
Profiling Metabolites Distribution among Various Leaf Layers of Chinese Cabbage


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material Collection and Preparation
2.2. Soluble Sugar Assay
2.3. Carotenoids and Chlorophyll
2.4. Antioxidant and Antioxidant Capacity Assay
2.4.1. Vitamin C
2.4.2. Total Phenolic Compounds
2.4.3. Flavonoids
2.4.4. Total Antioxidant Capacity Assay
2.5. Elements Determination
2.6. Principal Component Analysis (PCA) and Statistical Analysis
3. Results
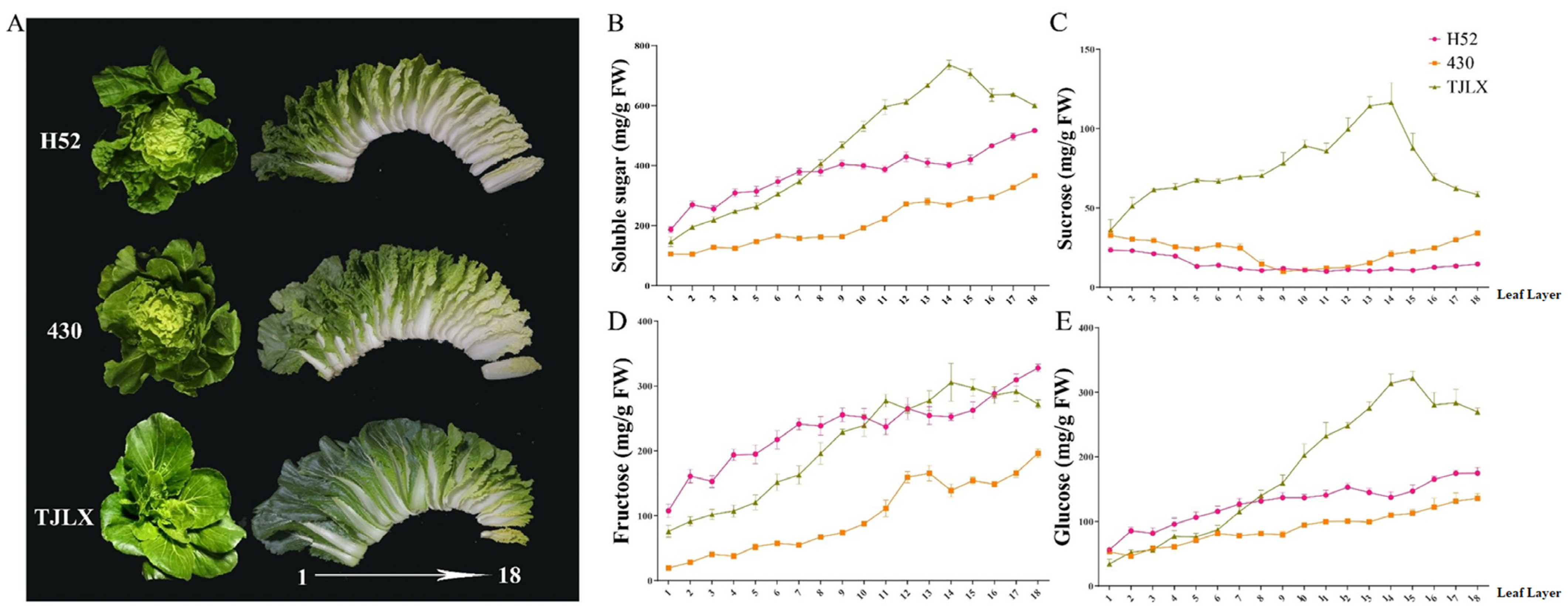
3.1. Soluble Sugar Content
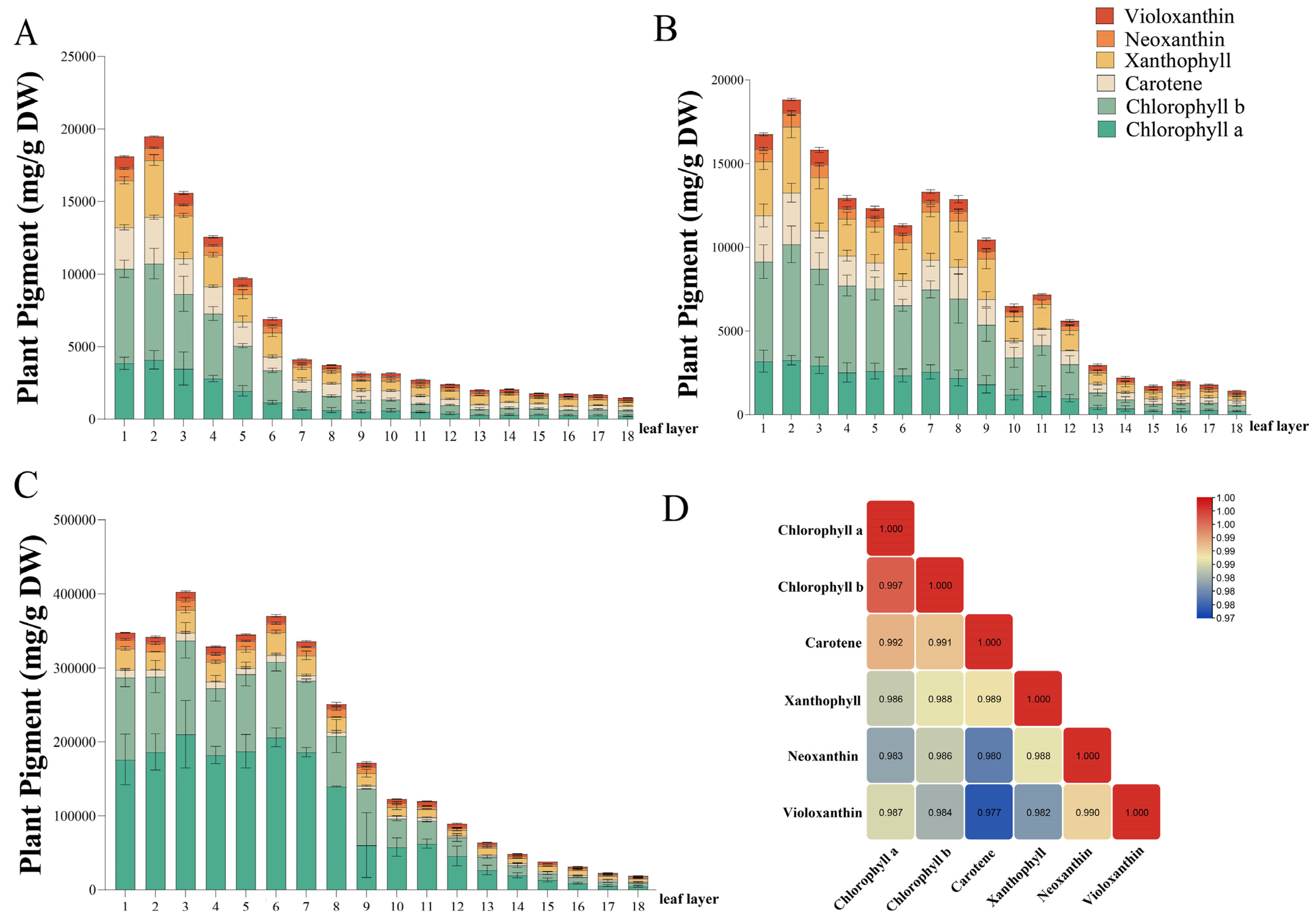
3.2. Vegetable Pigment
3.3. Vitamin C, Flavonoids, and Phenolic Contents
3.4. Glucosinolate Content
3.5. Mineral Content
3.6. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kris-Etherton, P.M.; Hecker, K.D.; Bonanome, A.; Coval, S.M.; Binkoski, A.E.; Hilpert, K.F.; Griel, A.E.; Etherton, T.D. Bioactive compounds in foods: Their role in the prevention of cardiovascular disease and cancer. Am. J. Med. 2002, 113, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Björkman, M.; Klingen, I.; Birch, A.; Bones, A.; Bruce, T.; Johansen, T.; Meadow, R.; Mølmann, J.; Seljåsen, R.; Smart, L.; et al. Phytochemicals of Brassicaceae in plant protection and human health--influences of climate, environment and agronomic practice. Phytochemistry 2011, 72, 538–556. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Giovannucci, E.; Hunter, D.; Neuberg, D.; Su, L.; Christiani, D. Dietary intake of Cruciferous vegetables, Glutathione S-transferase (GST) polymorphisms and lung cancer risk in a Caucasian population. Cancer Causes 2004, 15, 977–985. [Google Scholar] [CrossRef]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of α-Glucosidase and α-Amylase by Flavonoids. J. Nutr. Sci. Vitaminol. 2007, 52, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Wennberg, M.; Ekvall, J.; Olsson, K.; Nyman, M. Changes in carbohydrate and glucosinolate composition in white cabbage (Brassica oleracea var. capitata) during blanching and treatment with acetic acid. Food Chem. 2006, 95, 226–236. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef]
- Kusznierewicz, B.; Bartoszek, A.; Wolska, L.; Drzewiecki, J.; Gorinstein, S.; Namieśnik, J. Partial characterization of white cabbages (Brassica oleracea var. capitata f. alba) from different regions by glucosinolates, bioactive compounds, total antioxidant activities and proteins. Food Sci. Technol. 2008, 41, 1–9. [Google Scholar] [CrossRef]
- Seong, G.U.; Hwang, I.W.; Chung, S.-K. Antioxidant capacities and polyphenolics of Chinese cabbage (Brassica rapa L. ssp. Pekinensis) leaves. Food Chem. 2016, 199, 612–618. [Google Scholar] [CrossRef]
- Podsędek, A. Natural antioxidants and antioxidant capacity of Brassica vegetables: A review. Food Sci. Technol. 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, S.; Li, J.; Zhang, Y.; Zhou, D.; Li, C.; He, L.; Li, H.; Wang, F.; Gao, J. Identification of key genes controlling soluble sugar and glucosinolate biosynthesis in Chinese cabbage by integrating metabolome and genome-wide transcriptome analysis. Front. Plant Sci. 2022, 13, 1043489. [Google Scholar] [CrossRef]
- Heinze, M.; Hanschen, F.S.; Wiesner-Reinhold, M.; Baldermann, S.; Gräfe, J.; Schreiner, M.; Neugart, S. Effects of Developmental Stages and Reduced UVB and Low UV Conditions on Plant Secondary Metabolite Profiles in Pak Choi (Brassica rapa subsp. chinensis). J. Agric. Food Chem. 2018, 66, 1678–1692. [Google Scholar] [CrossRef] [PubMed]
- Harbaum, B.; Hubbermann, E.M.; Zhu, Z.; Schwarz, K. Free and bound phenolic compounds in leaves of pak choi (Brassica campestris L. ssp. chinensis var. communis) and Chinese leaf mustard (Brassica juncea Coss). Food Chem. 2008, 110, 838–846. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Wu, J.; Gao, W.; Wei, J.; Yang, J.; Guo, C. Antioxidant Capacity of Different Fractions of Vegetables and Correlation with the Contents of Ascorbic Acid, Phenolics, and Flavonoids. J. Food Sci. 2011, 76, C1257–C1261. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yue, Z.; Zhong, X.; Lei, J.; Tao, P.; Li, B. Distribution of primary and secondary metabolites among the leaf layers of headed cabbage (Brassica oleracea var. capitata)—ScienceDirect. Food Chem. 2020, 312, 126028. [Google Scholar] [CrossRef] [PubMed]
- Brantsæter, A.L.; Ydersbond, T.A.; Hoppin, J.A.; Haugen, M.; Meltzer, H.M. Organic food in the diet: Exposure and health implications. Annu. Rev. Public Health 2016, 38, 295–313. [Google Scholar] [CrossRef]
- Swegarden, H.; Stelick, A.; Dando, R.; Griffiths, P.D. Bridging sensory evaluation and consumer research for strategic leafy brassica (brassica oleracea) improvement. J. Food Sci. 2019, 84, 3746–3762. [Google Scholar] [CrossRef]
- Benmeziane, A.; Boulekbache-Makhlouf, L.; Mapelli-Brahm, P.; Khodja, N.K.; Remini, H.; Madani, K. Extraction of carotenoids from cantaloupe waste and determination of its mineral composition. Food Res. Int. 2018, 111, 391–398. [Google Scholar] [CrossRef]
- Liu, H.; Meng, F.; Miao, H.; Chen, S.; Yin, T.; Hu, S.; Shao, Z.; Liu, Y.; Gao, L.; Zhu, C. Effects of postharvest methyl jasmonate treatment on main health-promoting components and volatile organic compounds in cherry tomato fruits. Food Chem. 2018, 263, 194–200. [Google Scholar] [CrossRef]
- Fan, K.; Wang, M.; Gao, Y.; Ning, Q.; Shi, Y. Transcriptomic and ionomic analysis provides new insight into the beneficial effect of Al on tea roots’ growth and nutrient uptake. Plant Cell Rep. 2019, 38, 715–729. [Google Scholar] [CrossRef]
- Liu, Q.; Li, J.; Liu, W. Sugar accumulation and characterization of metabolizing enzyme genes in leafy head of Chinese cabbage (Brassica campestris L. ssp. pekinensis). Hortic. Environ. Biotechnol. 2020, 62, 17–29. [Google Scholar] [CrossRef]
- Shen, J.; Zou, Z.; Zhang, X.; Zhou, L.; Wang, Y.; Fang, W.; Zhu, X. Metabolic analyses reveal different mechanisms of leaf color change in two purple-leaf tea plant (Camellia sinensis L.) cultivars. Hortic. Res. 2018, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.; Mainson, D.; Porcheron, B.; Maurousset, L.; Lemoine, R.; Pourtau, N. Carbon source-sink relationship in Arabidopsis thaliana: The role of sucrose transporters. Planta Int. J. Plant Biol. 2018, 247, 587–611. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Qi, J.; Fang, Y.; Xiao, X.; Li, J.; Lan, J.; Tang, C. Characterization of Sugar Contents and Sucrose Metabolizing Enzymes in Developing Leaves of Hevea brasiliensis. Front. Plant Sci. 2018, 9, 58. [Google Scholar] [CrossRef]
- Rolland, F.; Baenagonzalez, E.; Sheen, J. Sugar sensing and signaling in plants: Conserved and Novel Mechanisms. Annu. Rev. 2006, 57, 675–709. [Google Scholar] [CrossRef]
- Young, A.J. The photoprotective role of carotenoids in higher plants. Physiol. Plant. 1991, 83, 702–708. [Google Scholar] [CrossRef]
- Baek, S.A.; Jung, Y.H.; Lim, S.H.; Park, S.U.; Kim, J.K. Metabolic Profiling in Chinese Cabbage (Brassica rapa L. subsp. pekinensis) Cultivars Reveals that Glucosinolate Content Is Correlated with Carotenoid Content. Agric. Food Chem. 2016, 64, 4426–4434. [Google Scholar] [CrossRef]
- Huxley, R.R.; Neil, H.A.W. The relation between dietary flavonol intake and coronary heart disease mortality: A meta-analysis of prospective cohort studies. Eur. J. Clin. Nutr. 2003, 57, 904–908. [Google Scholar] [CrossRef]
- Kim, E.H.; Ro, H.M.; Kim, S.L.; Kim, H.S.; Chung, I.M. Analysis of isoflavone, phenolic, soyasapogenol, and tocopherol compounds in soybean [Glycine max (L.) Merrill] germplasms of different seed weights and origins. J. Agric. Food Chem. 2012, 60, 6045–6055. [Google Scholar]
- Wei, J.; Miao, H.; Wang, Q. Effect of glucose on glucosinolates, antioxidants and metabolic enzymes in Brassica sprouts. Sci. Hortic. 2011, 129, 535–540. [Google Scholar] [CrossRef]
- Miao, H.; Wei, J.; Zhao, Y.; Yan, H.; Sun, B.; Huang, J.; Wang, Q. Glucose signalling positively regulates aliphatic glucosinolate biosynthesis. J. Exp. Bot. 2013, 64, 1097–1109. [Google Scholar] [CrossRef]
- Lee, M.K.; Chun, J.H.; Byeon, D.H.; Chung, S.O.; Park, S.U.; Park, S.; Arasu, M.V.; Al-Dhabi, N.A.; Lim, Y.P.; Kim, S.J. Variation of glucosinolates in 62 varieties of Chinese cabbage (Brassica rapa L. ssp. pekinensis) and their antioxidant activity. Food Sci. Technol. 2014, 58, 93–101. [Google Scholar] [CrossRef]
- Flyman, M.V.; Afolayan, A.J. The implication of the mineral ratios of Cucumis myriocarpus Naud. and Pergularia daemia (Forsk.) Chiov. in human diets. J. Med. Food 2007, 10, 548–551. [Google Scholar] [CrossRef] [PubMed]
- Mobeen; Wang, X.; Saleem, M.H.; Parveen, A.; Mumtaz, S.; Hassan, A.; Adnan, M.; Fiaz, S.; Ali, S.; Iqbal Khan, Z.; et al. Proximate Composition and Nutritive Value of Some Leafy Vegetables from Faisalabad, Pakistan. Sustainability 2021, 13, 8444. [Google Scholar] [CrossRef]
- Jie, W.; Zeci, L.; Jianhua, D.; Jian, L.; Ning, J.; Li, J.; Zhaozhuang, L.; Bo, Z.; Zhongqi, T.; Jihua, Y. A Comparative Study on the Nutrients, Mineral Elements, and Antioxidant Compounds in Different Types of Cruciferous Vegetables. Agronomy 2022, 12, 3121. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Layer | H52 | 430 | TJLX | ||||||
|---|---|---|---|---|---|---|---|---|---|
| AGS | IGS | GS | AGS | IGS | GS | AGS | IGS | GS | |
| 1 | 0.474 ± 0.121 a | 0.038 ± 0.005 ab | 0.512 ± 0.119 a | 0.207 ± 0.004 a | 0.157 ± 0.055 a | 0.364 ± 0.052 ab | 0.190 ± 0.088 cd | 0.235 ± 0.085 a | 0.424 ± 0.010 a |
| 2 | 0.333 ± 0.028 bc | 0.054 ± 0.032 a | 0.387 ± 0.007 bcde | 0.105 ± 0.041 a | 0.167 ± 0.044 a | 0.272 ± 0.085 a | 0.158 ± 0.026 cde | 0.318 ± 0.161 a | 0.476 ± 0.185 a |
| 3 | 0.369 ± 0.053 abc | 0.025 ± 0.004 bc | 0.394 ± 0.051 abcd | 0.103 ± 0.032 a | 0.078 ± 0.019 a | 0.181 ± 0.046 a | 0.078 ± 0.044 defg | 0.333 ± 0.015 a | 0.411 ± 0.037 a |
| 4 | 0.396 ± 0.035 abc | 0.020 ± 0.001 bc | 0.417 ± 0.035 cde | 0.087 ± 0.009 a | 0.160 ± 0.029 a | 0.248 ± 0.006 a | 0.098 ± 0.011 defg | 0.504 ± 0.038 a | 0.602 ± 0.038 a |
| 5 | 0.346 ± 0.008 bc | 0.008 ± 0.005 cd | 0.353 ± 0.003 cde | 0.097 ± 0.021 a | 0.108 ± 0.027 a | 0.205 ± 0.006 a | 0.369 ± 0.043 a | 0.267 ± 0.026 a | 0.635 ± 0.068 a |
| 6 | 0.357 ± 0.078 bc | 0.000 ± 0.000 d | 0.357 ± 0.078 cde | 0.109 ± 0.009 a | 0.158 ± 0.018 a | 0.267 ± 0.024 a | 0.126 ± 0.040 cdef | 0.528 ± 0.145 a | 0.654 ± 0.176 a |
| 7 | 0.338 ± 0.014 bc | 0.000 ± 0.000 d | 0.338 ± 0.014 cde | 0.092 ± 0.008 a | 0.135 ± 0.011 a | 0.227 ± 0.017 a | 0.226 ± 0.158 bc | 0.497 ± 0.071 a | 0.723 ± 0.160 a |
| 8 | 0.315 ± 0.039 bc | 0.000 ± 0.000 d | 0.315 ± 0.039 cde | 0.130 ± 0.023 a | 0.163 ± 0.049 a | 0.294 ± 0.069 a | 0.082 ± 0.013 defg | 0.505 ± 0.062 a | 0.587 ± 0.070 a |
| 9 | 0.373 ± 0.022 abc | 0.000 ± 0.000 d | 0.373 ± 0.022 bcde | 0.110 ± 0.017 a | 0.224 ± 0.041 a | 0.333 ± 0.040 ab | 0.041 ± 0.011 efg | 0.726 ± 0.049 a | 0.766 ± 0.051 a |
| 10 | 0.424 ± 0.053 ab | 0.000 ± 0.000 d | 0.424 ± 0.053 abcd | 0.112 ± 0.001 a | 0.243 ± 0.170 a | 0.355 ± 0.171 ab | 0.011 ± 0.003 fg | 0.698 ± 0.093 a | 0.709 ± 0.094 a |
| 11 | 0.293 ± 0.045 c | 0.000 ± 0.000 d | 0.293 ± 0.045 e | 0.218 ± 0.131 a | 0.545 ± 0.350 a | 0.763 ± 0.481 abc | 0.321 ± 0.025 ab | 0.311 ± 0.041 a | 0.631 ± 0.055 a |
| 12 | 0.365 ± 0.024 abc | 0.000 ± 0.000 d | 0.365 ± 0.024 bcde | 0.111 ± 0.037 a | 0.252 ± 0.120 a | 0.362 ± 0.155 ab | 0.013 ± 0.006 fg | 0.645 ± 0.057 a | 0.658 ± 0.052 a |
| 13 | 0.320 ± 0.019 bc | 0.000 ± 0.000 d | 0.320 ± 0.019 cde | 0.161 ± 0.034 a | 0.338 ± 0.278 a | 0.499 ± 0.300 ab | 0.000 ± 0.000 g | 0.659 ± 0.089 a | 0.759 ± 0.089 a |
| 14 | 0.336 ± 0.019 bc | 0.000 ± 0.000 d | 0.336 ± 0.019 cde | 0.180 ± 0.043 a | 0.265 ± 0.059 a | 0.446 ± 0.100 ab | 0.000 ± 0.000 g | 0.695 ± 0.020 a | 0.695 ± 0.020 a |
| 15 | 0.313 ± 0.013 bc | 0.000 ± 0.000 d | 0.313 ± 0.013 de | 0.125 ± 0.018 a | 0.107 ± 0.032 a | 0.232 ± 0.040 a | 0.012 ± 0.001 fg | 1.782 ± 0.080 b | 1.793 ± 0.081 b |
| 16 | 0.405 ± 0.031 abc | 0.000 ± 0.000 d | 0.405 ± 0.031 abcd | 0.755 ± 0.152 b | 0.412 ± 0.573 a | 1.167 ± 0.435 c | 0.008 ± 0.006 fg | 1.687 ± 0.024 b | 1.696 ± 0.030 b |
| 17 | 0.472 ± 0.039 a | 0.000 ± 0.000 d | 0.472 ± 0.039 ab | 0.874 ± 0.533 b | 0.009 ± 0.005 a | 0.882 ± 0.538 bc | 0.046 ± 0.004 efg | 2.558 ± 0.294 c | 2.604 ± 0.290 c |
| 18 | 0.412 ± 0.016 ab | 0.000 ± 0.000 d | 0.412 ± 0.016 abcd | 0.741 ± 0.292 b | 0.009 ± 0.004 a | 0.750 ± 0.296 abc | 0.086 ± 0.027 defg | 3.700 ± 1.028 d | 3.786 ± 1.054 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Wang, H.; Mei, Y.; Yue, Z.; Lei, J.; Tao, P.; Li, B.; Zhao, J.; Hu, Q. Profiling Metabolites Distribution among Various Leaf Layers of Chinese Cabbage. Horticulturae 2024, 10, 988. https://doi.org/10.3390/horticulturae10090988
Zhao Y, Wang H, Mei Y, Yue Z, Lei J, Tao P, Li B, Zhao J, Hu Q. Profiling Metabolites Distribution among Various Leaf Layers of Chinese Cabbage. Horticulturae. 2024; 10(9):988. https://doi.org/10.3390/horticulturae10090988
Chicago/Turabian StyleZhao, Yanting, Huasen Wang, Ying Mei, Zhichen Yue, Juanli Lei, Peng Tao, Biyuan Li, Jianjun Zhao, and Qizan Hu. 2024. "Profiling Metabolites Distribution among Various Leaf Layers of Chinese Cabbage" Horticulturae 10, no. 9: 988. https://doi.org/10.3390/horticulturae10090988
APA StyleZhao, Y., Wang, H., Mei, Y., Yue, Z., Lei, J., Tao, P., Li, B., Zhao, J., & Hu, Q. (2024). Profiling Metabolites Distribution among Various Leaf Layers of Chinese Cabbage. Horticulturae, 10(9), 988. https://doi.org/10.3390/horticulturae10090988