra, G.; et al. Current status and future challenges in implementing and upscaling vertical farming systems. Nat. Food 2021, 2, 944–956. [Google Scholar] [CrossRef]
- Zauli, I.; Pennisi, G. Are Vertical Farms Sustainable? Front. Young Minds 2023, 11, 964569. [Google Scholar] [CrossRef]
- Sheibani, F.; Bourget, M.; Morrow, R.C.; Mitchell, C.A. Close-canopy lighting, an effective energy-saving strategy for overhead sole-source LED lighting in indoor farming. Front. Plant Sci. 2023, 14, 1215919. [Google Scholar] [CrossRef]
- Orsini, F.; Pennisi, G.; Zulfiqar, F.; Gianquinto, G. Sustainable use of resources in plant factories with artificial lighting (PFALs). Eur. J. Hortic. Sci. 2020, 85, 297–309. [Google Scholar] [CrossRef]
- Pennisi, G.; Orsini, F.; Blasioli, S.; Cellini, A.; Crepaldi, A.; Braschi, I.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Stanghellini, C.; et al. Resource use efficiency of indoor lettuce (Lactuca sativa L.) cultivation as affected by red:blue ratio provided by LED lighting. Sci. Rep. 2019, 9, 14127. [Google Scholar] [CrossRef]
- Pennisi, G.; Pistillo, A.; Orsini, F.; Cellini, A.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Crepaldi, A.; Gianquinto, G.; Marcelis, L.F.M. Optimal light intensity for sustainable water and energy use in indoor cultivation of lettuce and basil under red and blue LEDs. Sci. Hortic. 2020, 272, 109508. [Google Scholar] [CrossRef]
- Pennisi, G.; Orsini, F.; Landolfo, M.; Pistillo, A.; Crepaldi, A.; Nicola, S.; Fernández, J.A.; Marcelis, L.F.M.; Gianquinto, G. Optimal photoperiod for indoor cultivation of leafy vegetables and herbs. Eur. J. Hortic. Sci. 2020, 85, 329–338. [Google Scholar] [CrossRef]
- Jin, W.; Urbina, J.L.; Heuvelink, E.; Marcelis, L.F.M. Adding Far-Red to Red-Blue Light-Emitting Diode Light Promotes Yield of Lettuce at Different Planting Densities. Front. Plant Sci. 2021, 11, 609977. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, Z.; Li, J.; Wang, R. Optimum Planting Density Improves Resource Use Efficiency and Yield Stability of Rainfed Maize in Semiarid Climate. Front. Plant Sci. 2021, 12, 752606. [Google Scholar] [CrossRef] [PubMed]
- Fekadu, G.M.; Getachew, T.; Zebenay, D.; Gizachew, A.; Fasil, T.T. Effect of planting density on yield and yield components of lettuce (Lactuca sativa L.) at two agro-ecologies of Ethiopia. Afr. J. Agric. Res. 2021, 17, 549–556. [Google Scholar] [CrossRef]
- Van Gerrewey, T.; Boon, N.; Geelen, D. Vertical Farming: The Only Way Is Up? Agronomy 2021, 12, 2. [Google Scholar] [CrossRef]
- Larsen, D.H.; Woltering, E.J.; Nicole, C.C.S.; Marcelis, L.F.M. Response of Basil Growth and Morphology to Light Intensity and Spectrum in a Vertical Farm. Front. Plant Sci. 2020, 11, 597906. [Google Scholar] [CrossRef]
- Kozai, T. Current Status of Plant Factories with Artificial Lighting (PFALs) and Smart PFALs. In Smart Plant Factory; Kozai, T., Ed.; Springer: Singapore, 2018; pp. 3–13. ISBN 9789811310645. [Google Scholar] [CrossRef]
- Shi, M.; Gu, J.; Wu, H.; Rauf, A.; Emran, T.B.; Khan, Z.; Mitra, S.; Aljohani, A.S.M.; Alhumaydhi, F.A.; Al-Awthan, Y.S.; et al. Phytochemicals, Nutrition, Metabolism, Bioavailability, and Health Benefits in Lettuce—A Comprehensive Review. Antioxidants 2022, 11, 1158. [Google Scholar] [CrossRef]
- Singletary, K.W. Basil: A Brief Summary of Potential Health Benefits. Nutr. Today 2018, 53, 92–97. [Google Scholar] [CrossRef]
- Touliatos, D.; Dodd, I.C.; McAinsh, M. Vertical farming increases lettuce yield per unit area compared to conventional horizontal hydroponics. Food Energy Secur. 2016, 5, 184–191. [Google Scholar] [CrossRef]
- Zauli, I.; Rossini, E.; Pennisi, G.; Martin, M.; Crepaldi, A.; Gianquinto, G.; Orsini, F. The Perfect Match: Testing the Effect of Increasing Red and Blue Ratio on Baby-Leaf Kale Growth, Yield and Physiology. Horticulturae 2024, 10, 1134. [Google Scholar] [CrossRef]
- Farhangi, H.; Mozafari, V.; Roosta, H.R.; Shirani, H.; Farhangi, M. Optimizing growth conditions in vertical farming: Enhancing lettuce and basil cultivation through the application of the Taguchi method. Sci. Rep. 2023, 13, 6717. [Google Scholar] [CrossRef] [PubMed]
- Nicole, C.C.S.; Charalambous, F.; Martinakos, S.; Van De Voort, S.; Li, Z.; Verhoog, M.; Krijn, M. Lettuce growth and quality optimization in a plant factory. Acta Hortic. 2016, 231–238. [Google Scholar] [CrossRef]
- Postma, J.A.; Hecht, V.L.; Hikosaka, K.; Nord, E.A.; Pons, T.L.; Poorter, H. Dividing the pie: A quantitative review on plant density responses. Plant Cell Environ. 2021, 44, 1072–1094. [Google Scholar] [CrossRef] [PubMed]
- Gifford, R.M.; Thorne, J.H.; Hitz, W.D.; Giaquinta, R.T. Crop Productivity and Photoassimilate Partitioning. Science 1984, 225, 801–808. [Google Scholar] [CrossRef]
- Li, Y.-C.; Dai, H.-Y.; Chen, H. Effects of plant density on the aboveground dry matter and radiation-use efficiency of field corn. PLoS ONE 2022, 17, e0277547. [Google Scholar] [CrossRef]
- Wu, L.; Deng, Z.; Cao, L.; Meng, L. Effect of plant density on yield and quality of perilla sprouts. Sci. Rep. 2020, 10, 9937. [Google Scholar] [CrossRef]
- Kawser, A.Q.M.R.; Hossain, M.A.; Yeasmin, M.F. Optimizing the planting density of lettuce (Lactuca sativa) with tilapia (Oreochromis niloticus) in a recirculation aquaponic system. Am. J. Agric. Sci. Eng. Technol. 2017, 4, 1–12. [Google Scholar] [CrossRef]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C.-O. Spectral Effects of Artificial Light on Plant Physiology and Secondary Metabolism: A Review. HortScience 2015, 50, 1128–1135. [Google Scholar] [CrossRef]
- Kim, H.-H. Stomatal Conductance of Lettuce Grown Under or Exposed to Different Light Qualities. Ann. Bot. 2004, 94, 691–697. [Google Scholar] [CrossRef]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant responses to rising vapor pressure deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef]
- Damour, G.; Simonneau, T.; Cochard, H.; Urban, L. An overview of models of stomatal conductance at the leaf level: Models of stomatal conductance. Plant Cell Environ. 2010, 33, 1419–1438. [Google Scholar] [CrossRef] [PubMed]
- Collado, C.E.; Hwang, S.J.; Hernández, R. Supplemental greenhouse lighting increased the water use efficiency, crop growth, and cutting production in Cannabis sativa. Front. Plant Sci. 2024, 15, 1371702. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Zhang, Y.; Zhang, Y.; Bian, Z.; Fanourakis, D.; Yang, Q.; Li, T. Morphological and physiological properties of indoor cultivated lettuce in response to additional far-red light. Sci. Hortic. 2019, 257, 108725. [Google Scholar] [CrossRef]
- Teobaldelli, M.; Basile, B.; Giuffrida, F.; Romano, D.; Toscano, S.; Leonardi, C.; Rivera, C.M.; Colla, G.; Rouphael, Y. Analysis of Cultivar-Specific Variability in Size-Related Leaf Traits and Modeling of Single Leaf Area in Three Medicinal and Aromatic Plants: Ocimum basilicum L., Mentha spp., and Salvia spp. Plants 2019, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Mahakosee, S.; Jogloy, S.; Vorasoot, N.; Theerakulpisut, P.; Toomsan, B.; Holbrook, C.C.; Kvien, C.K.; Banterng, P. Light Interception and Radiation Use Efficiency of Three Cassava Genotypes with Different Plant Types and Seasonal Variations. Agronomy 2022, 12, 2888. [Google Scholar] [CrossRef]
- Maboko, M.M.; Du Plooy, C.P. Effect of plant spacing on growth and yield of lettuce (Lactuca sativa L.) in a soilless production system. S. African J. Plant Soil 2009, 26, 195–198. [Google Scholar] [CrossRef]
- Maboko, M.M.; Du Plooy, C.P. High-plant density planting of basil (Ocimum basilicum) during summer/fall growth season improves yield in a closed hydroponic system. Acta Agric. Scand. Sect. B Soil Plant Sci. 2013, 63, 748–752. [Google Scholar] [CrossRef]
- Frantz, J.M.; Bugbee, B. Acclimation of Plant Populations to Shade: Photosynthesis, Respiration, and Carbon Use Efficiency. J. Am. Soc. Hortic. Sci. 2005, 130, 918–927. [Google Scholar] [CrossRef]
- Jin, W.; Formiga Lopez, D.; Heuvelink, E.; Marcelis, L.F.M. Light use efficiency of lettuce cultivation in vertical farms compared with greenhouse and field. Food Energy Secur. 2023, 12, e391. [Google Scholar] [CrossRef]
- Carotti, L.; Pistillo, A.; Zauli, I.; Pennisi, G.; Martin, M.; Gianquinto, G.; Orsini, F. Far-red radiation management for lettuce growth: Physiological and morphological features leading to energy optimization in vertical farming. Sci. Hortic. 2024, 334, 113264. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Cycle (DAS) | Planting Density | Fresh Weight (g FW plant−1) | Dry Weight (g DW plant−1) | Plant Height (cm) | Stomatal Conductance (mmol m−2 s−1) |
|---|---|---|---|---|---|
| Lettuce | |||||
| 15 | LD | a 0.84 | a 0.05 | a 5.36 | a 231 |
| MD | a 0.72 | b 0.04 | b 4.77 | a 267 | |
| HD | b 0.41 | c 0.02 | c 4.23 | a 256 | |
| 22 | LD | a 8.01 | a 0.42 | a 8.68 | a 245 |
| MD | b 6.30 | b 0.29 | b 8.05 | a 249 | |
| HD | c 3.88 | c 0.15 | a 8.69 | a 223 | |
| 29 | LD | a 24.3 | a 1.12 | b 11.6 | b 206 |
| MD | b 17.7 | b 0.73 | b 11.5 | ab 221 | |
| HD | c 13.6 | c 0.54 | a 14.8 | a 257 | |
| Basil | |||||
| 15 | LD | a 0.36 | a 0.04 | a 5.47 | - |
| MD | a 0.38 | a 0.04 | a 5.20 | - | |
| HD | b 0.25 | b 0.02 | b 4.20 | - | |
| 22 | LD | a 2.09 | a 0.21 | a 10.6 | a 161 |
| MD | a 2.30 | a 0.21 | a 9.80 | a 145 | |
| HD | b 1.28 | b 0.11 | b 8.07 | a 163 | |
| 29 | LD | a 6.62 | a 1.23 | a 18.6 | a 299 |
| MD | a 6.89 | ab 0.73 | a 17.6 | a 284 | |
| HD | b 4.93 | b 0.54 | a 17.8 | b 246 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jadhav, V.; Grondona, T.; Pistillo, A.; Pennisi, G.; Ghio, M.; Gianquinto, G.; Orsini, F. Optimizing Planting Density for Increased Resource Use Efficiency in Baby-Leaf Production of Lettuce (Lactuca sativa L.) and Basil (Ocimum basilicum L.) in Vertical Farms. Horticulturae 2025, 11, 343. https://doi.org/10.3390/horticulturae11040343
Jadhav V, Grondona T, Pistillo A, Pennisi G, Ghio M, Gianquinto G, Orsini F. Optimizing Planting Density for Increased Resource Use Efficiency in Baby-Leaf Production of Lettuce (Lactuca sativa L.) and Basil (Ocimum basilicum L.) in Vertical Farms. Horticulturae. 2025; 11(4):343. https://doi.org/10.3390/horticulturae11040343
Chicago/Turabian StyleJadhav, Vivek, Tiziano Grondona, Alessandro Pistillo, Giuseppina Pennisi, Marco Ghio, Giorgio Gianquinto, and Francesco Orsini. 2025. "Optimizing Planting Density for Increased Resource Use Efficiency in Baby-Leaf Production of Lettuce (Lactuca sativa L.) and Basil (Ocimum basilicum L.) in Vertical Farms" Horticulturae 11, no. 4: 343. https://doi.org/10.3390/horticulturae11040343
APA StyleJadhav, V., Grondona, T., Pistillo, A., Pennisi, G., Ghio, M., Gianquinto, G., & Orsini, F. (2025). Optimizing Planting Density for Increased Resource Use Efficiency in Baby-Leaf Production of Lettuce (Lactuca sativa L.) and Basil (Ocimum basilicum L.) in Vertical Farms. Horticulturae, 11(4), 343. https://doi.org/10.3390/horticulturae11040343