

Advancing Glycyrrhiza glabra L. Cultivation and Hairy Root Transformation and Elicitation for Future Metabolite Overexpression
 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material, Seed Germination, and Growth Conditions
2.2. Induction and Proliferation of Hairy Root Cultures
2.3. mRNA, cDNA Synthesis, and PCR Verification of Transformed Roots
2.4. Growth Curve of Hairy Roots
2.5. Light Induction Experiment
2.6. Ferric Reducing Antioxidant Power (FRAP) Assay
2.7. Total Phenolic Content (TPC) Assay
2.8. Statistical Analyses
3. Results
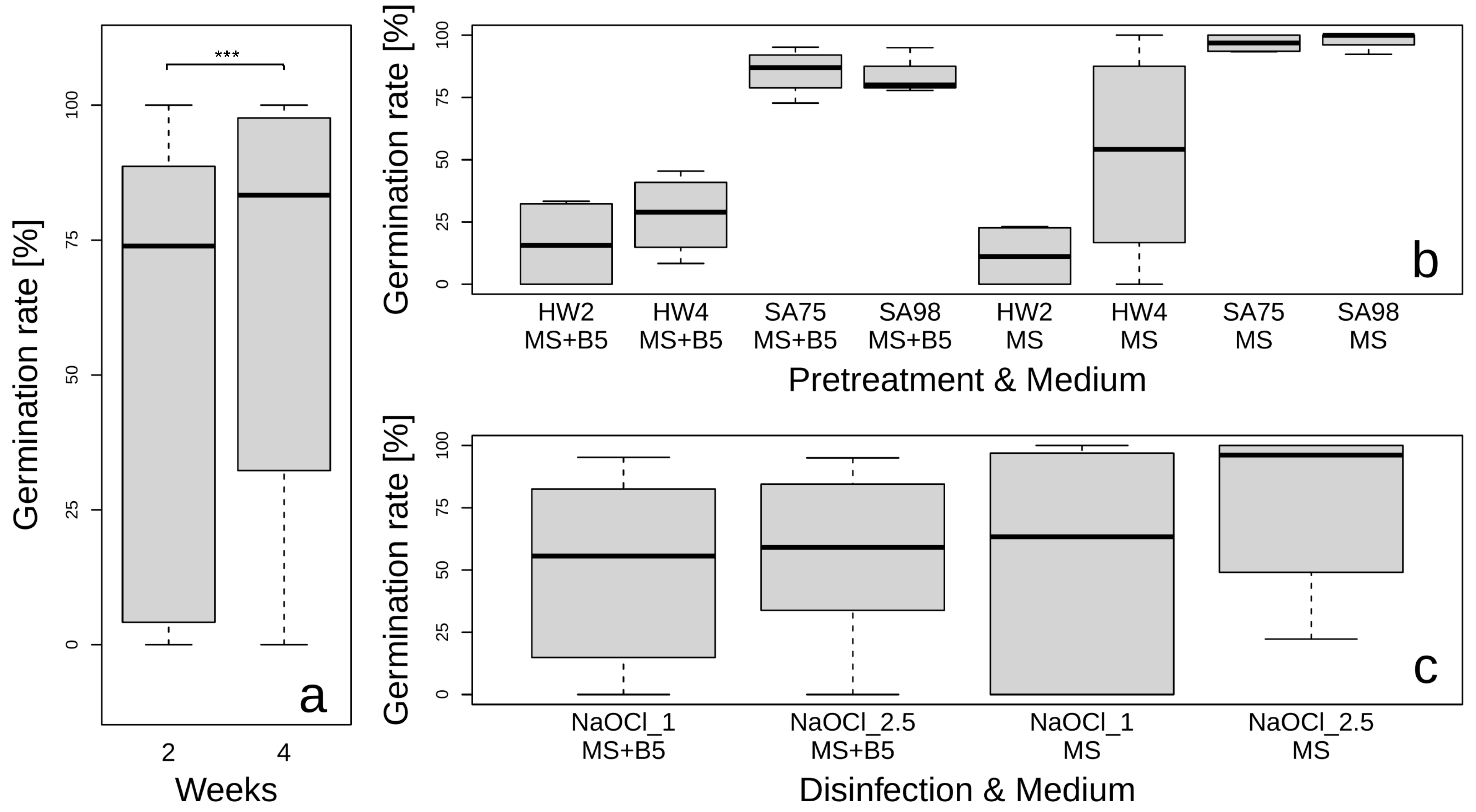
3.1. Pretreatment, Sterilization, and Growth Media Effects on Seed Germination in G. glabra
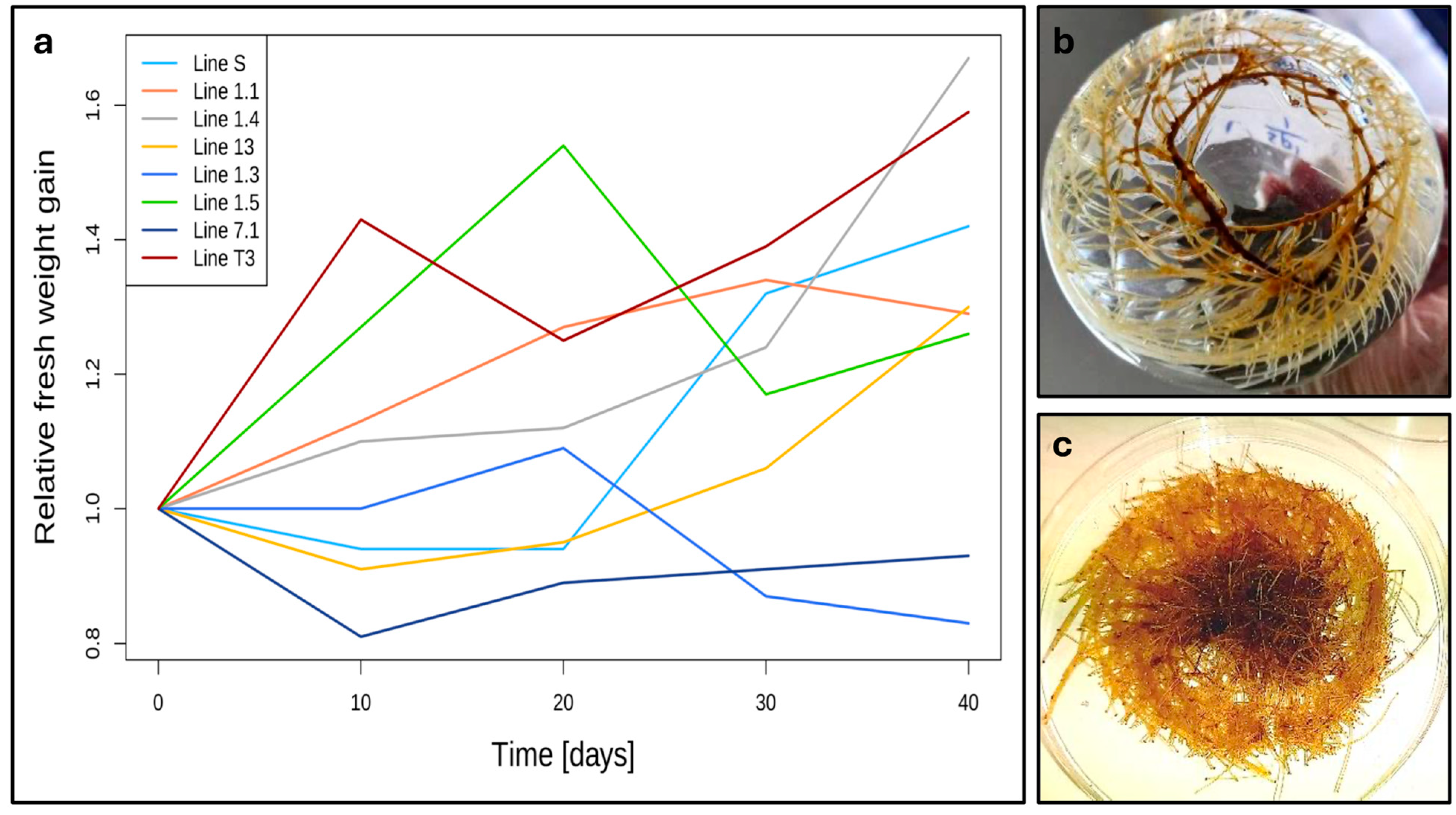
3.2. Optimal Conditions for Induction and Proliferation of Hairy Root Cultures in G. glabra
3.3. Growth Rate of Hairy Root Lines Showing Optimal Growth
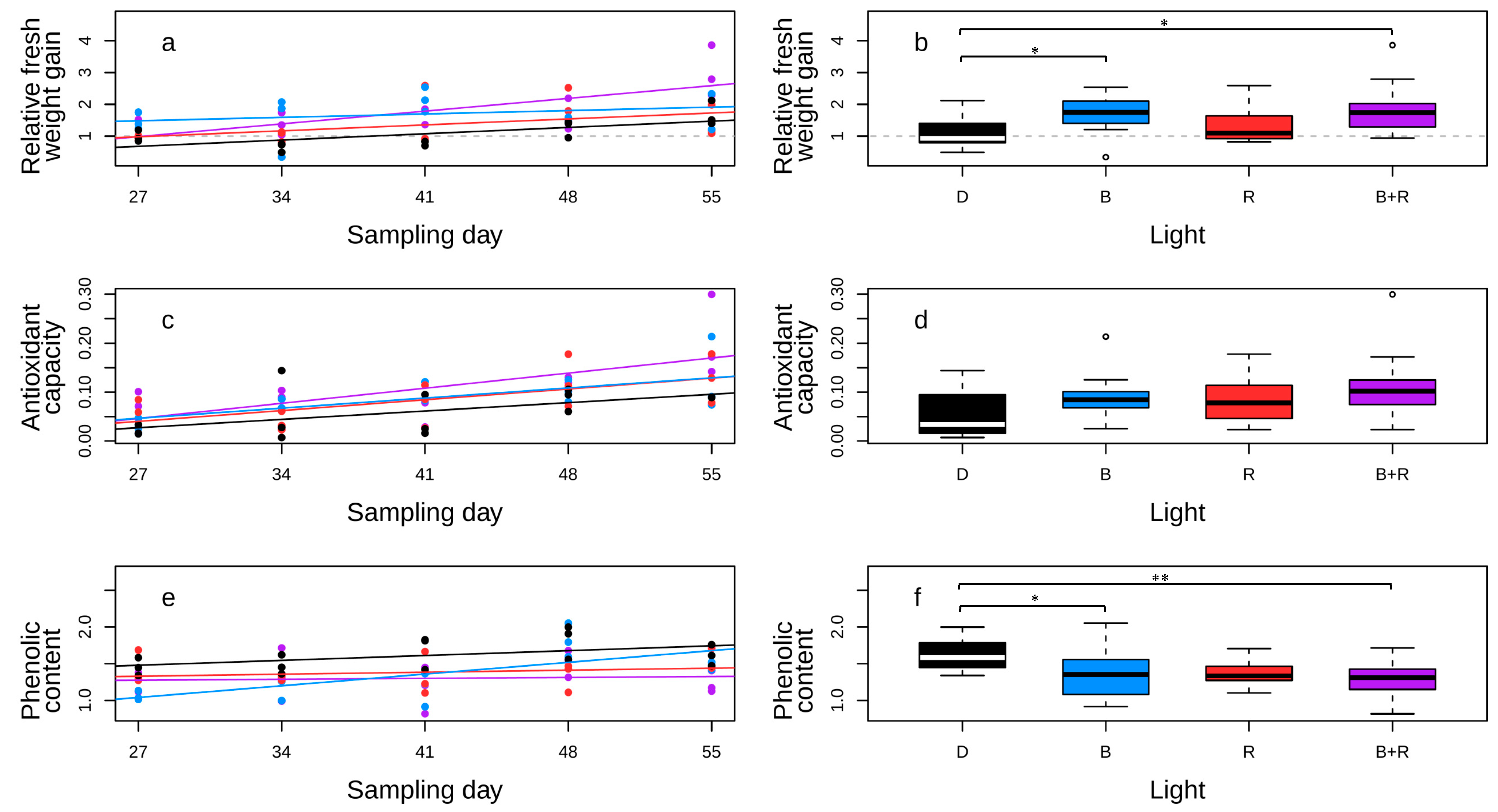
3.4. The Effect of Light Quality on the Growth Rate, FRAP, and Phenolic Content of Hairy Root Line S
4. Discussion
4.1. Effect of Pretreatments, Disinfection, and Media on Germination Rate of G. glabra
4.2. Effect of R. rhizogenes Strain on Hairy Root Transformation Rate of G. glabra
4.3. Effect of Explant Type and Infection Method on Hairy Root Transformation Rate
4.4. Testing Light Induction on the Growth of G. Glabra Hairy Root Line S
4.5. Enhancement of Antioxidant Capacity, as Measured by FRAP, in G. glabra Hairy Root Line S Exposed to Different Qualities of Light
4.6. Determining the Correlation Between FRAP and TPC
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yeskaliyeva, B.; Kırkın, C. Glycyrrhiza Genus: Enlightening Phytochemical Components for Pharmacological and Health-Promoting Abilities. Oxidative Med. Cell. Longev. 2021, 2021, 7571132. [Google Scholar]
- Abraham, J.; Florentine, S. Licorice (Glycyrrhiza glabra) Extracts-Suitable Pharmacological Interventions for COVID-19? A Review. Plants 2021, 10, 2600. [Google Scholar] [CrossRef]
- Yang, R.; Yuan, B.-C.; Ma, Y.-S.; Zhou, S.; Liu, Y. The Anti-Inflammatory Activity of Licorice, a Widely Used Chinese Herb. Pharm. Biol. 2017, 55, 5–18. [Google Scholar] [CrossRef]
- Li, Y.-J.; Chen, J.; Li, Y.; Li, Q.; Zheng, Y.-F.; Fu, Y.; Li, P. Screening and Characterization of Natural Antioxidants in Four Glycyrrhiza Species by Liquid Chromatography Coupled with Electrospray Ionization Quadrupole Time-of-Flight Tandem Mass Spectrometry. J. Chromatogr. A 2011, 1218, 8181–8191. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, L.; Xu, C.; Shi, J.; Chen, S.; Tan, M.; Chen, J.; Zou, L.; Chen, C.; Liu, Z.; et al. A Comprehensive Review for Phytochemical, Pharmacological, and Biosynthesis Studies on Glycyrrhiza spp. The Am. J. Chin. Med. 2020, 48, 17–45. [Google Scholar] [CrossRef]
- Hayashi, H.; Hiraoka, N.; Ikeshiro, Y.; Yamamoto, H. Organ Specific Localization of Flavonoids in Glycyrrhiza glabra L. Plant Sci. 1996, 116, 233–238. [Google Scholar] [CrossRef]
- Wang, Y.-C.; Yang, Y.-S. Simultaneous Quantification of Flavonoids and Triterpenoids in Licorice Using HPLC. J. Chromatogr. B 2007, 850, 392–399. [Google Scholar] [CrossRef]
- Song, W.; Qiao, X.; Chen, K.; Wang, Y.; Ji, S.; Feng, J.; Li, K.; Lin, Y.; Ye, M. Biosynthesis-Based Quantitative Analysis of 151 Secondary Metabolites of Licorice to Differentiate Medicinal Glycyrrhiza Species and Their Hybrids. Anal. Chem. 2017, 89, 3146–3153. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Sudo, H. Economic Importance of Licorice. Plant Biotechnol. 2009, 26, 101–104. [Google Scholar] [CrossRef]
- Khaitov, B.; Karimov, A.; Khaitbaeva, J.; Sindarov, O.; Karimov, A.; Li, Y. Perspectives of Licorice Production in Harsh Environments of the Aral Sea Regions. Int. J. Environ. Res. Public Health 2022, 19, 11770. [Google Scholar] [CrossRef]
- Gemedzhieva, N.; Khrokov, A.; Heral, E.; Timoshyna, A. Sweet dreams: Assessing opportunities and threats in Kazakhstan’s wild liquorice root trade. TRAFFIC 2021. Available online: https://www.traffic.org/site/assets/files/14086/sweet-dreams-en-final.pdf (accessed on 17 December 2024).
- Li, B.; Bhandari, D.R.; Janfelt, C.; Römpp, A.; Spengler, B. Natural Products in Glycyrrhiza Glabra (Licorice) Rhizome Imaged at the Cellular Level by Atmospheric Pressure Matrix-assisted Laser Desorption/Ionization Tandem Mass Spectrometry Imaging. Plant J. 2014, 80, 161–171. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Tani, T. Growth and Glycyrrhizin Contents in Glycyrrhiza uralensis Roots Cultivated for Four Years in Eastern Nei-Meng-gu of China. J. Tradit. Med. 2002, 19, 87–92. [Google Scholar]
- Zhu, S.; Sugiyama, R.; Batkhuu, J.; Sanchir, C.; Zou, K.; Komatsu, K. Survey of Glycyrrhiza radix Resources in Mongolia: Chemical Assessment of the Underground Part of Glycyrrhiza uralensis and Comparison with Chinese Glycyrrhizea radix. J. Nat. Medicines. 2009, 63, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, H.; Mirjalili, M.H.; Karami, A.; Nejad Ebrahimi, S. Introducing the Glycyrrhizic Acid and Glabridin Rich Genotypes from the Cultivated Iranian Licorice (Glycyrrhiza glabra L.) Populations to Exploit in Production Systems. Sci. Rep. 2024, 14, 11034. [Google Scholar] [CrossRef]
- Global Market Insights: Licorice Extract Market Size. Report ID: GMI8220. Available online: https://www.gminsights.com/industry-analysis/licorice-extract-market (accessed on 7 December 2024).
- Applequist, W.L.; Brinckmann, J.A.; Cunningham, A.B.; Hart, R.E.; Heinrich, M.; Katere, D.R.; van Andel, T. Scientists’ Warning on Climate Change and Medicinal Plants. Planta Medica 2020, 86, 10–18. [Google Scholar] [CrossRef]
- Sathasivam, R.; Choi, M.; Radhakrishnan, R.; Kwon, H.; Yoon, J.; Yang, S.H.; Kim, J.K.; Chung, Y.S.; Park, S.U. Effects of Various Agrobacterium rhizogenes Strains on Hairy Root Induction and Analyses of Primary and Secondary Metabolites in Ocimum basilicum. Front. Plant Sci. 2022, 13, 983776. [Google Scholar] [CrossRef]
- Shirazi, Z.; Aalami, A.; Tohidfar, M.; Sohani, M.M. Metabolic Engineering of Glycyrrhizin Pathway by Over-Expression of Beta-Amyrin 11-Oxidase in Transgenic Roots of Glycyrrhiza glabra. Mol. Biotechnol. 2018, 60, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Wongwicha, W.; Tanaka, H.; Shoyama, Y.; Putalun, W. Methyl Jasmonate Elicitation Enhances Glycyrrhizin Production in Glycyrrhiza inflata Hairy Roots Cultures. Z. Für Naturforschung C 2011, 66, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Toivonen, L.; Rosenqvist, H. Establishment and Growth Characteristics of Glycyrrhiza glabra Hairy Root Cultures. Plant Cell Tissue Organ Cult. 1995, 41, 249–258. [Google Scholar] [CrossRef]
- Srivastava, M.; Singh, G.; Sharma, S.; Shukla, S.; Misra, P. Elicitation Enhanced the Yield of Glycyrrhizin and Antioxidant Activities in Hairy Root Cultures of Glycyrrhiza glabra L. J. Plant Growth Regul. 2019, 38, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Singh, S.K.; Lyu, R.; Pattanaik, S.; Wang, Y.; Li, Y.; Yuan, L.; Liu, Y. Metabolic Engineering to Enhance the Accumulation of Bioactive Flavonoids Licochalcone A and Echinatin in Glycyrrhiza inflata (Licorice) Hairy Roots. Front. Plant Sci. 2022, 13, 932594. [Google Scholar] [CrossRef] [PubMed]
- Chiyo, N.; Seki, H.; Kanamoto, T.; Ueda, H.; Kojoma, M.; Muranaka, T. Glycyrrhizin Production in Licorice Hairy Roots Based on Metabolic Redirection of Triterpenoid Biosynthetic Pathway by Genome Editing. Plant Cell Physiol. 2024, 65, 185–198. [Google Scholar] [CrossRef]
- Jiao, J.; Fu, J.-X.; Yao, L.; Gai, Q.-Y.; He, X.-J.; Feng, X.; Fu, Y.-J. The Growth, Adventitious Bud Formation, Bioactive Flavonoid Production, Antioxidant Response, and Cryptochrome-Mediated Light Signal Transduction in Isatis tinctoria L. Hairy Root Cultures Exposed to LED Lights. Ind. Crops Prod. 2023, 195, 116496. [Google Scholar] [CrossRef]
- Gai, Q.-Y.; Feng, X.; Jiao, J.; Xu, X.-J.; Fu, J.-X.; He, X.-J.; Fu, Y.-J. Blue LED Light Promoting the Growth, Accumulation of High-Value Isoflavonoids and Astragalosides, Antioxidant Response, and Biosynthesis Gene Expression in Astragalus membranaceus (Fisch.) Bunge Hairy Root Cultures. Plant Cell Tissue Organ Cult. (PCTOC) 2023, 153, 511–523. [Google Scholar] [CrossRef]
- Yang, Y.; Hou, S.; Fan, W.; Lilan, L.; Hui, N.; Xia, W.; Wei, J. Expression Patterns of Some Genes Involved in Tanshinone Biosynthesis in Salvia miltiorrhiza Roots. Ind. Crops Prod. 2019, 130, 606–614. [Google Scholar] [CrossRef]
- Afreen, F.; Zobayed, S.; Kozai, T. Spectral Quality and UV-B Stress Stimulate Glycyrrhizin Concentration of Glycyrrhiza uralensis in Hydroponic and Pot System. Plant Physiol. Biochem. 2005, 43, 1074–1081. [Google Scholar] [CrossRef] [PubMed]
- Afreen, F.; Zobayed, S.M.; Kozai, T. Melatonin in Glycyrrhiza uralensis: Response of Plant Roots to Spectral Quality of Light and UV-B Radiation. J. Pineal Res. 2006, 41, 108–115.27. [Google Scholar] [CrossRef] [PubMed]
- Ghadiri, H.; Bagherani, T.N. Effects of Scarification and Temperature on Germination of Licorice (Glycyrrhiza glabra L.). Seeds. J. Agric. Sci. Technol. 2000, 2, 257–262. [Google Scholar]
- Velempini, P.; Riddoch, I.; Batisani, N. Seed Treatments for Enhancing Germination of Wild Okra (Corchorus olitorius). Exp. Agric. 2003, 39, 441–447. [Google Scholar] [CrossRef]
- Han, Y.; Hou, Z.; Zhang, X.; Yan, K.; Liang, Z.; He, Q. Important Changes in Germination, Seedling Tolerance, and Active Components Content Due to Drought Stress on Three Licorice (Glycyrrhiza) Species. Ind. Crops Prod. 2022, 175, 114240. [Google Scholar] [CrossRef]
- Mao, P.; Wang, Y.; Wang, X.; Lian, J.; Huang, Y. Conditions and Stimulation for Germination in Glycyrrhiza uralensis Fisch Seeds. Agric. Sci. China 2008, 7, 1438–1444. [Google Scholar] [CrossRef]
- Muhammad, S.; Amusa, N.A. Effects of Sulphuric Acid and Hot Water Treatments on Seed Germination of Tamarind (Tamarindus indica L). Afr. J. Biotechnol. 2003, 2, 279–376. [Google Scholar]
- Msanga, H.P.; Maghembe, J.A. Effect of Hot Water and Chemical Treatments on the Germination of Albizia schimperana Seed. For. Ecol. Manag. 1986, 17, 137–146. [Google Scholar] [CrossRef]
- Müller, L.; Fröhlich, K.; Böhm, V. Comparative Antioxidant Activities of Carotenoids Measured by Ferric Reducing Antioxidant Power (FRAP), ABTS Bleaching Assay (αTEAC), DPPH Assay and Peroxyl Radical Scavenging Assay. Food Chem. 2011, 129, 139–148. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing 2024. Available online: https://www.R-project.org/ (accessed on 18 October 2024).
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models 2022. Available online: https://CRAN.R-project.org/package=DHARMa (accessed on 18 October 2024).
- Kozhuharova, A.; Nikolova, M.; Stoyanov, S.; Yankova-Tsvetkova, E.; Ilinkin, V.; Berkov, S.; Stanilova, M. Establishment of an Ex Situ Collection of Glycyrrhiza glabra L. as a Prerequisite for Field Cultivation in Bulgaria. Horticulturae 2024, 10, 86. [Google Scholar] [CrossRef]
- Seng, M.; Cheong, E.J. Comparative Study of Various Pretreatment on Seed Germination of Dalbergia cochinchinensis. For. Sci. Technol. 2020, 16, 68–74. [Google Scholar] [CrossRef]
- Cavallaro, V.; Maucieri, C.; Patanè, C.; Fascella, G.; Pellegrino, A.; Barbera, A.C. Polyphenols Leaching and Seed Dormancy in Carob (Ceratonia siliqua L.) in Relation to Hot Water Treatment. Acta Physiol. Plant. 2021, 43, 141. [Google Scholar] [CrossRef]
- Wang, Y.; Hanson, J.; Mariam, Y. Breaking Hard Seed Dormancy in Diverse Accessions of Five Wild Vigna Species by Hot Water and Mechanical Scarification. Seed Sci. Technol. 2011, 39, 12–20. [Google Scholar] [CrossRef]
- Boyle, T.H.; Hladun, K. Influence of Seed Size, Testa Color, Scarification Method, and Immersion in Cool or Hot Water on Germination of Baptisia australis (L.) R. Br. Seeds. HortScience 2005, 40, 1846–1849. [Google Scholar] [CrossRef]
- Neri-Soto, A.V.; Chávez-Simental, J.A.; Corral-Rivas, J.J.; Montiel-Antuna, E.; Rubio-Graciano, R.B. In Vitro Germination of Picea chihuahuana in Different Culture Media and Light Conditions. In Vitro 2015, 2, 3246–3250. [Google Scholar]
- Kovalenko, P.G.; Maliuta, S.S. Effect of the Elicitors on Secondary Metabolites Production by Licorice (Glycyrrhiza uralensis Fisch.) Hairy Roots Cultures. Ukr. Bioorganica Acta. 2003, 1, 50–60. [Google Scholar]
- Zhang, H.-C.; Liu, J.-M.; Lu, H.-Y.; Gao, S.-L. Enhanced Flavonoid Production in Hairy Root Cultures of Glycyrrhiza uralensis Fisch by Combining the Over-Expression of Chalcone Isomerase Gene with the Elicitation Treatment. Plant Cell Rep. 2009, 28, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Shirazi, Z.; Piri, K.; Asl, A.M.; Hasanloo, T. Glycyrrhizin and Isoliquiritigenin Production by Hairy Root Culture of Glycyrrhiza glabra. J. Med. Plants Res. 2012, 6, 4640–4646.45. [Google Scholar] [CrossRef]
- Zhang, H.-C.; Liu, J.-M.; Chen, H.-M.; Gao, C.-C.; Lu, H.-Y.; Zhou, H.; Li, Y.; Gao, S.-L. Up-Regulation of Licochalcone A Biosynthesis and Secretion by Tween 80 in Hairy Root Cultures of Glycyrrhiza uralensis Fisch. Mol. Biotechnol. 2011, 47, 50–56. [Google Scholar] [CrossRef]
- Hamill, J.D.; Lidgett, A.J. Hairy Root Cultures—Opportunities and Key Protocols for Studies in Metabolic Engineering. In Hairy Roots; CRC Press: Boca Raton, FL, USA, 2019; pp. 1–30. [Google Scholar]
- Hakeem, K.R.; Ahmad, P.; Ozturk, M. Crop Improvement: New Approaches and Modern Techniques, 1st ed.; Springer Science & Business Media: New York, NY, USA, 2013. [Google Scholar]
- Montazeri, M.; Pakdin-Parizi, A.; Najafi-Zarrini, H.; Azadbakht, M.; Nematzadeh, G.; Gholami, Z. A Comparative Analysis of the Hairy Root Induction Methods in Hypericum perforatum. J. Plant Mol. Breed. 2019, 7, 67–76. [Google Scholar] [CrossRef]
- Triplett, B.A.; Moss, S.C.; Bland, J.M.; Dowd, M.K. Induction of Hairy Root Cultures from Gossypium hirsutum and Gossypium barbadense to Produce Gossypol and Related Compounds. Vitr. Cell Dev. Biol. Plant 2008, 44, 508–517. [Google Scholar] [CrossRef]
- Xu, T.; Zhang, L.; Sun, X.; Zhang, H.; Tang, K. Production and Analysis of Organic Acids in Hairy-root Cultures of Isatis indigotica Fort.(Indigo Woad). Biotechnol. Appl. Biochem. 2004, 39, 123–128. [Google Scholar] [CrossRef]
- Cui, M.-L.; Liu, C.; Piao, C.-L.; Liu, C.-L. A Stable Agrobacterium rhizogenes-Mediated Transformation of Cotton (Gossypium hirsutum L.) and Plant Regeneration from Transformed Hairy Root via Embryogenesis. Front. Plant Sci. 2020, 11, 604255. [Google Scholar] [CrossRef]
- Chen, G.J.; Lee, M.-S.; Lin, M.-K.; Ko, C.-Y.; Chang, W.-T. Blue Light Decreases Tanshinone IIA Content in Salvia miltiorrhiza Hairy Roots via Genes Regulation. J. Photochem. Photobiol. B Biol. 2018, 183, 164–171. [Google Scholar] [CrossRef]
- Wongshaya, P.; Chayjarung, P.; Tothong, C.; Pilaisangsuree, V.; Somboon, T.; Kongbangkerd, A.; Limmongkon, A. Effect of Light and Mechanical Stress in Combination with Chemical Elicitors on the Production of Stilbene Compounds and Defensive Responses in Peanut Hairy Root Culture. Plant Physiol. Biochem. 2020, 157, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.W.; Lee, J.G.; Yoo, J.H.; Lim, J.D.; Choi, I.Y.; Kim, M.J.; Yu, C.Y.; Seong, E.S. Cellular Morphology and Transcriptome Comparative Analysis of Astragalus membranaceus Bunge Sprouts Cultured in Vitro under Different LED Light. Plants 2023, 12, 1914. [Google Scholar] [CrossRef] [PubMed]
- Sani, S.; Abushattal, M.A.K.; Subramaniam, S.; Hassan, N.H.; Mad’Atari, M.F. Enhancing Bioactive Compounds in Hairy Roots Culture of Precious Medicinal Plant Eurycoma longifolia Jack. through LED Elicitation. Plant Cell Tissue Organ Cult. (PCTOC) 2024, 158, 62. [Google Scholar] [CrossRef]
- Gupta, M.; Karmakar, N.; Sasmal, S.; Chowdhury, S.; Biswas, S. Free Radical Scavenging Activity of Aqueous and Alcoholic Extracts of Glycyrrhiza glabra Linn. Measured by Ferric Reducing Antioxidant Power (FRAP), ABTS Bleaching Assay (αTEAC), DPPH Assay and Peroxyl Radical Antioxidant Assay. Int. J. Pharmacol. Toxicol. 2016, 4, 235–240. [Google Scholar] [CrossRef]
- Kim, H.J.; Seo, J.-Y.; Suh, H.-J.; Lim, S.S.; Kim, J.-S. Antioxidant Activities of Licorice-Derived Prenylflavonoids. Nutr. Res. Pract. 2012, 6, 491–498. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Devaki, M. The Ferric Reducing/Antioxidant Power (FRAP) Assay for Non-enzymatic Antioxidant Capacity: Concepts, Procedures, Limitations and Applications. In Measurement of Antioxidant Activity & Capacity: Recent Trends and Applications; Wiley: Hoboken, NJ, USA, 2018; pp. 77–106. [Google Scholar] [CrossRef]
- Ohashi, H.; Kyogoku, T.; Ishikawa, T.; Kawase, S.; Kawai, S. Antioxidative Activity of Tree Phenolic Constituents 1: Radical-Capturing Reaction of Flavon-3-Ols with Radical Initiator. J. Wood Sci. 1999, 45, 53–63. [Google Scholar] [CrossRef]
- Son, K.-H.; Oh, M.-M. Growth, Photosynthetic and Antioxidant Parameters of Two Lettuce Cultivars as Affected by Red, Green, and Blue Light-Emitting Diodes. Hortic. Environ. Biotechnol. 2015, 56, 639–653. [Google Scholar] [CrossRef]
- Kapoor, S.; Raghuvanshi, R.; Bhardwaj, P.; Sood, H.; Saxena, S.; Chaurasia, O.P. Influence of Light Quality on Growth, Secondary Metabolites Production and Antioxidant Activity in Callus Culture of Rhodiola imbricata Edgew. J. Photochem. Photobiol. B Biol. 2018, 183, 258–265. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Shirazi, Z.; Aalami, A.; Tohidfar, M.; Sohani, M.M. Triterpenoid Gene Expression and Phytochemical Content in Iranian Licorice under Salinity Stress. Protoplasma 2019, 256, 827–837. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, W.; Sun, Y.; Shen, Y.; Mao, L.; Dai, Y.; Yang, B.; Liu, Z. Integrated Transcriptome and Metabolome Analysis Reveals Anthocyanin Biosynthesis Mechanisms in Pepper (Capsicum annuum L.) Leaves under Continuous Blue Light Irradiation. BMC Plant Biol. 2024, 24, 210. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, L.; Wang, Y.; Zhang, L.; Xu, S.; Wang, X.; He, W.; Zhang, Y.; Lin, Y.; Wang, Y. The Blue Light Signal Transduction Module FaCRY1-FaCOP1-FaHY5 Regulates Anthocyanin Accumulation in Cultivated Strawberry. Front. Plant Sci. 2023, 14, 1144273. [Google Scholar] [CrossRef]
- Kadomura-Ishikawa, Y.; Miyawaki, K.; Noji, S.; Takahashi, A. Phototropin 2 Is Involved in Blue Light-Induced Anthocyanin Accumulation in Fragaria × ananassa Fruits. J. Plant Res. 2013, 126, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chi, C.; Jin, L.; Zhu, J.; Yu, J.; Zhou, Y. The bZip Transcription Factor HY5 Mediates CRY1a-induced Anthocyanin Biosynthesis in Tomato. Plant Cell Environ. 2018, 41, 1762–1775. [Google Scholar] [CrossRef]
- Ojha, S.; Javed, H.; Azimullah, S.; Abul Khair, S.B.; Haque, M.E. Glycyrrhizic Acid Attenuates Neuroinflammation and Oxidative Stress in Rotenone Model of Parkinson’s Disease. Neurotox. Res. 2016, 29, 275–287. [Google Scholar] [CrossRef]
- Li, F.; Liu, B.; Li, T.; Wu, Q.; Xu, Z.; Gu, Y.; Li, W.; Wang, P.; Ma, T.; Lei, H. Review of Constituents and Biological Activities of Triterpene Saponins from Glycyrrhizae radix et Rhizoma and Its Solubilization Characteristics. Molecules 2020, 25, 3904. [Google Scholar] [CrossRef] [PubMed]
- Mutaillifu, P.; Bobakulov, K.; Abuduwaili, A.; Huojiaaihemaiti, H.; Nuerxiati, R.; Aisa, H.A.; Yili, A. Structural Characterization and Antioxidant Activities of a Water Soluble Polysaccharide Isolated from Glycyrrhiza glabra. Int. J. Biol. Macromol. 2020, 144, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.-K.; Wu, H.-T.; Ma, T.; Liu, W.-J.; He, X.-J. Effects of Glycyrrhiza glabra Polysaccharides on Immune and Antioxidant Activities in High-Fat Mice. Int. J. Biol. Macromol. 2009, 45, 61–64. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Strain | Explant | Hairy Roots Produced by Injection (%) | Hairy Roots Produced by Wounding and Soaking (%) | ||
|---|---|---|---|---|---|
| 10 min | 1 h | 24 h | |||
| 1724 | Cotyledon | 83.8 (±21.4) | 70.0 (±16.8) | 30.1 (±8.0) | 5.0 (±3.1) |
| Hypocotyl | 45.1 (±20.1) | 50.2 (±15.3) | 12.4 (±3.3) | 1.7 (±3.7) | |
| ATCC15834 | Cotyledon | 40.1 (±28.1) | 30.2 (±14.8) | 35.2 (±8.1) | 20.1 (±7.8) |
| Hypocotyl | 15.7 (±11.5) | 40.2 (±10.9) | 31.3 (±10.9) | 0.0 (±0.0) | |
| R1000 | Cotyledon | 10.2 (±9.5) | 3.3 (±4.7) | 10.3 (±3.4) | 0.0 (±0.0) |
| Hypocotyl | 12.1 (±11.5) | 5.1 (±3.7) | 4.7 (±2.7) | 0.0 (±0.0) | |
| A4 | Cotyledon | 26.2 (±14.0) | 25.0 (±11.2) | 4.9 (±2.8) | 2.5 (±5.6) |
| Hypocotyl | 0.0 (±0.0) | 10.3 (±7.4) | 0.0 (±0.0) | 10.3 (±3.3) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Afsharzadeh, N.; Paltram, R.; Jungwirth, A.; Tabrizi, L.; Nazeri, V.; Kalantari, H.; Halbwirth, H.; Samiei, L.; Sheehan, H.; Shokrpour, M. Advancing Glycyrrhiza glabra L. Cultivation and Hairy Root Transformation and Elicitation for Future Metabolite Overexpression. Horticulturae 2025, 11, 62. https://doi.org/10.3390/horticulturae11010062
Afsharzadeh N, Paltram R, Jungwirth A, Tabrizi L, Nazeri V, Kalantari H, Halbwirth H, Samiei L, Sheehan H, Shokrpour M. Advancing Glycyrrhiza glabra L. Cultivation and Hairy Root Transformation and Elicitation for Future Metabolite Overexpression. Horticulturae. 2025; 11(1):62. https://doi.org/10.3390/horticulturae11010062
Chicago/Turabian StyleAfsharzadeh, Negin, Renate Paltram, Arne Jungwirth, Leila Tabrizi, Vahideh Nazeri, Hadi Kalantari, Heidi Halbwirth, Leila Samiei, Hester Sheehan, and Majid Shokrpour. 2025. "Advancing Glycyrrhiza glabra L. Cultivation and Hairy Root Transformation and Elicitation for Future Metabolite Overexpression" Horticulturae 11, no. 1: 62. https://doi.org/10.3390/horticulturae11010062
APA StyleAfsharzadeh, N., Paltram, R., Jungwirth, A., Tabrizi, L., Nazeri, V., Kalantari, H., Halbwirth, H., Samiei, L., Sheehan, H., & Shokrpour, M. (2025). Advancing Glycyrrhiza glabra L. Cultivation and Hairy Root Transformation and Elicitation for Future Metabolite Overexpression. Horticulturae, 11(1), 62. https://doi.org/10.3390/horticulturae11010062