Study on the Identification, Biological Characteristics, and Fungicide Sensitivity of the Causal Agent of Strawberry Red Core Root Rot

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Sample Collection
2.2. Isolation, Purification, and Morphological Observations
2.3. Pathogenicity Test
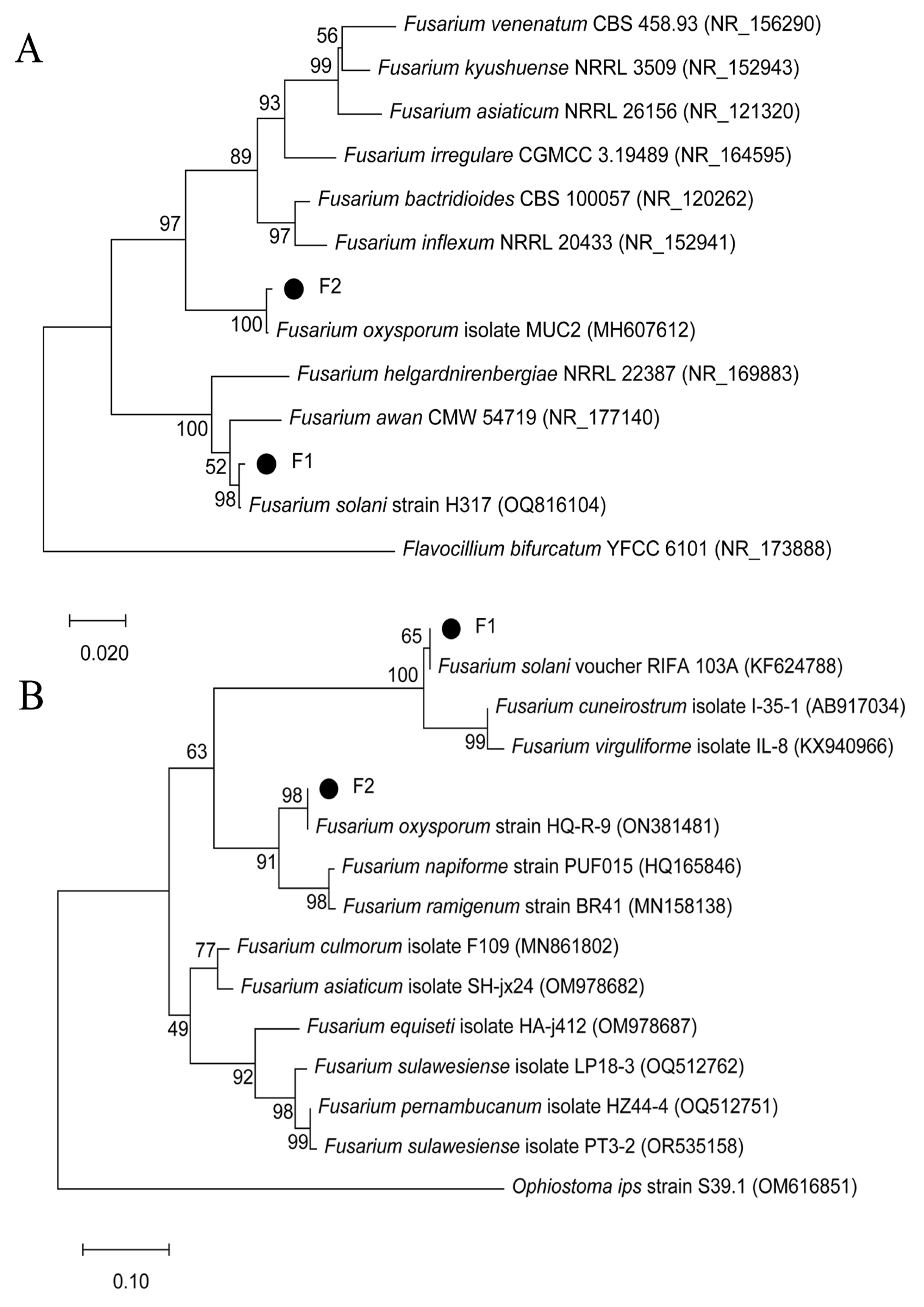
2.4. Molecular Identification of the Pathogen
2.5. Morphological Characteristics
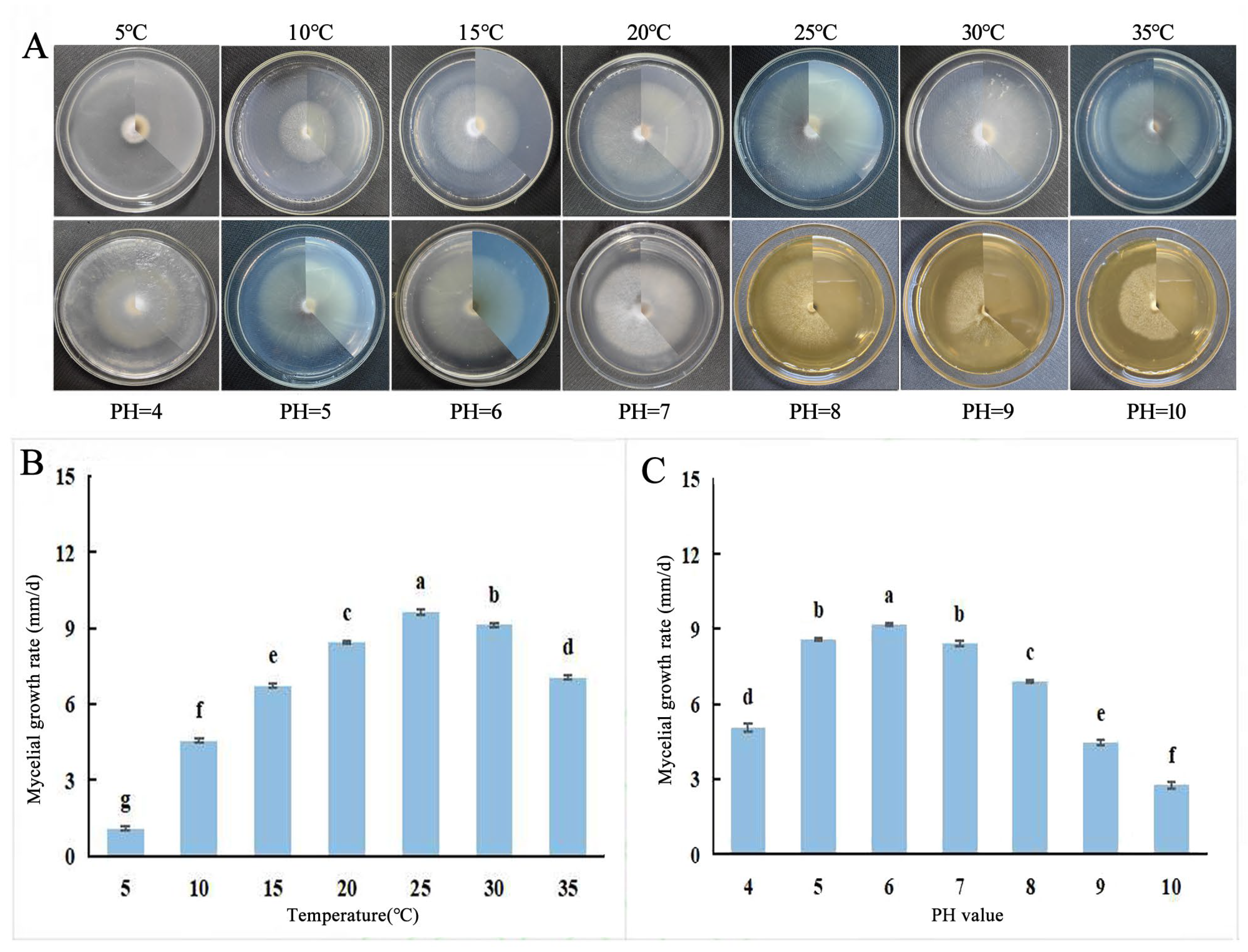
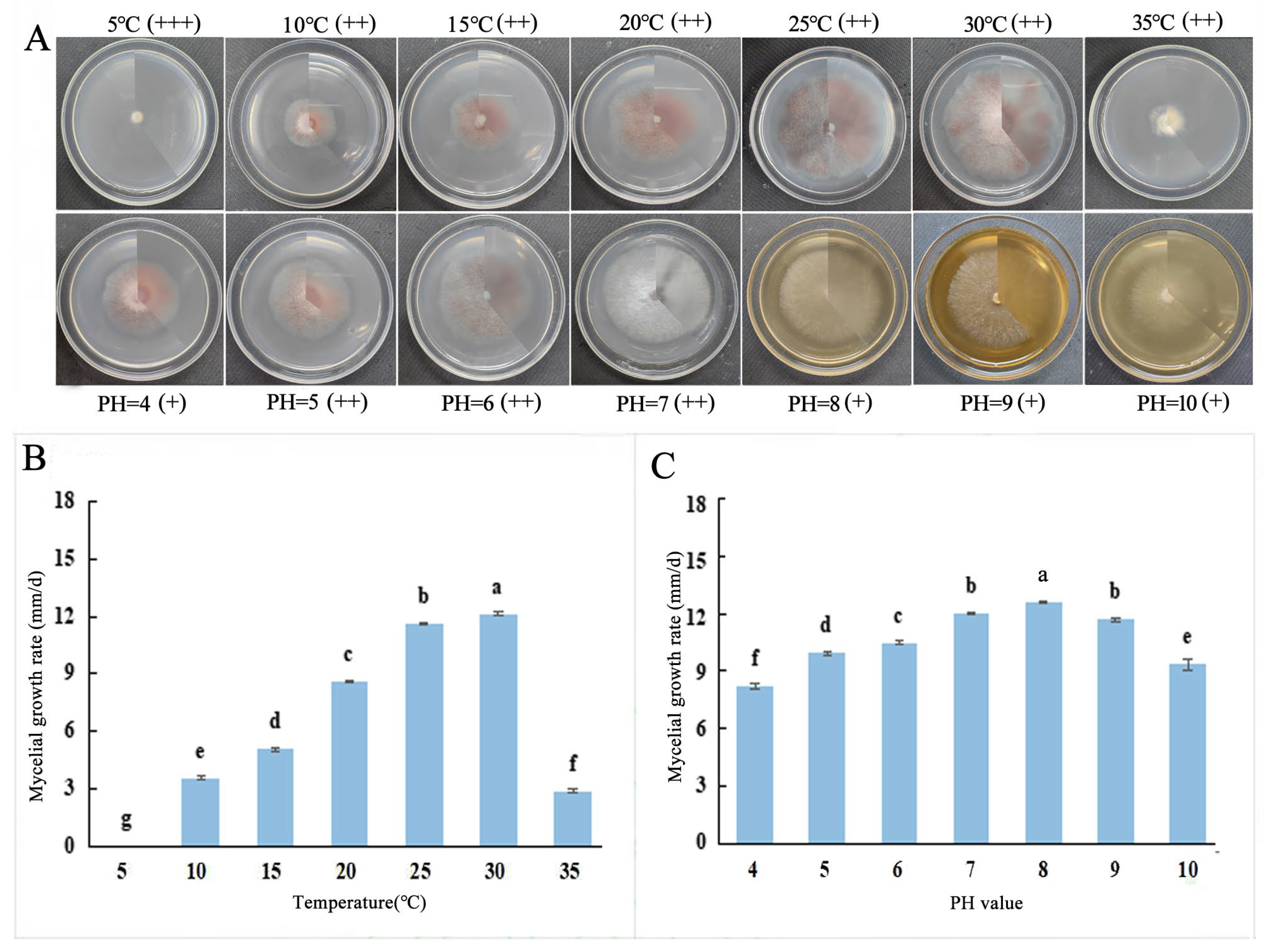
2.5.1. Effects of Different Temperatures, pH Values, and Lighting Conditions on the Growth of Fungal Hyphae
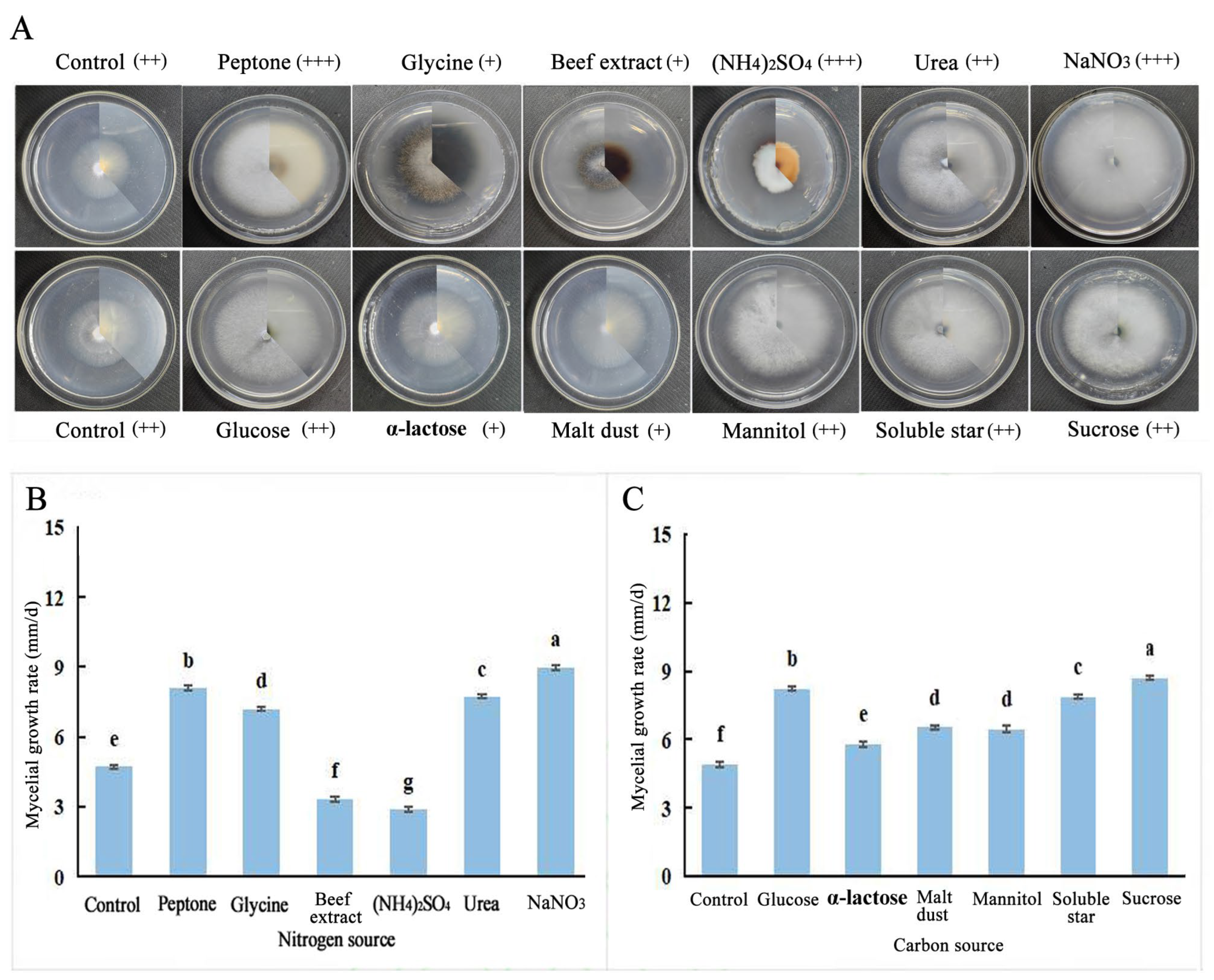
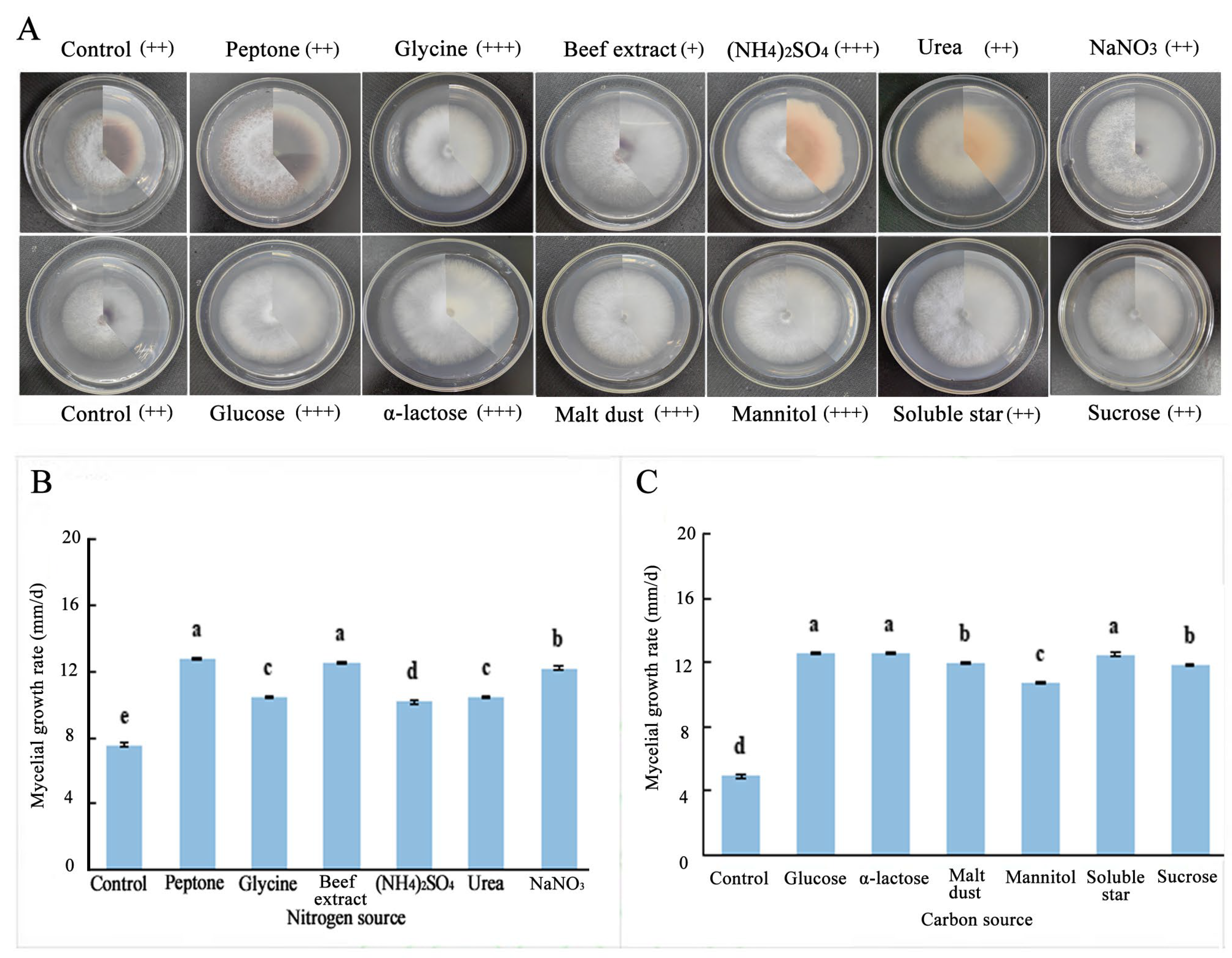
2.5.2. Effects of Different Carbon and Nitrogen Sources on the Growth of Fungal Hyphae
2.6. Evaluation of the Susceptibility of the Pathogens to Five Fungicides
2.7. Statistical Analysis
3. Results and Discussion
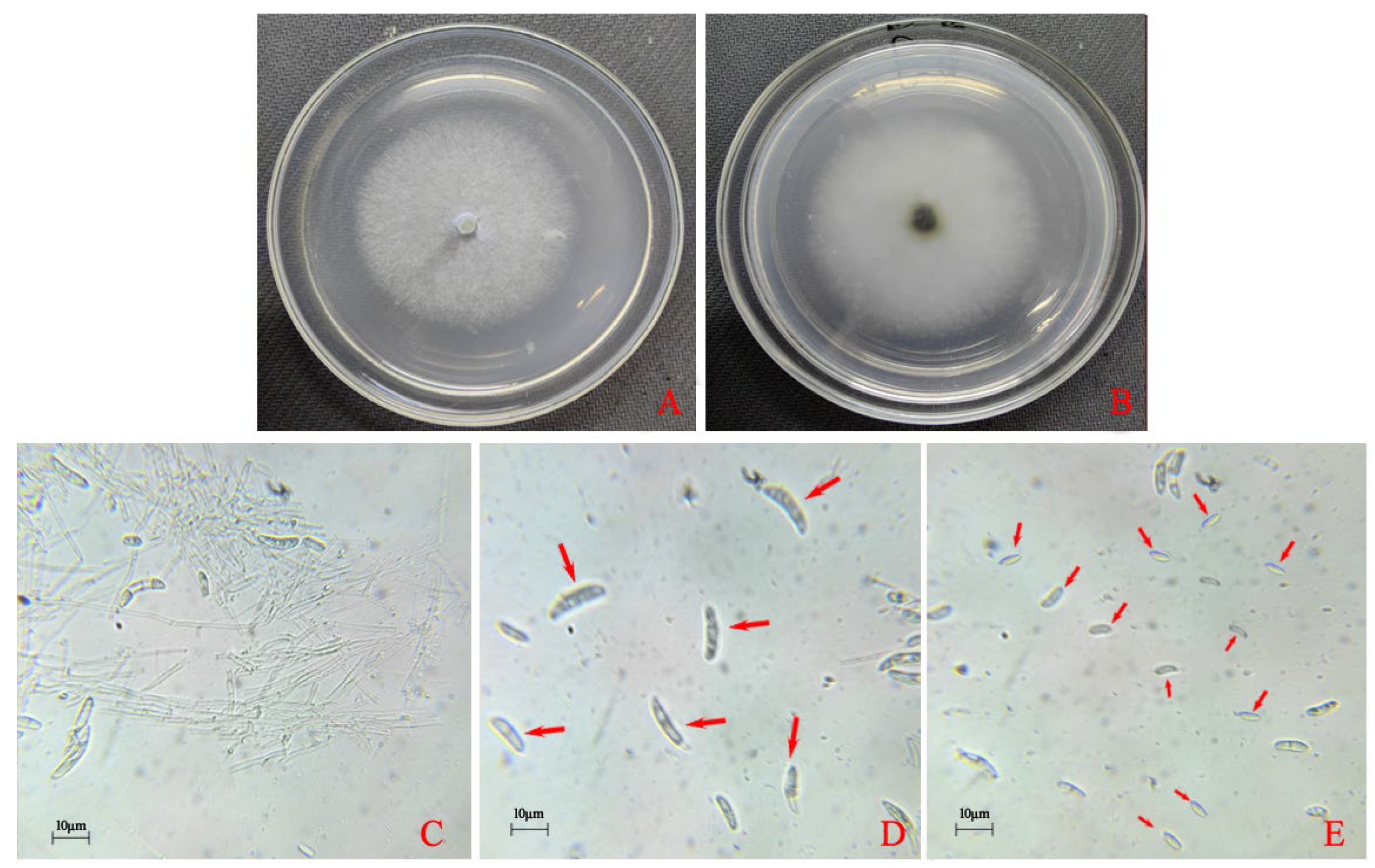
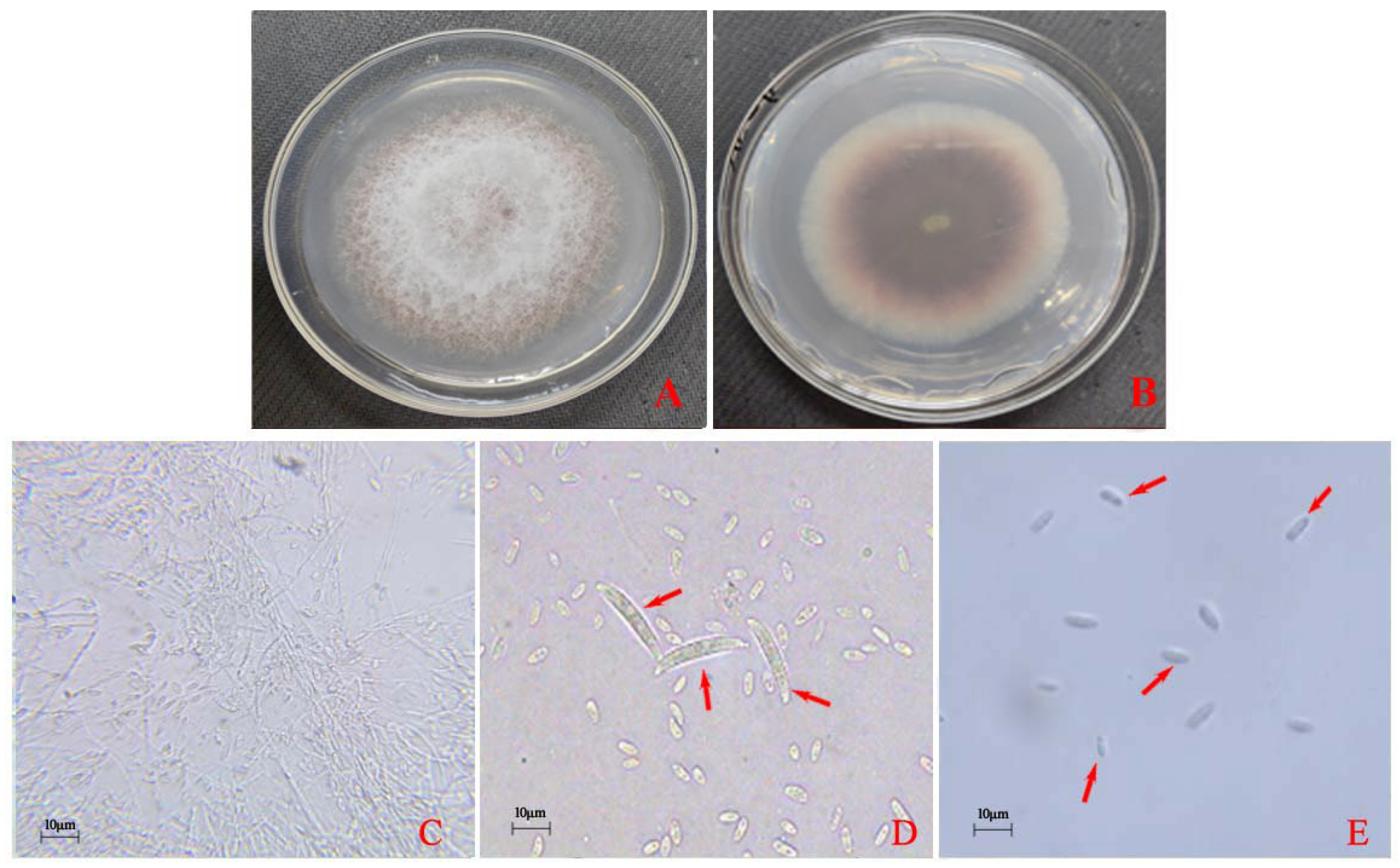
3.1. Morphological and Molecular Identification of the Pathogens
3.2. Incidence and Symptoms of F. solani and F. oxysporum
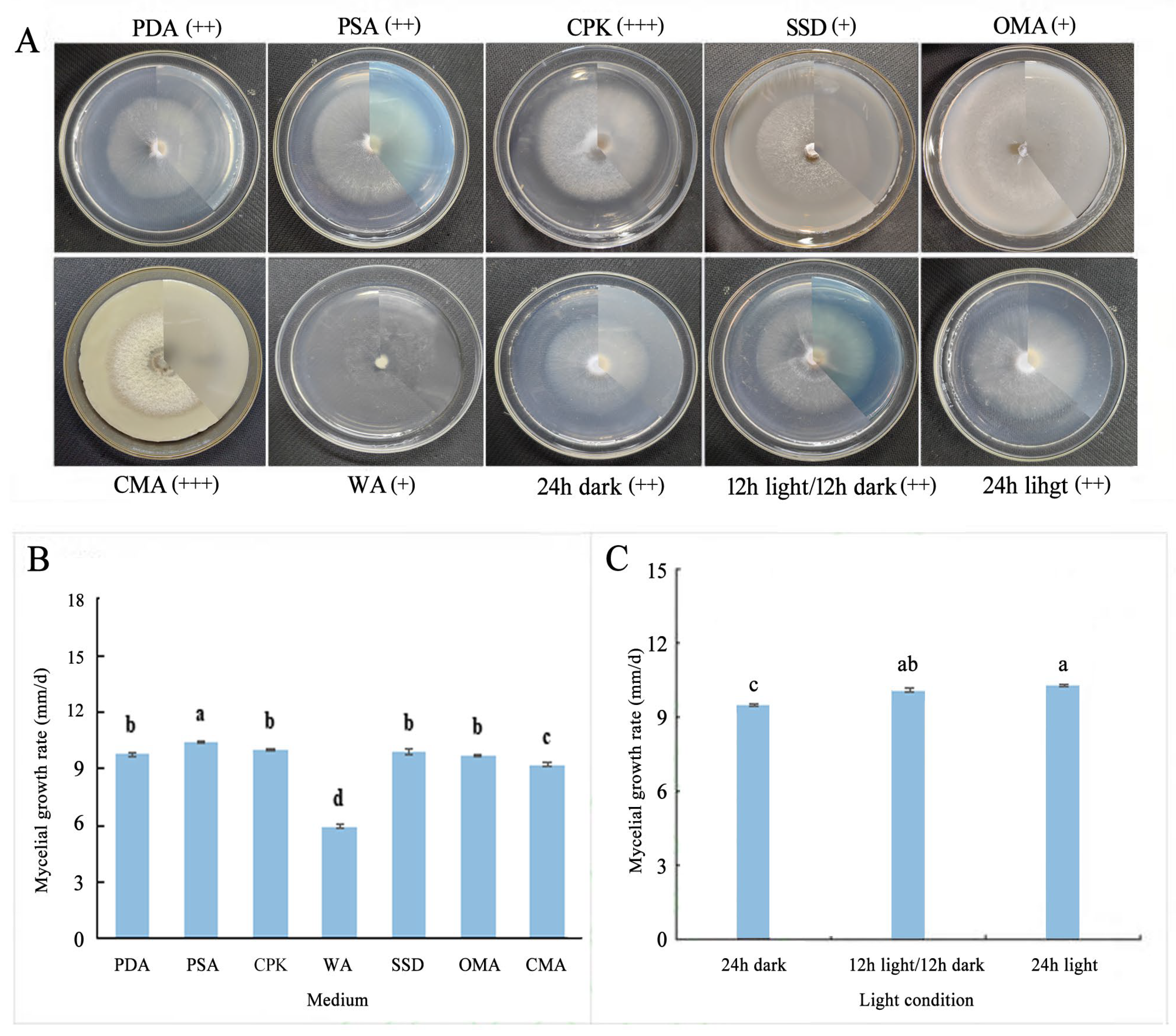
3.3. Biological Characteristics of F. solani and F. oxysporum
3.4. Effects of Five Different Fungicides on the Growth of the Pathogen
3.5. Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Maas, J.L. Compendium of Strawberry Diseases; China Agricultural Press: Beijing, China, 2012. [Google Scholar]
- Lei, J.J.; Dai, H.P.; Tan, C.H.; Deng, M.Q.; Zhao, M.Z. Studies on the taxonomy of the strawberry (Fragaria) species distributed in China. Acta Hortic. Sin. 2006, 33, 1–5. [Google Scholar]
- Giampieri, F.; Alvarez-Suarez, J.M.; Battino, M. Strawberry and human health: Effects beyond antioxidant activity. J. Agric. Food Chem. 2014, 62, 3867–3876. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.H.; Zhang, L.; Peng, F.T. Effect of sodium selenite on the growth and development of strawberries under salt stress. China Fruits 2023, 10, 52–57. [Google Scholar]
- Peng, M.L.; He, Y.L.; Yao, B.; Wei, F.J.; Zhang, J.H.; Qiu, Y.; Ji, G.H. Isolation, identification and resistance evaluation of strawberry angular leaf spot pathogen in Yunnan Province. Acat Phytpaththologica 2023, 10, 1–13. [Google Scholar]
- Su, D.F.; Dai, Q.Z.; Yan, C.W.; Zhang, Z.R.; Dong, Q.E.; Tian, Y.X.; Zhou, W.X.; Chen, S.Y.; Yang, J.Y.; Tong, J.Y.; et al. Research progress on strawberry root rot and its biological control. Jiangsu Agric. Sci. 2022, 50, 16–26. [Google Scholar]
- Wang, F.; Zhang, S.Y.; Lu, J.; Ma, Y. Research Developments on Strawberry Anthracnose. J. Shenyang Agric. Univ. 2023, 54, 490–501. [Google Scholar]
- Gao, H.L.; Yuan, L.; Yao, W.L. Difficulties in the prevention and control of strawberry gray mold in greenhouses and green prevention and control measures. Mod. Agric. Sci. Technol. 2020, 12, 140–141. [Google Scholar]
- Shen, G.H. Studies on Mechanism of Strawberry Replant Root Rot Disease and Alleviation with Biological and Cemmical Methods; Northwest A&F University: Xianyang, China, 2012. [Google Scholar]
- Hickman, C.J. The red core root disease of the strawberry caused by Phytophthora fragariae n. sp. J. Pomol. Hortic. Sci. 1940, 18, 89–118. [Google Scholar]
- Morita, H. Red stele root disease of strawberry caused by Phytophthora Fragariae. Jpn. J. Phytopathol. 1965, 30, 239–245. [Google Scholar] [CrossRef]
- Milholland, R.D.; Daykin, M.E. Colonization of roots of strawberry cultivars with different levels of susceptibility to Phytophthora fragariae. Phytopathology 1993, 83, 538–542. [Google Scholar] [CrossRef]
- Gaetán, S.; Madia, M.; Cepeda, R. First report of Fusarium crown and root rot caused by Fusarium solani on St. John’s-wort in Argentina. Plant Dis. 2004, 88, 1050. [Google Scholar] [CrossRef] [PubMed]
- Mu, L.S.; Liu, Z.H.; Sun, J. Study on the Biological Characteristics of Strawberry Red Pillar Root Rot Pathogen. North. Hortic. 2008, 3, 219–222. [Google Scholar]
- Ren, J.J.; Liu, Z.Y.; Wang, Y.; Fong, J.T.; He, J. Indoor screening of biofungicides for strawberry root rot and verification of field control efficacy. Chin. J. Pestic. Sci. 2022, 24, 1456–1465. [Google Scholar]
- Chen, X.X.; Zheng, Y.J.; Wang, Z.Y. Identification and Biological Characteristics of Pathogenic Fungi of Strawberry Root Rot in Dandong. Liaoning Agric. Sci. 2023, 6, 75–78. [Google Scholar]
- Sheng, R.Y.; Xiao, C.K.; Zheng, S.H. Identification of Strawberry Root Rot Pathogeny Caused by Fusarium in Beijing. China Veg. 2012, 12, 52–56. [Google Scholar]
- Wang, Y.M.; Chen, Q.Q.; Zhu, H.M. Pathogenicity and Fungicides Virulence Assessment of the Strawberry Red Pillar Rot Pathogen. J. Anhui Sci. Technol. Univ. 2014, 28, 15–18. [Google Scholar]
- Lavtižar, V.; Helmus, R.; Kools, S.A.E.; Dolenc, D.; van Gestel, C.A.M.; Trebše, P.; Waaijers, S.L.; Kraak, M.H.S. Daphnid life cycle responses to the insecticide chlorantraniliprole and its transformation products. Environ. Sci. Technol. 2015, 49, 3922–3929. [Google Scholar] [CrossRef]
- Dong, H.F.; Li, C.Y.; Xu, Y.B.; Zhang, X.Y.; Zhang, X.D.; Liu, Y.B. Field Efficacy Test of Different Pesticides Against Strawberry Red Stele Root Rot. Mod. Agric. Sci. Technol. 2019, 18, 73–74+78. [Google Scholar]
- Wang, Y.C.; Zheng, X.B.; Hua, Y.G.; Lu, J.Y. Inheritance of the resistance of Phytophthora spp. to cymoxanil and its cross resistance to cymoxanil and metalaxyl. Plant Prot. 1997, 23, 7–10. [Google Scholar]
- Ma, D.C. Resistance of Phytophthora Capsici to Azoxystrobin and Sensitivity to Tetramycin and Benzothiostrobin; Shandong Agricultural University: Taian, China, 2019. [Google Scholar]
- Liu, S.T. Monitoring and Carbendzim Mechanisms of Resistance to Difenoconazole and Carbendzim by Didymella Bryoniae in the Region of Chongqing and Guizhou; Southwest University: El Paso, TX, USA, 2016. [Google Scholar]
- Wei, C.; Guo, L.A.; Hu, L.; Dai, X.H.; Mao, J.F. Pesticide residues detection of strawberries surface and its effects on microflora. Southwest China J. Agric. Sci. 2014, 27, 2534–2538. [Google Scholar]
- Cao, S.; Yang, N.B.; Zhao, C. Diversity of Fusarium species associated with root rot of sugar beet in China. J. Gen. Plant Pathol. 2018, 84, 321–329. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Ames, IA, USA, 2006. [Google Scholar]
- Zhang, X.L.; Cao, Z.Y. Identification of the pathogen causing root rot of Dendrobium officinale and sensitivity to the fermentation broth of Streptomyces. Chin. J. Biol. Control 2022, 38, 258–266. [Google Scholar]
- An, X.L.; Wu, X.; Ren, Y.F.; Yin, Q.X.; Wang, X.; Li, D.X.; Chen, Z. Identification and biological characteristics of Pseu dopestalotiopsis theae, an important pathogenic fungus causing tealeaf spot. China Plant Prot. 2020, 40, 12–19. [Google Scholar]
- Xi, Z.G.; You, J.M.; Zheng, L.; Liu, H.; Huang, J.B. Identification and biological characteristics of the pathogen causing leaf spot on Coptis chinensis in hubei province. J. Plant Prot. 2018, 45, 315–321. [Google Scholar]
- Wen, J.L.; Ming, H.Y.; Xue, Z.J.; Li, Y.F.; Zhang, D.X.; Zheng, G.T.; Lu, J.X.; Wang, J.N.; Zhou, J.N. Five fungal pathogens are responsible for bayberry twig blight and fungicides were screened for disease control. Microorganisms 2020, 8, 689. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- O’donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, J.; Shi, H.; Liang, C.H.; Wang, Z.G. Species identification of Fusarium spp. causing root rot on strawberry. Acta Phytopathol. Sin. 2024, 54, 451–456. [Google Scholar]
- Wang, B.B.; Zhang, N.; Gong, P.T.; Li, J.H.; Ju, Y.; Zhang, H.B.; Zhang, X.C.; Cai, K.Z. Morphological variability, molecular phylogeny, and biological characteristics of the nematophagous fungus Duddingtonia flagrans. J. Basic Microbiol. 2019, 59, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Luo, N.; Zhang, F. Identification and biological characteristics of the pathogen of Alternaria tenuifolia leaf spot of raspberry. North. Hortic. 2021, 16, 24–32. [Google Scholar]
- Mu, L. Research Methods of Chemical Plant Protection; China Agriculture Press: Beijing, China, 1994. [Google Scholar]
- Wen, H.; Wei, J.S.; Zhang, G.J.; Bi, Y.; Yan, Z. Laboratory toxicity of nine fungicides against Neopestalotiopsis clavispora. Chin. J. Pestic. Sci. 2019, 21, 437–443. [Google Scholar]
- Wang, Q.H.; Fan, K.; Li, D.W.; Han, C.M.; Qu, Y.Y.; Qi, Y.K.; Wu, X.Q. Identification, Virulence and Fungicide Sensitivity of Colletotrichum gloeosporioides s.s. Responsible for Walnut Anthracnose Disease in China. Plant Dis. 2019, 104, 1358–1368. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.Y.; Shang, J.Y.; Gao, C.; Su, Z.H.; Wang, Y.; Zhang, X.Y.; Guo, Q.G.; Li, S.Z.; Ma, P. Identification, biological characteristics and control agents of the pathogen causing belladonna basal stem rot. Acta Phytopathol. Sin. 2023, 1–15. [Google Scholar] [CrossRef]
- Eikemo, H.; Brurberg, M.B.; Davik, J. Resistance to Phytophthora cactorum in diploid Fragaria species. HortScience 2010, 45, 193–197. [Google Scholar] [CrossRef]
- Chen, X.R.; Wang, Z.R.; Xu, J.Y.; Dong, Y.H. Construction and analysis of SSH of strawberry crown upon infection with Phytophthora cactorum. Yangzhou Univ. Agric. Life Sci. Ed. 2011, 32, 64–69. [Google Scholar]
- Chen, Z.; Huang, J.; Zhao, J.; Liang, H. Research Advance on the Red Stele Root Rot of Strawberry. Biotechnol. Bull. 2017, 33, 37–44. [Google Scholar]
- Cao, K.R. Studies on Pathogen Identification and Biological Characteristics of Strawberry Root Rot; Gansu Agricultural University: Lanzhou, China, 2006. [Google Scholar]
- Chen, W.L. Biological Characteristics of Gerbera Root Rot Pathogen and Field Screening of Efficient Fungicides in Sanming of Fujian Province. Chin. Agric. Sci. Bull. 2023, 39, 107–111. [Google Scholar]
- Qu, J.; Wu, H.B.; Fan, K.; Sun, Y.G.; Li, J.; Tao, J.H. Biologicalcharacteristics of Collelotrichum gloeosporioides and inhibitory effects of several fungicide. Chin. J. Pestic. Sci. 2012, 14, 503–509. [Google Scholar]
- Gao, Y.Y.; He, L.F.; Li, B.X.; Lin, J.; Mu, W.; Liu, F. Identification of the pathogen causing pepper anthracnose in shandong province and screening of highly effective fungicides. Sci. Agric. Sin. 2017, 50, 1452–1464. [Google Scholar]
- Xue, J.; Tian, Z.; Liu, D. Occurrence rule and control test of persimmon anthracno. Shanxi Fruits 2003, 4, 5–7. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Media | Content |
|---|---|
| Potato dextrose agar (PDA) | 200 g peeled potato, 20 g agar, 20 g glucose, 1000 mL sterile water |
| Potato sucrose agar (PSA) | 200 g peeled potato, 20 g agar, 20 g sucrose, 1000 mL sterile water |
| Corn meal agar (CMA) | 40 g corn flour, 20 g agar, 1000 mL sterile water |
| Czapek agar (CPK) | 20 g agar, 2 g sodium nitrate, 1 g dipotassium hydrogen phosphate, 0.5 g potassium chloride, 0.5 g magnesium sulfate heptahydrate, 0.01 g ferrous sulfate heptahydrate, 20 g sucrose, 1000 mL sterile water |
| Oatmeal agar (OMA) | 30 g oatmeal, 20 g agar, 1000 mL sterile water |
| Strawberry stem decoction (SSD) | 200 g fresh strawberry stem stock, 20 g agar, 20 g glucose, 1000 mL sterile water |
| Water agar (WA) | 20 g agar, 1000 mL sterile water |
| Fungicide (Purity %) | Manufacturer |
|---|---|
| Tebuconazole (97.5%) | Jiangsu Fengdeng Pesticide Co., Ltd., Changzhou, China |
| Azoxystrobin (98.5%) | Jiangyin Suli Chemical Co., Ltd., Jiangyin, China |
| Chlorothalonil (98.0%) | Shandong Huayang Pesticide Fertilizer Chemical Co., Ltd., Taian, China |
| Prochloraz (97.0%) | Hubei Shenglong Chemical Co., Ltd., Wuhan, China |
| Carbendazim (98.0%) | Shanghai Linkong Chemical Trade Co., Ltd., Shanghai, China |
| Fungicide | Concentrations (μg/mL) | Colonidia Meter (cm) | Inhibition Rates (%) | Toxicity Regression Equations | R Correlation Coefficient | EC50 (μg/mL) | 95% CL (μg/mL) |
|---|---|---|---|---|---|---|---|
| Prochloraz (95%) | 0.004 | 6.58 ± 0.14 b | 6.27 ± 0.12 g | y = 1.232x + 1.297 | 0.996 | 0.088 ± 0.002 d | 0.072–0.109 |
| 0.010 | 6.28 ± 0.08 c | 10.54 ± 0.99 f | |||||
| 0.025 | 5.49 ± 0.09 d | 21.79 ± 2.40 e | |||||
| 0.063 | 3.88 ± 0.08 e | 44.73 ± 0.86 d | |||||
| 0.158 | 2.42 ± 0.11 f | 65.53 ± 5.28 c | |||||
| 0.395 | 1.53 ± 0.13 g | 78.21 ± 1.76 b | |||||
| 0.988 | 0.76 ± 0.18 h | 89.17 ± 1.96 a | |||||
| Control | 7.02 ± 0.11 a | — | |||||
| Carbendazim (95%) | 0.50 | 6.49 ± 0.09 b | 7.55 ± 0.15 g | y = 1.564x + 0.046 | 0.856 | 0.935 ± 0.026 d | 0.503–2.753 |
| 0.10 | 6.15 ± 0.10 c | 12.39 ± 1.23 f | |||||
| 0.20 | 5.53 ± 0.16 d | 21.23 ± 0.67 e | |||||
| 0.40 | 4.89 ± 0.11 e | 30.34 ± 1.83 d | |||||
| 0.80 | 4.06 ± 0.15 f | 42.17 ± 1.17 c | |||||
| 1.60 | 2.29 ± 0.20 g | 67.38 ± 1.20 b | |||||
| 3.20 | 0.80 ± 0.11 h | 88.60 ± 2.99 a | |||||
| Control | 7.02 ± 0.11 a | — | |||||
| Azoxystrobin (98.5%) | 25 | 6.93 ± 0.08 a | 1.28 ± 0.23 g | y = 1.626x − 4.405 | 0.998 | 511.531 ± 12.201 a | 434.101–613.096 |
| 50 | 6.70 ± 0.20 b | 4.56 ± 0.13 f | |||||
| 100 | 6.15 ± 0.13 c | 12.39 ± 0.68 e | |||||
| 200 | 5.17 ± 0.09 d | 26.35 ± 2.13 d | |||||
| 400 | 3.77 ± 0.24 e | 46.30 ± 1.89 c | |||||
| 800 | 2.82 ± 0.11 f | 59.83 ± 1.73 b | |||||
| 1600 | 1.50 ± 0.10 g | 78.63 ± 1.13 a | |||||
| Control | 7.02 ± 0.15 a | — | |||||
| Tebuconazole (97.5%) | 5 | 6.59 ± 0.12 b | 6.13 ± 0.12 g | y = 1.452x − 2.453 | 0.997 | 48.965 ± 1.231 b | 41.389–58.126 |
| 10 | 5.95 ± 0.14 c | 15.24 ± 0.73 f | |||||
| 20 | 4.81 ± 0.11 d | 31.48 ± 2.06 e | |||||
| 40 | 3.73 ± 0.11 e | 46.87 ± 0.93 d | |||||
| 80 | 2.83 ± 0.13 f | 59.69 ± 1.86 c | |||||
| 160 | 1.52 ± 0.10 g | 78.35 ± 2.56 b | |||||
| 320 | 0.90 ± 0.07 h | 87.18 ± 2.40 a | |||||
| Control | 7.02 ± 0.11 a | — | |||||
| Chlorothalonil (98%) | 1 | 6.44 ± 0.15 b | 8.26 ± 0.14 g | y = 1.025x − 1.004 | 0.997 | 11.453 ± 0.333 c | 9.414–123.975 |
| 2 | 5.72 ± 0.11 c | 18.52 ± 1.06 f | |||||
| 5 | 4.60 ± 0.10 d | 34.47 ± 1.30 e | |||||
| 10 | 3.60 ± 0.11 e | 48.72 ± 1.29 d | |||||
| 20 | 2.94 ± 0.13 f | 58.12 ± 1.39 c | |||||
| 50 | 1.41 ± 0.11 g | 79.91 ± 1.83 b | |||||
| 100 | 0.90 ± 0.12 h | 87.18 ± 1.99 a | |||||
| Control | 7.02 ± 0.11 a | — |
| Fungicide | Concentrations (μg/mL) | Colony Diameter (cm) | Inhibition Rates (%) | Toxicity Regression Equations | R Correlation Coefficient | EC50 (μg/mL) | 95% CL (ug/mL) |
|---|---|---|---|---|---|---|---|
| Prochloraz (95%) | 0.016 | 7.58 ± 0.13 b | 8.12 ± 0.27 g | y = 1.172x + 0.926 | 0.979 | 0.162 ± 0.009 d | 0.133–0.200 |
| 0.032 | 6.14 ± 0.16 c | 25.58 ± 1.84 f | |||||
| 0.064 | 5.25 ± 0.15 d | 36.36 ± 1.69 e | |||||
| 0.128 | 4.62 ± 0.18 e | 44.00 ± 1.32 d | |||||
| 0.256 | 3.92 ± 0.15 f | 52.48 ± 1.41 c | |||||
| 0.512 | 2.56 ± 0.17 f | 68.97 ± 1.38 b | |||||
| 1.024 | 1.05 ± 0.15 h | 87.27 ± 1.82 a | |||||
| Control | 8.25 ± 0.13 a | — | |||||
| Carbendazim (95%) | 1.875 | 7.26 ± 0.12 b | 12.00 ± 0.50 g | y = 1.154x − 1.509 | 0.993 | 20.304 ± 0.882 a | 16.590–25.122 |
| 3.75 | 6.54 ± 0.15 c | 20.73 ± 0.86 f | |||||
| 7.5 | 5.54 ± 0.17 d | 32.85 ± 1.77 e | |||||
| 15 | 4.72 ± 0.11 e | 42.79 ± 1.36 d | |||||
| 30 | 3.97 ± 0.11 f | 51.88 ± 2.01 c | |||||
| 60 | 2.44 ± 0.16 g | 70.42 ± 1.15 b | |||||
| 120 | 1.25 ± 0.10 h | 84.85 ± 1.53 a | |||||
| Control | 8.25 ± 0.13 a | — | |||||
| Azoxystrobin (98.5%) | 0.5 | 7.03 ± 0.14 a | 14.79 ± 1.44 g | y = 1.095x − 0.698 | 0.997 | 4.337 ± 0.179 c | 3.508–5.379 |
| 1 | 6.14 ± 0.22 b | 25.58 ± 0.81 f | |||||
| 2 | 5.29 ± 0.13 c | 35.88 ± 1.72 e | |||||
| 4 | 4.38 ± 0.16 d | 46.91 ± 1.50 d | |||||
| 8 | 3.12 ± 0.16 e | 62.18 ± 1.64 c | |||||
| 16 | 2.03 ± 0.14 f | 75.39 ± 1.19 b | |||||
| 32 | 1.55 ± 0.15 g | 81.21 ± 1.22 a | |||||
| Control | 8.25 ± 0.13 a | — | |||||
| Tebuconazole (97.5%) | 0.05 | 7.52 ± 0.17 b | 8.85 ± 0.25 g | y = 1.297x + 0.371 | 0.997 | 0.518 ± 0.009 d | 0.431–0.626 |
| 0.1 | 6.62 ± 0.20 c | 19.76 ± 1.01 f | |||||
| 0.2 | 5.84 ± 0.12 d | 29.21 ± 2.16 e | |||||
| 0.4 | 4.93 ± 0.16 e | 40.24 ± 2.12 d | |||||
| 0.8 | 3.08 ± 0.20 f | 62.67 ± 1.66 c | |||||
| 1.6 | 2.16 ± 0.17 g | 73.82 ± 2.37 d | |||||
| 3.2 | 1.28 ± 0.24 h | 84.48 ± 1.22 a | |||||
| Control | 8.25 ± 0.13 a | — | |||||
| Chlorothalonil (98%) | 1 | 7.66 ± 0.15 b | 7.15 ± 0.38 g | y = 1.198x − 1.409 | 0.994 | 15.021 ± 0.362 b | 12.213–18.549 |
| 2 | 6.82 ± 0.17 c | 17.33 ± 1.46 f | |||||
| 5 | 5.84 ± 0.14 d | 29.21 ± 1.31 e | |||||
| 10 | 4.93 ± 0.16 e | 40.24 ± 1.31 d | |||||
| 20 | 3.86 ± 0.21 f | 53.21 ± 1.35 c | |||||
| 50 | 2.45 ± 0.17 g | 70.30 ± 0.89 b | |||||
| 100 | 1.01 ± 0.25 h | 87.76 ± 1.99 a | |||||
| Control | 8.25 ± 0.13 a | — |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Song, M.; Li, Y.; Zhang, L.; Zhu, Z.; Li, L.; Wang, L. Study on the Identification, Biological Characteristics, and Fungicide Sensitivity of the Causal Agent of Strawberry Red Core Root Rot. Horticulturae 2024, 10, 771. https://doi.org/10.3390/horticulturae10070771
Zhang Y, Song M, Li Y, Zhang L, Zhu Z, Li L, Wang L. Study on the Identification, Biological Characteristics, and Fungicide Sensitivity of the Causal Agent of Strawberry Red Core Root Rot. Horticulturae. 2024; 10(7):771. https://doi.org/10.3390/horticulturae10070771
Chicago/Turabian StyleZhang, Yiming, Minyan Song, Yanan Li, Lina Zhang, Zhi Zhu, Liqi Li, and Li Wang. 2024. "Study on the Identification, Biological Characteristics, and Fungicide Sensitivity of the Causal Agent of Strawberry Red Core Root Rot" Horticulturae 10, no. 7: 771. https://doi.org/10.3390/horticulturae10070771
APA StyleZhang, Y., Song, M., Li, Y., Zhang, L., Zhu, Z., Li, L., & Wang, L. (2024). Study on the Identification, Biological Characteristics, and Fungicide Sensitivity of the Causal Agent of Strawberry Red Core Root Rot. Horticulturae, 10(7), 771. https://doi.org/10.3390/horticulturae10070771