

Cucurbita ficifolia Rootstock Enhances Resistance to Low-Temperature Stress in Cucumber

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experimental Design
2.3. Determination of Morphological Indicators
2.4. Chlorophyll Determination
2.5. Determination of Electrolyte Leakage Rate and Malondialdehyde Content
2.6. Osmotic Regulatory Substance Content Determination
2.7. Detection of Antioxidant Enzyme Activities
2.8. RNA Extraction and qRT–PCR
2.9. Statistical Analysis
3. Results
3.1. Seedlings with C. ficifolia Rootstock Showed Enhanced Resistance Against Low-Temperature Stress
3.2. C. ficifolia, as Grafting Rootstock, Increased the Light and Pigment Content of Cucumber
3.3. The Content of Osmoregulatory Substances in NH Was Higher than That in ZG
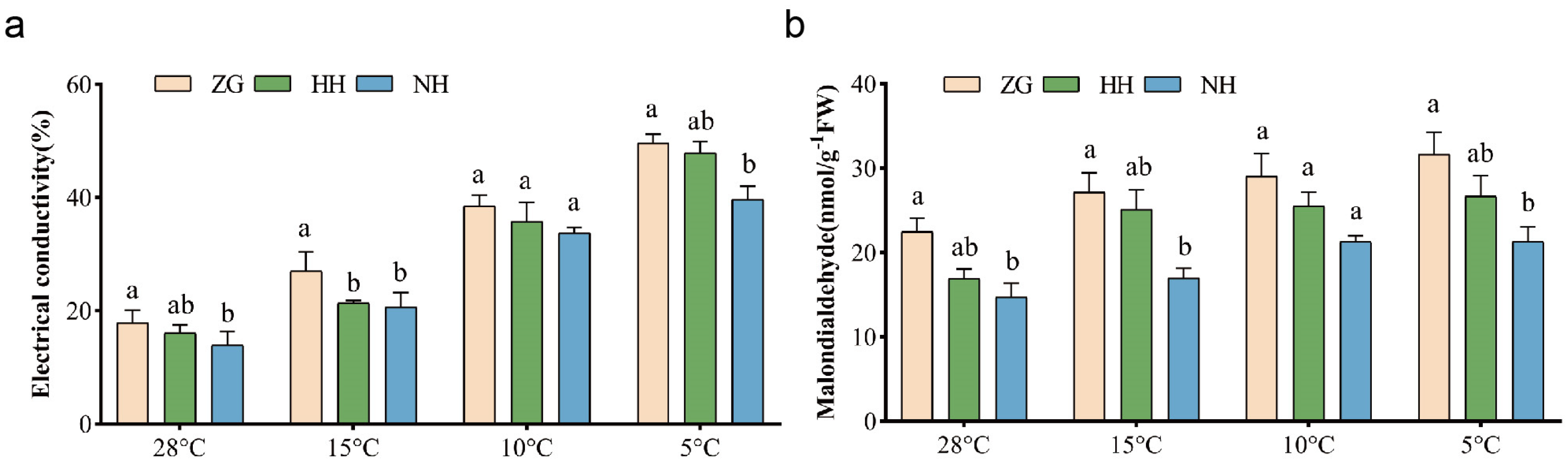
3.4. The Electrical Conductivity Rate and Malondialdehyde Content of NH Were Always Lower than Those of ZG
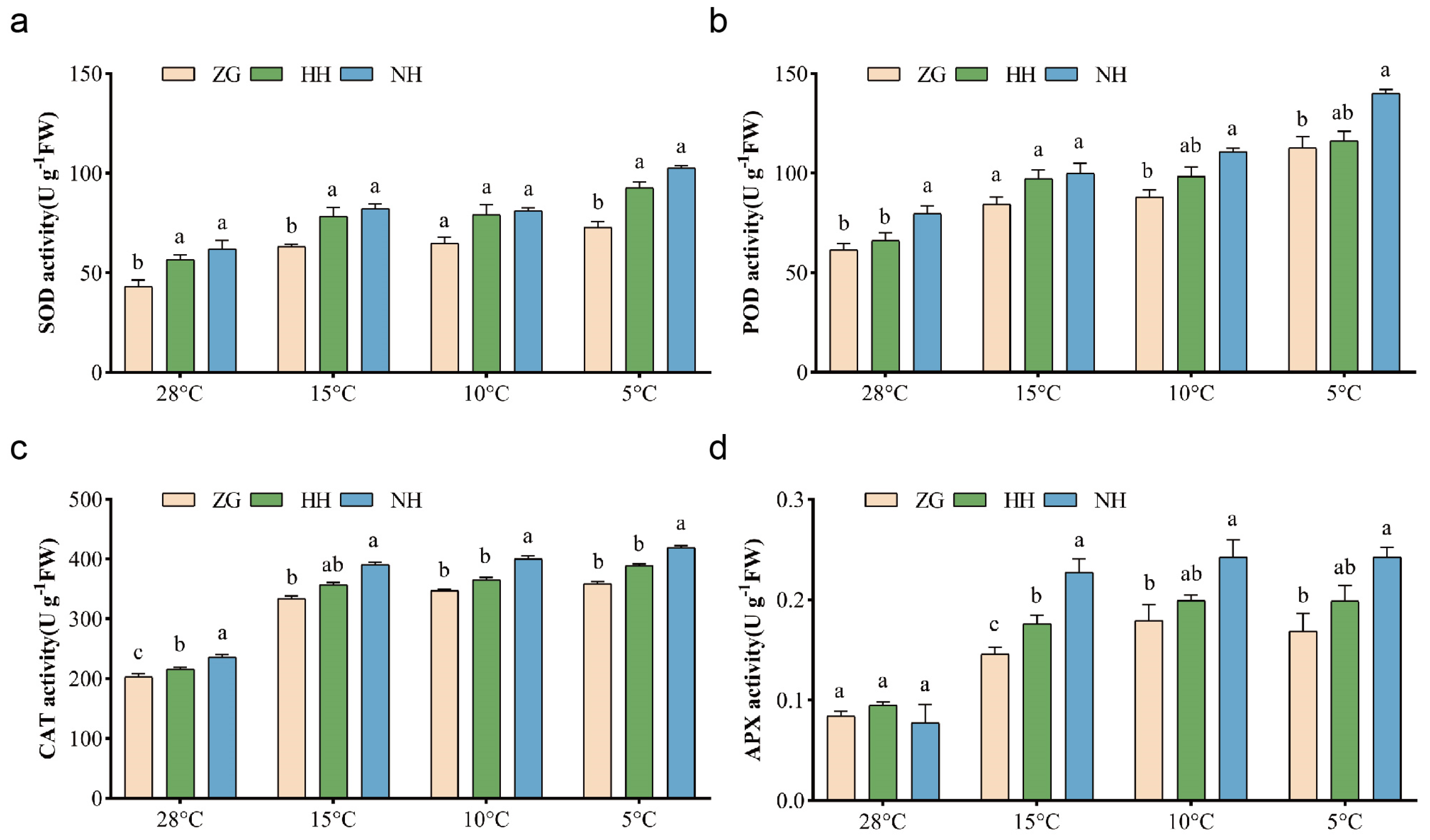
3.5. The Activity of Antioxidant Enzymes Was Significantly Enhanced in NH
3.6. Grafting Resists Oxidative Damage Caused by Up-Regulating the Expression of Antioxidant System Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| EL | Electrolyte leakage |
| MDA | Malondialdehyde |
| Pro | Proline |
| SP | Soluble protein |
| SS | Soluble sugar |
| SOD | Superoxide dismutase |
| POD | Peroxidase |
| CAT | Catalase |
| APX | Ascorbate peroxidase |
References
- Zhang, J.; Feng, S.; Yuan, J.; Wang, C.; Lu, T.; Wang, H.; Yu, C. The formation of fruit quality in Cucumis sativus L. Front. Plant Sci. 2021, 12, 729448. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Low temperature disorders and preventive measures in cucumber. Mod. Rural Technol. 2016, 4, 27. [Google Scholar]
- Wang, Y.; Zhang, F.; Xu, Y.; Chen, Q.; Zhang, H. The main progress in the research on the mechanism and application of cucumber’s low temperature and weak light tolerance. China Veg. 2005, 7–12. [Google Scholar] [CrossRef]
- Cabrera, R.M.; Saltveit, M.E.; Owens, K. Cucumber cultivars differ in their response to chilling temperatures. J. Am. Soc. Hortic. Sci. 1992, 117, 802–807. [Google Scholar] [CrossRef]
- Hao, J.; Li, Y.; Meng, S.; Zhao, B. Effects of night low temperature on sugar accumulation and sugar-metabolizing enzyme activities in melon fruit. Sci. Agric. Sin. 2009, 42, 3592–3599. [Google Scholar]
- Pei, Z.-Q.; Ma, C.; Dong, C.-Y.; Xu, T.-T.; Chai, C.-H.; Zhu, Q.; Wang, J.; Zheng, S.; Zhang, T.-G. Target of rapamycin coordinates auxin are involved in exogenous melatonin regulated low temperature tolerance in cucumber seedlings. Plant Physiol. Biochem. 2024, 215, 109055. [Google Scholar] [CrossRef]
- Jian, L. Advances of the studies on the mechanism of plant cold hardiness. Chin. Bull. Bot. 1992, 9, 17–22+16. [Google Scholar]
- Ando, Y.; Maeda, Y.; Mizutani, K.; Wakatsuki, N.; Hagiwara, S.; Nabetani, H. Impact of blanching and freeze-thaw pretreatment on drying rate of carrot roots in relation to changes in cell membrane function and cell wall structure. LWT-Food Sci. Technol. 2016, 71, 40–46. [Google Scholar] [CrossRef]
- Amin, B.; Atif, M.J.; Kandegama, W.; Nasar, J.; Alam, P.; Fang, Z.; Cheng, Z. Low temperature and high humidity affect dynamics of chlorophyll biosynthesis and secondary metabolites in Cucumber. BMC Plant Biol. 2024, 24, 903. [Google Scholar] [CrossRef] [PubMed]
- Blum, A. Plant Breeding for Stress Environments; CRC Press: Boca Raton, FL, USA, 1988. [Google Scholar]
- Kaplan, F.; Kopka, J.; Haskell, D.W.; Zhao, W.; Schiller, K.C.; Gatzke, N.; Sung, D.Y.; Guy, C.L. Exploring the Temperature-Stress Metabolome of Arabidopsis. Plant Physiol. 2004, 136, 4159–4168. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, F.; Kopka, J.; Sung, D.Y.; Zhao, W.; Popp, M.; Porat, R.; Guy, C.L. Transcript and metabolite profiling during cold acclimation of Arabidopsis reveals an intricate relationship of cold-regulated gene expression with modifications in metabolite content. Plant J. 2007, 50, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Dang, P.; Liu, L.; He, C. Cold acclimation by the CBF–COR pathway in a changing climate: Lessons from Arabidopsis thaliana. Plant Cell Rep. 2019, 38, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Feng, Y.; Jing, T.; Liu, X.; Ai, X.; Bi, H. Melatonin Promotes the Chilling Tolerance of Cucumber Seedlings by Regulating Antioxidant System and Relieving Photoinhibition. Front. Plant Sci. 2021, 12, 789617. [Google Scholar] [CrossRef]
- Pan, D.-Y.; Fu, X.; Zhang, X.-W.; Liu, F.-J.; Bi, H.-G.; Ai, X.-Z. Hydrogen sulfide is required for salicylic acid–induced chilling tolerance of cucumber seedlings. Protoplasma 2020, 257, 1543–1557. [Google Scholar] [CrossRef]
- Nie, W.; Wen, D. Study on the applications and regulatory mechanisms of grafting on vegetables. Plants 2023, 12, 2822. [Google Scholar] [CrossRef] [PubMed]
- Robledo-Torres, V.; González-Cortés, A.; Luna-García, L.R.; Mendoza-Villarreal, R.; Pérez-Rodríguez, M.Á.; Camposeco-Montejo, N. Histological Variations in Cucumber Grafted Plants and Their Effect on Yield. Agronomy 2024, 14, 1377. [Google Scholar] [CrossRef]
- Fu, X.; Lv, C.-Y.; Zhang, Y.-Y.; Ai, X.-Z.; Bi, H.-G. Comparative transcriptome analysis of grafting to improve chilling tolerance of cucumber. Protoplasma 2023, 260, 1349–1364. [Google Scholar] [CrossRef]
- Huang, Y.; Bie, Z.; He, S.; Hua, B.; Zhen, A.; Liu, Z. Improving cucumber tolerance to major nutrients induced salinity by grafting onto Cucurbita ficifolia. Environ. Exp. Bot. 2010, 69, 32–38. [Google Scholar] [CrossRef]
- Zeng, Y.; Zhu, Y.; Wang, B.; Yang, L. Effects of Cucurbita ficifolia as rootstock on growth, fruit setting, disease resistance and leaf nutrient element contents in Cucumis sativus. J. Plant Resour. Environ. 2004, 13, 15–19. [Google Scholar]
- Miao, L.; Li, S.; Bai, L.; Anwar, A.; Li, Y.; He, C.; Yu, X. Effect of grafting methods on physiological change of graft union formation in cucumber grafted onto bottle gourd rootstock. Sci. Hortic. 2019, 244, 249–256. [Google Scholar] [CrossRef]
- Liu, Y.; Yan, Y.; Ma, L.; Cao, D.; Jin, X. Physiological and metabolomic analyses reveal the resistance response mechanism to tea aphid infestation in new shoots of tea plants (Camellia sinensis). Plant Stress 2024, 13, 100545. [Google Scholar] [CrossRef]
- Cheng, H.; Chang, S.; Shi, X.; Chen, Y.; Cong, X.; Cheng, S.; Li, L. Molecular mechanisms of the effects of sodium selenite on the growth, Nutritional quality, and species of organic selenium in Dandelions. Horticulturae 2024, 10, 209. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Gu, W.; Sun, J.; Wang, Y. Regulation of DCPTA treatment on chlorophyll content and fluorescence parameters of maize seedlings leaves under low-temperature stress. Crops 2012, 5, 63–67. [Google Scholar] [CrossRef]
- Zhou, Q.; Wang, J.; Chen, X. Effect of durative low temprature on morphological and physiological characteristics of cucumber seedling. North. Hortic. 2010, 16, 1–3. [Google Scholar]
- Xiong, Z.; Yuan, W.; Chen, X.; Xu, Q.; Chen, C. Influence of sub-low temperature on the growth of cucumber (Cucumis sativus L.) grown in greenhouse. China Veg. 2007, 19–21. [Google Scholar] [CrossRef]
- Jurczyk, B.; Grzesiak, M.; Pociecha, E.; Wlazło, M.; Rapacz, M. Diverse stomatal behaviors mediating photosynthetic acclimation to Low temperatures in hordeum vulgare. Front. Plant Sci. 2019, 9, 1963. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Lou, Q.; Chen, J. A review on chilling injury and cold tolerance in Cucumis sativus L. Chin. Bull. Bot. 2004, 21, 578–586. [Google Scholar]
- Ren, J.; Huang, Z.; Zeng, L.; Shi, Z. Review of physiological reaction mechanism of plants exposed to low temperature stress. World For. Res. 2013, 26, 15–20. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, X.; Zhang, Y.; Wei, M. Effects of chlorophyllin-iron on osmotic adjustment and activities of antioxidantive enzymes in cucumber seedlings under suboptimal temperature. Chin. J. Appl. Ecol. 2014, 25, 3527–3532. [Google Scholar] [CrossRef]
- Xu, X. Study on the Physiological Response and Molecular Mechanism of Grafted Cucumber Seedlings Under Low Temperature Stress. Ph.D. Thesis, Gansu Agricultural University, Lanzhou, China, 2016. [Google Scholar]
- Dat, J.; Vandenabeele, S.; Vranová, E.; Van Montagu, M.; Inzé, D.; Van Breusegem, F. Dual action of the active oxygen species during plant stress responses. Cell. Mol. Life Sci. 2000, 57, 779–795. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, T.; Gao, X. Active oxygen metabolism and ascorbate-glutathione cycle of tomato leaves under low nocturnal temperature. Acta Bot. Boreali-Occident. Sin. 2011, 31, 707–714. [Google Scholar]
- Yu, J.; Zhang, G.; Feng, Z.; Li, X. Effects of low temperature and weak light on Anti-oxidative enzyme activities and Plasm- membrane permeability of pepper seedlings. Acta Bot. Boreali-Occident. Sin. 2005, 25, 2478–2483. [Google Scholar]
- Wang, H.; Ai, X.; Zheng, N.; Jiang, F. Effects of graft on lipid peroxidation and antioxidative enzyme activities of Capsicum annum seedlings under low temperature and weak light intensity. Chin. J. Appl. Ecol. 2010, 21, 1289–1294. [Google Scholar]
- Erich, F. Oxygen activation and oxygen toxicity. Annu. Rev. Plant Physiol. 1982, 33, 73–96. [Google Scholar] [CrossRef]
- Bowler, C.; Vanmontagu, M.; Inze, D. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Wang, R.; Shi, L.; Tang, G.; Liang, Y. Effect of osmotic stress on activities of protective enzymes system in Agropyron mongolicum seedling. Chin. Bull. Bot. 2003, 20, 330–335. [Google Scholar]
- Yang, S.; Chen, X.; Hui, W.; Ren, Y. Progress in responses of antioxidant enzyme systems in plant to environmental stresses. J. Fujian Agric. For. Univ. 2016, 45, 481–489. [Google Scholar]
- Zhang, Z.; Liu, S.; Wang, Z.; Zhang, Q.; Meng, F.; Yu, X. Effect of low temperature stress on physiological and biochemical indices of cucumber seedlings grafted on different rootstocks. Shandong Agric. Sci. 2009, 36–40. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, P.; Huang, M.; Chen, H. Effects of high temperature on the activity and expression of antioxidative enzymes in rice flag leaves during the flowering stage. Plant Sci. J. 2015, 33, 355–361. [Google Scholar]
- Ma, D.; Sun, Q. Effects of temperature stress on membrane protective system of cucumber seedlings. Acta Bot. Boreali-Occident. Sin. 2001, 656–661. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wenzel, O.; Buschmann, C.; Gitelson, A. Plant Stress Detection by Reflectance and Fluorescence. Ann. N. Y. Acad. Sci. 1998, 851, 271–285. [Google Scholar] [CrossRef]
- Miller, A.R.; Kelley, T.J. Mechanical Stress Stimulates Peroxidase Activity in Cucumber Fruit. HortScience 1989, 24, 650–652. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High Salinity Induces Different Oxidative Stress and Antioxidant Responses in Maize Seedlings Organs. Front. Plant Sci. 2016, 7, 276. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Time | Temperature |
|---|---|---|
| self-rooted seedling (ZG) | 7 d | 28 °C |
| self-grafted seedling (HH) | 7 d | 28 °C |
| C. ficifolia-grafted (NH) | 7 d | 28 °C |
| self-rooted seedling (ZG) | 7 d | 15 °C |
| self-grafted seedling (HH) | 7 d | 15 °C |
| C. ficifolia-grafted (NH) | 7 d | 15 °C |
| self-rooted seedling (ZG) | 7 d | 10 °C |
| self-grafted seedling (HH) | 7 d | 10 °C |
| C. ficifolia-grafted (NH) | 7 d | 10 °C |
| self-rooted seedling (ZG) | 7 d | 5 °C |
| self-grafted seedling (HH) | 7 d | 5 °C |
| C. ficifolia-grafted (NH) | 7 d | 5 °C |
| Gene | Forward Primer Sequence | Reverse Primer Sequence |
|---|---|---|
| SOD | AGCACCAGGCATCCTTACTT | GAACCTCCAATGTCCTCCGA |
| POD | ATGGGTCGGTGCTAC | CTGATGCCAAGGTGA |
| CAT | CGGAGTGGAGGCTTTA | CGGCACCAACGACAT |
| APX | TGGTGTTGTTGCTGTTGAGG | GCTCGGGTTTGTCCTCTCTA |
| Actin | AGCCATCTCTCATCGGTAT | CATGGTTGAACCACCACTG |
| Treatment | Fresh Weight (g) | Dry-Weight (g) | Plant Height (cm) | Stem Diameter (mm) |
|---|---|---|---|---|
| 28 °C-ZG | 23.10 ± 0.45 a | 2.17 ± 0.020 ab | 15.30 ± 0.44 a | 5.12 ± 0.071 a |
| 28 °C-HH | 23.47 ± 0.35 a | 2.23 ± 0.026 a | 14.97 ± 0.18 a | 5.17 ± 0.112 a |
| 28 °C-NH | 23.57 ± 0.33 a | 2.27 ± 0.023 a | 14.73 ± 0.19 ab | 5.17 ± 0.122 a |
| 15 °C–ZG | 18.90 ± 0.53 c | 1.76 ± 0.046 d | 14.00 ± 0.20 cd | 4.86 ± 0.080 bc |
| 15 °C-HH | 20.77 ± 0.34 b | 2.01 ± 0.026 c | 14.23 ± 0.26 bc | 4.93 ± 0.045 b |
| 15 °C-NH | 21.13 ± 0.71 b | 2.05 ± 0.101 bc | 13.67 ± 0.15 cd | 4.89 ± 0.026 bc |
| 10 °C-ZG | 17.37 ± 0.62 def | 1.67 ± 0.055 de | 13.43 ± 0.12 d | 4.63 ± 0.020 cd |
| 10 °C-HH | 17.93 ± 0.44 cd | 1.69 ± 0.023 de | 12.77 ± 0.09 e | 4.65 ± 0.028 de |
| 10 °C-NH | 17.53 ± 0.48 cde | 1.67 ± 0.057 de | 12.53 ± 0.09 ef | 4.71 ± 0.020 cd |
| 5 °C-ZG | 15.87 ± 0.18 f | 1.48 ± 0.024 f | 12.57 ± 0.19 e | 4.38 ± 0.015 e |
| 5 °C-HH | 16.40 ± 0.57 ef | 1.63 ± 0.039 de | 12.17 ± 0.23 ef | 4.48 ± 0.036 ef |
| 5 °C-NH | 16.30 ± 0.50 ef | 1.56 ± 0.030 ef | 11.90 ± 0.12 f | 4.47 ± 0.059 ef |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, H.; Deng, W.; Ahmad, B.; Guo, C.; Shi, S.; He, S.; Yang, Z. Cucurbita ficifolia Rootstock Enhances Resistance to Low-Temperature Stress in Cucumber. Horticulturae 2025, 11, 242. https://doi.org/10.3390/horticulturae11030242
Xiao H, Deng W, Ahmad B, Guo C, Shi S, He S, Yang Z. Cucurbita ficifolia Rootstock Enhances Resistance to Low-Temperature Stress in Cucumber. Horticulturae. 2025; 11(3):242. https://doi.org/10.3390/horticulturae11030242
Chicago/Turabian StyleXiao, Haojun, Wenxin Deng, Bilal Ahmad, Chunmei Guo, Songmei Shi, Shuilian He, and Zheng’an Yang. 2025. "Cucurbita ficifolia Rootstock Enhances Resistance to Low-Temperature Stress in Cucumber" Horticulturae 11, no. 3: 242. https://doi.org/10.3390/horticulturae11030242
APA StyleXiao, H., Deng, W., Ahmad, B., Guo, C., Shi, S., He, S., & Yang, Z. (2025). Cucurbita ficifolia Rootstock Enhances Resistance to Low-Temperature Stress in Cucumber. Horticulturae, 11(3), 242. https://doi.org/10.3390/horticulturae11030242